
Engineering Resistance against Sclerotinia sclerotiorum Using a Truncated NLR (TNx) and a Defense-Priming Gene

 , , and
, , and 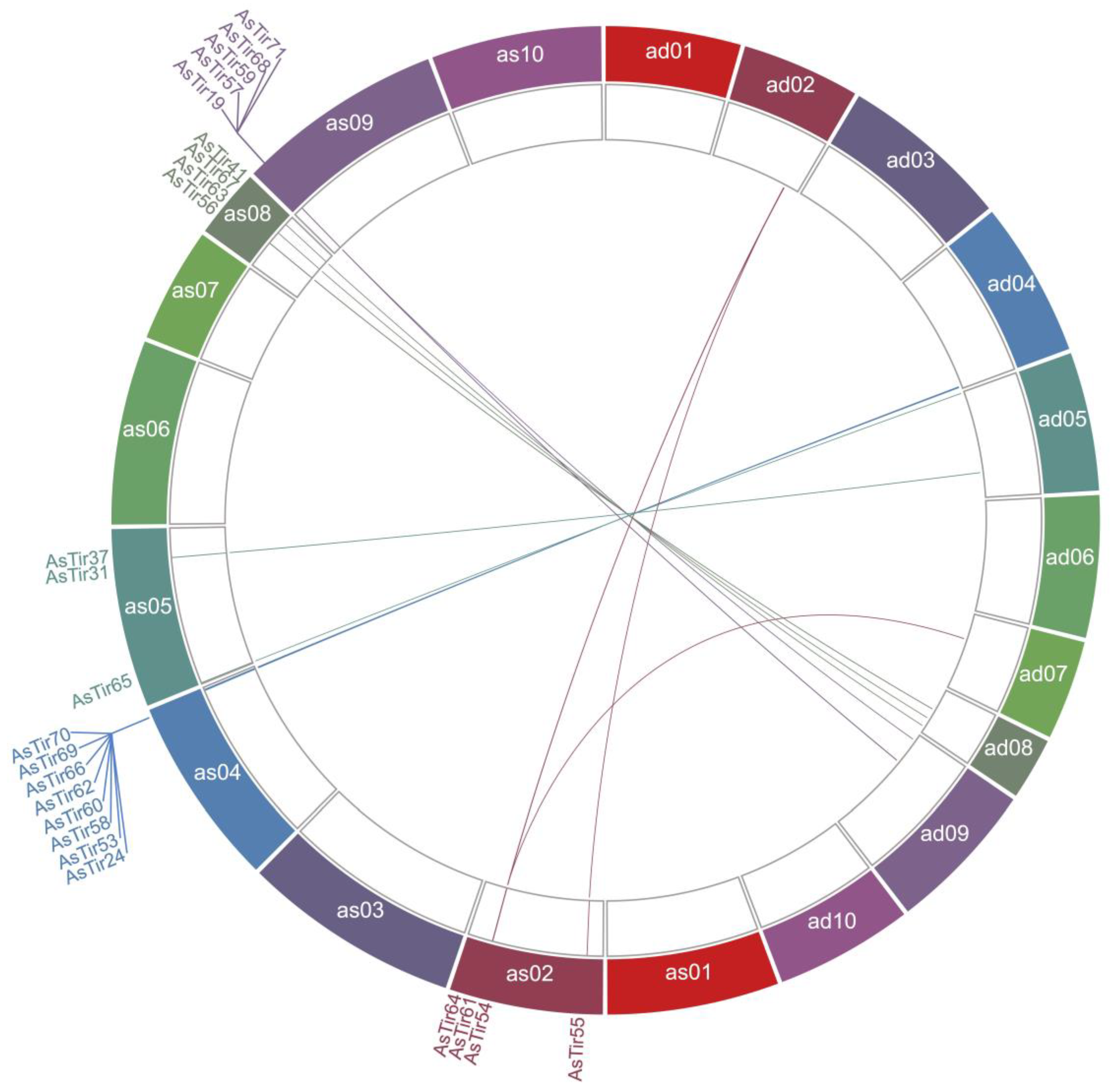{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Truncated NLR (TNx) Genomic Distribution, Gene Structure and Protein Domains
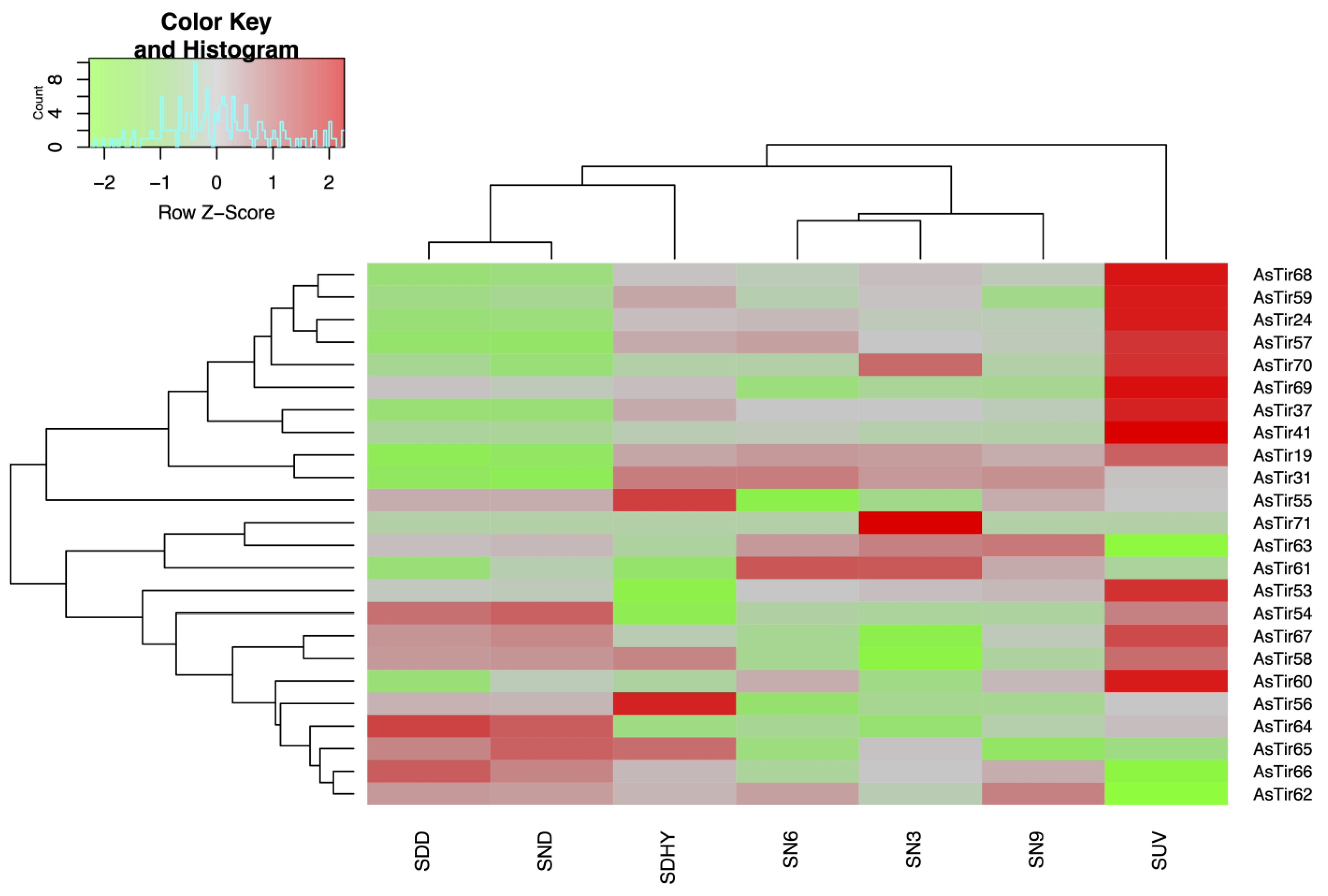
2.2. TNx Genes in A. stenosperma Are Responsive to Biotic and Abiotic Stresses
2.3. AsTIR19 Codes for an NLR-ID Fused Protein
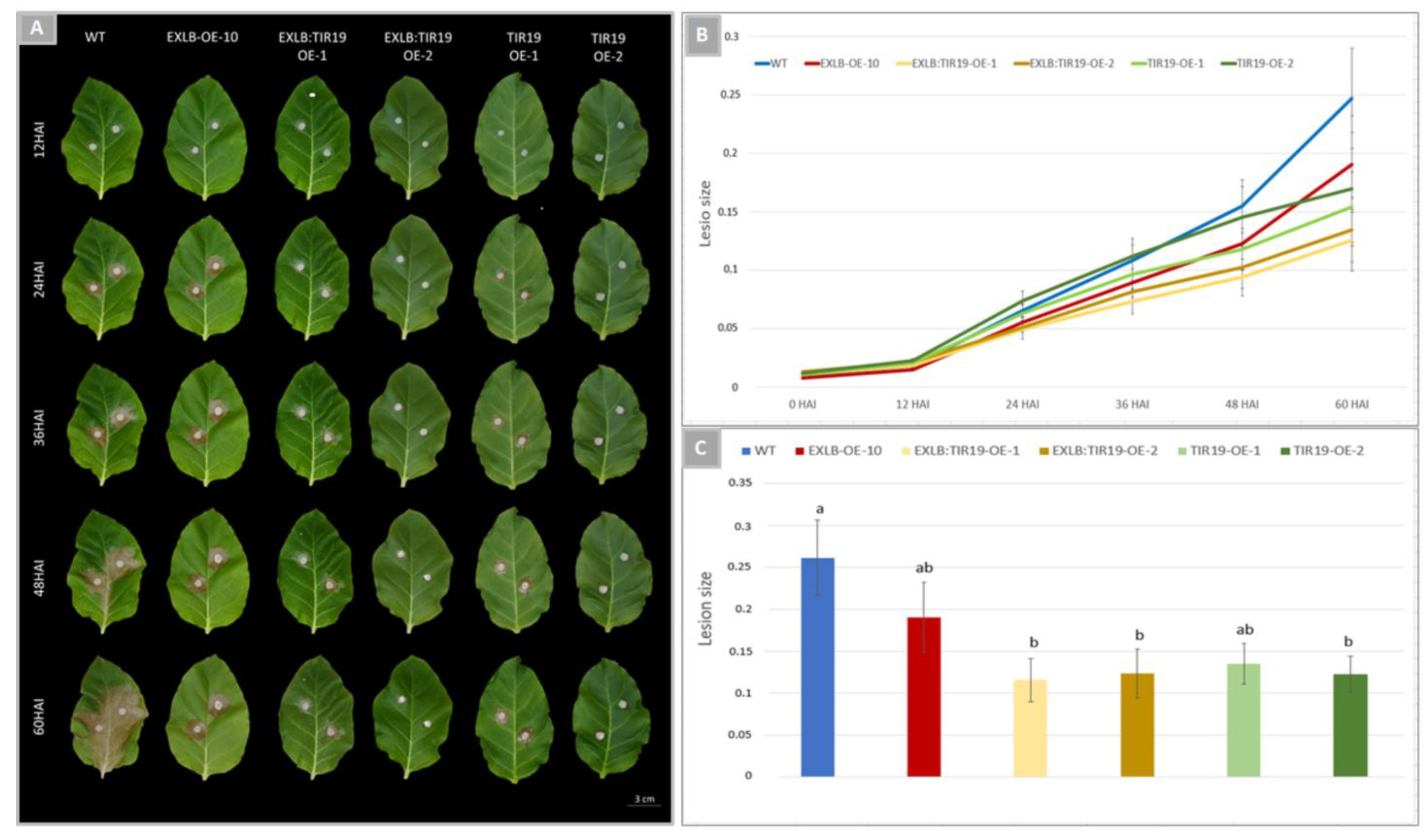
2.4. Overexpression of AsTIR19 and AdEXLB:AsTIR19 Reduces S. sclerotiorum Infection in Tobacco OE Lines
2.5. S. sclerotiorum Hyphae Growth in OE Lines
2.6. qRT-PCR Analysis of Marker Genes for Hormonal Defense Pathways in Tobacco OE Lines
3. Discussion
3.1. AsTIR19 Is a Broad-Spectrum Responsive TNx
3.2. AsTIR19 and AdEXLB8:AsTIR19 Induce Hormonal Defense Pathways in Tobacco OE Lines
3.3. RbohD Mediates ETI and PTI Defense Responses in Pyramid Lines
3.4. Novel Strategies for the Use of NLR Pyramids in Pathogen Control
4. Materials and Methods
4.1. Genomic Distribution of TNx Genes in A. stenosperma
4.2. Expression Profile of A. stenosperma TNx Genes in Response to Stress
4.3. qRT-PCR Expression Analysis of AsTIR19 in A. stenosperma Plants under Different Stresses
4.4. AsTIR19 Characterization and Cloning
4.5. AsTIR19 and AdEXLB: AsTIR19 Overexpression in Tobacco Plants
4.6. S. sclerotiorum Bioassays in TIR19 and EXLB:TIR19 OE Lines
4.7. Microscopy Analysis of S. sclerotiorum Infection in the OE Lines
4.8. Hormonal and Transcriptional Regulation in OE Lines
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACC | aminocyclopropane-1-carboxylate deaminase |
| ACS | 1-aminocyclopropane-1-carboxylate synthase |
| APX1 | Ascorbate peroxidase |
| CA | Beta-Carbonic anhydrase |
| CAT | catalase |
| CC domain | Coiled-Coil Domain |
| CPKs | Calcium-dependent Protein Knases |
| CDS | Coding Sequence |
| DAI | Days after Inoculation |
| DEG | Differentially Expressed Gene |
| EDS1 | Enhanced Disease Susceptibility1 |
| EF26 | Ethylene-forming enzyme |
| ET | Ethylene |
| ETI | Effector Triggered Immunity |
| FC | Fold Change |
| FDR | False Discovery Rate |
| GO | Gene Ontology |
| HAI | Hours after Inoculation |
| HR | Hypersensitive Response |
| HIN1 | Harpin-induced gene 1 |
| HSR515 | Hypersensitivity-related 515 |
| HSR201 | Hypersensitivity-related 201 |
| ID | Integrated Domains |
| J2 | Second-stage Juveniles |
| JA | Jasmonic Acid |
| LRR | Leucine Rich Repeat |
| MAMPS | Microbe-Associated Molecular Pattern |
| MAPK | Mitogen-Activated Protein Kinase |
| NACHT | NTPase domain found in apoptosis proteins as well as those involved in MHC transcription activation |
| NAD | nicotinamide adenine nucleotide |
| NBS | nucleotide-binding site |
| NLR | Nucleotide-binding and Leucine-rich repeat LRR domains |
| NPR1 | Nonexpressor of Pathogenesis-Related Genes 1 |
| OA | Oxalic Acid |
| OE | Overexpression Lines |
| PAMPs | Pathogen Associated Molecular Patterns |
| PB1 domain | Phox/Bem1p |
| PK | Protein Kinase |
| PR | Pathogenesis related |
| PRR | Pattern Recognition Receptors |
| PTI | Patterns Triggered Immunity |
| qRT-PCR | quantitative Reverse Transcription-Polymerase Chain Reaction |
| RbohD | NADPH/respiratory burst oxidase protein D |
| RKN | Root-Knot Nematode |
| RLKs | Receptor-Like Kinases |
| RLM3 | Leptosphaeria maculans 3 gene |
| RLPs | Receptor-Like Proteins |
| RPW8 | Resistance to Powdery Mildew 8 |
| RQ | Relative Quantification |
| ROS | Reactive Oxygen Species |
| SA | Salicylic Acid |
| SAR | Systemic Acquired Resistance |
| SDD | Submitted to dry-down |
| SND | Submitted to combined nematode and drought stress |
| SDHY | Submitted to dehydration |
| SN3, SN6, SN9 | Submitted to nematode infection at 3, 6 and 9 Days After Infection |
| SNPs | Single Nucleotide Polymorphism |
| SUV | Submitted to UV exposur |
| SsPINE1 | Sclerotinia sclerotiorum PGIP-Inactivating Effector 1 |
| TaRCR1 | Triticum aestivum RCR1 gene |
| TIR | Toll and IL-1 receptor |
| TF | Transcription Factor |
| TNL | TIR-NBS-LRR |
| TNx | TIR-NBS |
| UV | Ultraviolet |
| WT | Wild Type |
References
- Han, G. Origin and evolution of the plant immune system. N. Phytol. 2019, 222, 70–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karrer, E.E.; Beachy, R.N.; Holt, C.A. Cloning of tobacco genes that elicit the hypersensitive response. Plant Mol. Biol. 1998, 36, 681–690. [Google Scholar] [CrossRef] [PubMed]
- Saijo, Y.; Loo, E.P.; Yasuda, S. Pattern recognition receptors and signaling in plant-microbe interactions. Plant J. 2018, 93, 592–613. [Google Scholar] [CrossRef] [PubMed]
- Boutrot, F.; Zipfel, C. Function, Discovery, and Exploitation of Plant Pattern Recognition Receptors for Broad-Spectrum Disease Resistance. Annu. Rev. Phytopathol. 2017, 55, 257–286. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Kumar, V.; Olson, A.; Ware, D. Reviving the transcriptome studies: An Insight Into the Emergence of Single-Molecule Transcriptome Sequencing. Front. Genet. 2019, 10, 384. [Google Scholar] [CrossRef] [Green Version]
- Martin, R.; Qi, T.; Zhang, H.; Liu, F.; King, M.; Toth, C.; Nogales, E.; Staskawicz, B.J. Structure of the activated ROQ1 resistosome directly recognizing the pathogen effector XopQ. Science 2020, 370, eabd9993. [Google Scholar] [CrossRef]
- Saile, S.C.; Jacob, P.; Castel, B.; Jubic, L.M.; Salas-Gonzáles, I.; Bäcker, M.; Jones, J.D.G.; Dangl, J.L.; El Kasmi, F. Two unequally redundant “helper” immune receptor families mediate Arabidopsis thaliana intracellular “sensor” immune receptor functions. PLoS Biol. 2020, 18, e3000783. [Google Scholar] [CrossRef]
- Wu, Z.; Li, M.; Dong, O.X.; Xia, S.; Liang, W.; Bao, Y.; Wasteneys, G.; Li, X. Differential regulation of TNL-mediated immune signaling by redundant helper CNLs. N. Phytol. 2019, 222, 938–953. [Google Scholar] [CrossRef]
- Yuan, M.; Ngou, B.P.M.; Ding, P.; Xin, X.-F. PTI-ETI crosstalk: An integrative view of plant immunity. Curr. Opin. Plant Biol. 2021, 62, 102030. [Google Scholar] [CrossRef]
- Ngou, B.P.M.; Ahn, H.-K.; Ding, P.; Jones, J.D.G. Mutual potentiation of plant immunity by cell-surface and intracellular receptors. Nature 2021, 592, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.-M.; Iswanto, A.B.B.; Son, G.H.; Kim, S.H. Recent Advances in Effector-Triggered Immunity in Plants: New Pieces in the Puzzle Create a Different Paradigm. Int. J. Mol. Sci. 2021, 22, 4709. [Google Scholar] [CrossRef] [PubMed]
- Pruitt, R.N.; Gust, A.A.; Nürnberger, T. Plant immunity unified. Nat. Plants 2021, 7, 382–383. [Google Scholar] [CrossRef] [PubMed]
- Takken, F.L.; Albrecht, M.; Tameling, W. IL Resistance proteins: Molecular switches of plant defence. Curr. Opin. Plant Biol. 2006, 9, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Takken, F.L.; Goverse, A. How to build a pathogen detector: Structural basis of NB-LRR function. Curr. Opin. Plant Biol. 2012, 15, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Baggs, E.; Dagdas, G.; Krasileva, K. NLR diversity, helpers and integrated domains: Making sense of the NLR IDentity. Curr. Opin. Plant Biol. 2017, 38, 59–67. [Google Scholar] [CrossRef] [PubMed]
- de Araújo, A.C.; Fonseca, F.C.D.A.; Cotta, M.G.; Alves, G.S.C.; Miller, R.N.G. Plant NLR receptor proteins and their potential in the development of durable genetic resistance to biotic stresses. Biotechnol. Res. Innov. 2019, 3, 80–94. [Google Scholar] [CrossRef]
- Grund, E.; Tremousaygue, D.; Deslandes, L. Plant NLRs with Integrated Domains: Unity Makes Strength. Plant Physiol. 2019, 179, 1227–1235. [Google Scholar] [CrossRef] [Green Version]
- Borrelli, G.M.; Mazzucotelli, E.; Marone, D.; Crosatti, C.; Michelotti, V.; Valè, G.; Mastrangelo, A.M. Regulation and Evolution of NLR Genes: A Close Interconnection for Plant Immunity. Int. J. Mol. Sci. 2018, 19, 1662. [Google Scholar] [CrossRef] [Green Version]
- Shao, Z.-Q.; Xue, J.-Y.; Wang, Q.; Wang, B.; Chen, J.-Q. Revisiting the Origin of Plant NBS-LRR Genes. Trends Plant Sci. 2019, 24, 9–12. [Google Scholar] [CrossRef]
- Lorang, J.M.; Sweat, T.A.; Wolpert, T.J. Plant disease susceptibility conferred by a “resistance” gene. Proc. Natl. Acad. Sci. USA 2007, 104, 14861–14866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbacci, A.; Navaud, O.; Mbengue, M.; Barascud, M.; Godiard, L.; Khafif, M.; Lacaze, A.; Raffaele, S. Rapid identification of an Arabidopsis NLR gene as a candidate conferring susceptibility to Sclerotinia sclerotiorum using time-resolved automated phenotyping. Plant J. 2020, 103, 903–917. [Google Scholar] [CrossRef] [PubMed]
- Staal, J.; Kaliff, M.; Dewaele, E.; Persson, M.; Dixelius, C. RLM3, a TIR domain encoding gene involved in broad-range immunity of Arabidopsis to necrotrophic fungal pathogens. Plant J. 2008, 55, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Lu, C.; Du, L.; Ye, X.; Liu, X.; Coules, A.; Zhang, Z. The wheat NB-LRR gene TaRCR1 is required for host defence response to the necrotrophic fungal pathogen Rhizoctonia cerealis. Plant Biotechnol. J. 2017, 15, 674–687. [Google Scholar] [CrossRef] [Green Version]
- Sagi, M.S.; Deokar, A.A.; Tar’an, B. Genetic Analysis of NBS-LRR Gene Family in Chickpea and Their Expression Profiles in Response to Ascochyta Blight Infection. Front. Plant Sci. 2017, 8, 838. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Rollins, J.A. Mechanisms of Broad Host Range Necrotrophic Pathogenesis in Sclerotinia sclerotiorum. Phytopathology 2018, 108, 1128–1140. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Zhu, Y.-X.; Balint-Kurti, P.J.; Wang, G.-F. Fine-Tuning Immunity: Players and Regulators for Plant NLRs. Trends Plant Sci. 2020, 25, 695–713. [Google Scholar] [CrossRef]
- Wang, Z.; Wan, L.; Xin, Q.; Chen, Y.; Zhang, X.; Dong, F.; Hong, D.; Yang, G. Overexpression of OsPGIP2 confers Sclerotinia sclerotiorum resistance in Brassica napus through increased activation of defense mechanisms. J. Exp. Bot. 2018, 69, 3141–3155. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, W.-H.; Ma, L.-Y.; Li, X.; Zhao, F.-Y.; Tan, X.-L. Overexpression of Brassica napus NPR1 enhances resistance to Sclerotinia sclerotiorum in oilseed rape. Physiol. Mol. Plant Pathol. 2020, 110, 101460. [Google Scholar] [CrossRef]
- Verma, R.; Kaur, J. Expression of barley oxalate oxidase confers resistance against Sclerotinia sclerotiorum in transgenic Brassica juncea cv Varuna. Transgenic Res. 2021, 30, 143–154. [Google Scholar] [CrossRef]
- Maximiano, M.R.; Santos, L.S.; Santos, C.; Aragão, F.J.L.; Dias, S.C.; Franco, O.L.; Mehta, A. Host induced gene silencing of Sclerotinia sclerotiorum effector genes for the control of white mold. Biocatal. Agric. Biotechnol. 2022, 40, 102302. [Google Scholar] [CrossRef]
- Tamborski, J.; Krasileva, K.V. Evolution of Plant NLRs: From Natural History to Precise Modifications. Annu. Rev. Plant Biol. 2020, 71, 355–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, S.; Kim, S.; Lee, K.; Oh, J.; Choi, I.; Do, J.; Yoon, J.; Han, J.; Park, S. The Capsicum baccatum-Specific Truncated NLR Protein CbCN Enhances the Innate Immunity against Colletotrichum acutatum. Int. J. Mol. Sci. 2021, 22, 7672. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Rui, L.; Li, J.; Nishimura, M.T.; Vogel, J.P.; Liu, N.; Liu, S.; Zhao, Y.; Dangl, J.L.; Tang, D. A truncated NLR protein, TIR-NBS2, is required for activated defense responses in the exo70B1 mutant. PLoS Genet. 2015, 11, e1004945. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Wersch, S.; Tong, M.; Li, X. TIR-NB-LRR immune receptor SOC3 pairs with truncated TIR-NB protein CHS1 or TN2 to monitor the homeostasis of E3 ligase SAUL1. N. Phytol. 2019, 221, 2054–2066. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, F.; Nishimura, M.T. Structural, functional, and genomic diversity of plant NLR proteins: An evolved resource for rational engineering of plant immunity. Annu. Rev. Phytopathol. 2018, 56, 243–267. [Google Scholar] [CrossRef] [Green Version]
- Roth, C.; Lüdke, D.; Klenke, M.; Quathamer, A.; Valerius, O.; Braus, G.H.; Wiermer, M. The truncated NLR protein TIR-NBS13 is a MOS6/IMPORTIN-α3 interaction partner required for plant immunity. Plant J. 2017, 92, 808–821. [Google Scholar] [CrossRef]
- Lai, Y.; Eulgem, T. Transcript-level expression control of plant NLR genes. Mol. Plant Pathol. 2018, 19, 1267–1281. [Google Scholar] [CrossRef] [Green Version]
- Kabbage, M.; Yarden, O.; Dickman, M.B. Pathogenic attributes of Sclerotinia sclerotiorum: Switching from a biotrophic to necrotrophic lifestyle. Plant Sci. 2015, 233, 53–60. [Google Scholar] [CrossRef]
- McCaghey, M.; Willbur, J.; Smith, D.L.; Kabbage, M. The complexity of the Sclerotinia sclerotiorum pathosystem in soybean: Virulence factors, resistance mechanisms, and their exploitation to control Sclerotinia stem rot. Trop. Plant Pathol. 2019, 44, 12–22. [Google Scholar] [CrossRef]
- Wei, W.; Xu, L.; Peng, H.; Zhu, W.; Tanaka, K.; Cheng, J.; Sanguinet, K.A.; Vandemark, G.; Chen, W. A fungal extracellular effector inactivates plant polygalacturonase-inhibiting protein. Nat. Commun. 2022, 13, 2213. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.-P.; Lv, L.-H.; Xu, Y.-J.; Yang, J.; Cao, J.-Y.; Cai, X.-Z. Leaf stage-associated resistance is correlated with phytohormones in a pathosystem-dependent manner. J. Integr. Plant Biol. 2018, 60, 703–722. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ma, L.-Y.; Cao, J.; Li, Y.-L.; Ding, L.-N.; Zhu, K.-M.; Yang, Y.-H.; Tan, X.-L. Recent Advances in Mechanisms of Plant Defense to Sclerotinia sclerotiorum. Front. Plant Sci. 2019, 10, 1314. [Google Scholar] [CrossRef] [PubMed]
- Warburton, M.L.; Rauf, S.; Marek, L.; Hussain, M.; Ogunola, O.; de Jesus Sanchez Gonzalez, J. The Use of Crop Wild Relatives in Maize and Sunflower Breeding. Crop. Sci. 2017, 57, 1227–1240. [Google Scholar] [CrossRef]
- Funk, A.; Galewski, P.; McGrath, J.M. Nucleotide-binding resistance gene signatures in sugar beet, insights from a new reference genome. Plant J. 2018, 95, 659–671. [Google Scholar] [CrossRef] [Green Version]
- Guimarães, P.M.; Brasileiro, A.C.M.; Morgante, C.V.; Martins, A.C.Q.; Pappas, G.; Silva, O.B.; Togawa, R.; Leal-Bertioli, S.C.M.; Araújo, A.C.G.; Moretzsohn, M.C.; et al. Global transcriptome analysis of two wild relatives of peanut under drought and fungi infection. BMC Genom. 2012, 13, 387. [Google Scholar] [CrossRef] [Green Version]
- Guimarães, P.M.; Guimarães, L.A.; Morgante, C.V.; Silva, O.B.; Araújo, A.C.G.; Martins, A.C.Q.; Saraiva, M.A.P.; Oliveira, T.N.; Togawa, R.C.; Leal-Bertioli, S.C.M.; et al. Root transcriptome analysis of wild peanut reveals candidate genes for nematode resistance. PLoS ONE 2015, 10, e0140937. [Google Scholar] [CrossRef]
- Guimarães, P.M.; Brasileiro, A.C.M.; Mehta, A.; Araújo, A.C.G. Functional genomics in peanut wild relatives. In The Peanut Genome-Compendium of Plant Genomes; Varshney, R.K., Pandey, M.K., Puppala, N., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 149–164. [Google Scholar]
- Mota, A.P.Z.; Vidigal, B.; Danchin, E.G.J.; Togawa, R.C.; Leal-Bertioli, S.C.M.; Bertioli, D.J.; Araújo, A.C.G.; Brasileiro, A.C.M.; Guimarães, P.M. Comparative root transcriptome of wild Arachis reveals NBS-LRR genes related to nematode resistance. BMC Plant Biol. 2018, 18, 159. [Google Scholar] [CrossRef]
- Martins, A.C.Q.; Mota, A.P.Z.; Carvalho, P.A.S.V.; Passos, M.A.S.; Gimenes, M.A.; Guimaraes, P.M.; Brasileiro, A.C.M. Transcriptome Responses of Wild Arachis to UV-C Exposure Reveal Genes Involved in General Plant Defense and Priming. Plants 2022, 11, 408. [Google Scholar] [CrossRef]
- Vinson, C.C.; Mota, A.P.Z.; Oliveira, T.N.; Guimarães, L.A.; Leal-Bertioli, S.C.M.; Williams, T.C.R.; Nepomuceno, A.L.; Saraiva, M.A.P.; Araújo, A.C.G.; Guimarães, P.M.; et al. Early responses to dehydration in contrasting wild Arachis species. PLoS ONE 2018, 13, e0198191. [Google Scholar] [CrossRef]
- Brasileiro, A.C.M.; Lacorte, C.; Pereira, B.M.; Oliveira, T.N.; Ferreira, D.S.; Mota, A.P.Z.; Saraiva, M.A.P.; Araújo, A.C.G.; Paulino Silva, L.; Guimarães, P.M. Ectopic expression of an expansin-like B gene from wild Arachis enhances tolerance to both abiotic and biotic stresses. Plant J. 2021, 10, 1681–1696. [Google Scholar] [CrossRef] [PubMed]
- Bertioli, D.J.; Seijo, G.; Freitas, F.O.; Valls, J.F.M.; Leal-Bertioli, S.C.M.; Moretzsohn, M.C. An overview of peanut and its wild relatives. Plant Genet. Resour. 2011, 9, 134–149. [Google Scholar] [CrossRef] [Green Version]
- Guimarães, L.A.; Mota, A.P.Z.; Araújo, A.C.G.; Alencar Figueiredo, L.F.; Pereira, B.M.; Passos Saraiva, M.A.; Silva, R.B.; Danchin, E.G.J.J.; Guimarães, P.M.; Brasileiro, A.C.M. Genome-wide analysis of expansin superfamily in wild Arachis discloses a stress-responsive expansin-like B gene. Plant Mol. Biol. 2017, 94, 79–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumimoto, H.; Kamakura, S.; Ito, T. Structure and Function of the PB1 Domain, a Protein Interaction Module Conserved in Animals, Fungi, Amoebas, and Plants. Sci. STKE 2007, 2007, re6. [Google Scholar] [CrossRef] [PubMed]
- Kufer, T.A.; Fritz, J.H.; Philpott, D.J. NACHT-LRR proteins (NLRs) in bacterial infection and immunity. Trends Microbiol. 2005, 13, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Kourelis, J.; van der Hoorn, R.A.L. Defended to the Nines: 25 years of Resistance Gene Cloning Identifies Nine Mechanisms for R Protein Function. Plant Cell 2018, 30, 285–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhang, Y.; Yin, L.; Lu, J. Overexpression of pathogen-induced grapevine TIR-NB-LRR gene VaRGA1 enhances disease resistance and drought and salt tolerance in Nicotiana benthamiana. Protoplasma 2017, 254, 957–969. [Google Scholar] [CrossRef]
- Ren, S.; Sun, M.; Yan, H.; Wu, B.; Jing, T.; Huang, L.; Zeng, B. Identification and Distribution of NBS-Encoding Resistance Genes of Dactylis glomerata L. and Its Expression Under Abiotic and Biotic Stress. Biochem. Genet. 2020, 58, 824–847. [Google Scholar] [CrossRef]
- Thorvaldsdottir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef] [Green Version]
- van Ooijen, G.; Mayr, G.; Kasiem, M.M.A.; Albrecht, M.; Cornelissen, B.J.C.; Takken, F.L.W. Structure–function analysis of the NB-ARC domain of plant disease resistance proteins. J. Exp. Bot. 2008, 59, 1383–1397. [Google Scholar] [CrossRef]
- Qi, D.; Innes, R.W. Recent Advances in Plant NLR Structure, Function, Localization, and Signaling. Front. Immunol. 2013, 4, 348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyers, B.C.; Morgante, M.; Michelmore, R.W. TIR-X and TIR-NBS proteins: Two new families related to disease resistance TIR-NBS-LRR proteins encoded in Arabidopsis and other plant genomes. Plant J. 2002, 32, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Sarris, P.F.; Cevik, V.; Dagdas, G.; Jones, J.D.G.; Krasileva, K.V. Comparative analysis of plant immune receptor architectures uncovers host proteins likely targeted by pathogens. BMC Biol. 2016, 14, 8. [Google Scholar] [CrossRef] [Green Version]
- Mundt, C.C. Pyramiding for resistance durability: Theory and practice. Phytopathology 2018, 108, 792–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, C.; Guo, Z.-H.; Hao, P.-P.; Wang, G.-M.; Jin, Z.-M.; Zhang, S.-L. Multiple regulatory roles of AP2/ERF transcription factor in angiosperm. Bot. Stud. 2017, 58, 6. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Alférez, F.M.; Johnson, E.G.; Graham, J.H. Up-regulation of PR1 and less disruption of hormone and sucrose metabolism in roots is associated with lower susceptibility to ‘Candidatus Liberibacter asiaticus’. Plant Pathol. 2018, 67, 1426–1435. [Google Scholar] [CrossRef]
- Pandey, V.P.; Awasthi, M.; Singh, S.; Tiwari, S.; Dwivedi, U.N. A Comprehensive Review on Function and Application of Plant Peroxidases. Biochem. Anal. Biochem. 2017, 06, 308. [Google Scholar] [CrossRef]
- Fichman, Y.; Mittler, R. Rapid systemic signaling during abiotic and biotic stresses: Is the ROS wave master of all trades? Plant J. 2020, 102, 887–896. [Google Scholar] [CrossRef] [Green Version]
- Si, Z.; Qiao, Y.; Zhang, K.; Ji, Z.; Han, J. Characterization of Nucleotide Binding Site-Encoding Genes in Sweetpotato, Ipomoea batatas (L.) Lam., and Their Response to Biotic and Abiotic Stresses. Cytogenet. Genome Res. 2021, 161, 257–271. [Google Scholar] [CrossRef]
- Richard, M.M.S.; Gratias, A.; Meyers, B.C.; Geffroy, V. Molecular mechanisms that limit the costs of NLR-mediated resistance in plants. Mol. Plant Pathol. 2018, 19, 2516–2523. [Google Scholar] [CrossRef]
- Wu, C.-H.; Derevnina, L.; Kamoun, S. Receptor networks underpin plant immunity. Science 2018, 360, 1300–1301. [Google Scholar] [CrossRef] [PubMed]
- Ariga, H.; Katori, T.; Tsuchimatsu, T.; Hirase, T.; Tajima, Y.; Parker, J.E.; Alcázar, R.; Koornneef, M.; Hoekenga, O.; Lipka, A.E.; et al. NLR locus-mediated trade-off between abiotic and biotic stress adaptation in Arabidopsis. Nat. Plants 2017, 3, 17072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dangl, J.L.; McDowell, J.M. Two modes of pathogen recognition by plants. Proc. Natl. Acad. Sci. USA 2006, 103, 8575–8576. [Google Scholar] [CrossRef] [Green Version]
- Bonardi, V.; Tang, S.; Stallmann, A.; Roberts, M.; Cherkis, K.; Dangl, J.L. Expanded functions for a family of plant intracellular immune receptors beyond specific recognition of pathogen effectors. Proc. Natl. Acad. Sci. USA 2011, 108, 16463–16468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesari, S.; Bernoux, M.; Moncuquet, P.; Kroj, T.; Dodds, P.N. A novel conserved mechanism for plant NLR protein pairs: The “integrated decoy” hypothesis. Front. Plant Sci. 2014, 5, 606. [Google Scholar] [CrossRef] [Green Version]
- Staal, J.; Dixelius, C. RLM3, a potential adaptor between specific TIR-NB-LRR receptors and DZC proteins. Commun. Integr. Biol. 2008, 1, 59–61. [Google Scholar] [CrossRef]
- Lin, Y.; Guo, R.; Ji, C.; Zhou, J.; Jiang, L. New insights into AtNBR1 as a selective autophagy cargo receptor in Arabidopsis. Plant Signal. Behav. 2021, 16, 1839226. [Google Scholar] [CrossRef]
- Zientara-Rytter, K.; Sirko, A. Significant role of PB1 and UBA domains in multimerization of Joka2, a selective autophagy cargo receptor from tobacco. Front. Plant Sci. 2014, 5, 13. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, Z. Broad and Complex Roles of NBR1-Mediated Selective Autophagy in Plant Stress Responses. Cells 2020, 9, 2562. [Google Scholar] [CrossRef]
- Cesari, S.; Xi, Y.; Declerck, N.; Chalvon, V.; Mammri, L.; Pugnière, M.; Henriquet, C.; de Guillen, K.; Chochois, V.; Padilla, A.; et al. New recognition specificity in a plant immune receptor by molecular engineering of its integrated domain. Nat. Commun. 2022, 13, 1524. [Google Scholar] [CrossRef]
- Larkan, N.J.; Lydiate, D.J.; Yu, F.; Rimmer, S.R.; Borhan, M.H. Co-localisation of the blackleg resistance genes Rlm2 and LepR3 on Brassica napus chromosome A10. BMC Plant Biol. 2014, 14, 387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkan, N.J.; Lydiate, D.J.; Parkin, I.A.P.; Nelson, M.N.; Epp, D.J.; Cowling, W.A.; Rimmer, S.R.; Borhan, M.H. The B rassica napus blackleg resistance gene LepR3 encodes a receptor-like protein triggered by the L eptosphaeria maculans effector AVRLM 1. New Phytol. 2013, 197, 595–605. [Google Scholar] [CrossRef]
- Dangl, J.L.; Horvath, D.M.; Staskawicz, B.J. Pivoting the Plant Immune System from Dissection to Deployment. Science 2013, 341, 746–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glazebrook, J. Contrasting Mechanisms of Defense Against Biotrophic and Necrotrophic Pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Pastrana, R.; Arcos-Ortega, G.F.; Souza-Perera, R.A.; Sánchez-Borges, C.A.; Nakazawa-Ueji, Y.E.; García-Villalobos, F.J.; Guzmán-Antonio, A.A.; Zúñiga-Aguilar, J.J. Ethylene, but not salicylic acid or methyl jasmonate, induces a resistance response against Phytophthora capsici in Habanero pepper. Eur. J. Plant Pathol. 2011, 131, 669–683. [Google Scholar] [CrossRef]
- Grover, A. Plant Chitinases: Genetic Diversity and Physiological Roles. Crit. Rev. Plant Sci. 2012, 31, 57–73. [Google Scholar] [CrossRef]
- Ganesan, M.; Bhanumathi, P.; Kumari, K.G.; Prabha, A.L.; Song, P.-S.; Jayabalan, N. Transgenic Indian Cotton (Gossypium hirsutum) Harboring Rice Chitinase Gene (Chi II) Confers Resistance to Two Fungal Pathogens. Am. J. Biochem. Biotechnol. 2009, 5, 63–74. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Han, X.; Feng, D.; Yuan, D.; Huang, L.-J. Signaling Crosstalk between Salicylic Acid and Ethylene/Jasmonate in Plant Defense: Do We Understand What They Are Whispering? Int. J. Mol. Sci. 2019, 20, 671. [Google Scholar] [CrossRef] [Green Version]
- Berrocal-Lobo, M.; Molina, A.; Solano, R. Constitutive expression of Ethylene-Response-Factor1 in arabidopsis confers resistance to several necrotrophic fungi. Plant J. 2002, 29, 23–32. [Google Scholar] [CrossRef]
- Helliwell, E.E.; Wang, Q.; Yang, Y. Transgenic rice with inducible ethylene production exhibits broad-spectrum disease resistance to the fungal pathogens Magnaporthe oryzae and Rhizoctonia solani. Plant Biotechnol. J. 2013, 11, 33–42. [Google Scholar] [CrossRef]
- Gilroy, S.; Suzuki, N.; Miller, G.; Choi, W.-G.; Toyota, M.; Devireddy, A.R.; Mittler, R. A tidal wave of signals: Calcium and ROS at the forefront of rapid systemic signaling. Trends Plant Sci. 2014, 19, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Li, N.-Y.; Zhou, L.; Zhang, D.-D.; Klosterman, S.J.; Li, T.-G.; Gui, Y.-J.; Kong, Z.-Q.; Ma, X.-F.; Short, D.P.G.; Zhang, W.-Q.; et al. Heterologous Expression of the Cotton NBS-LRR Gene GbaNA1 Enhances Verticillium Wilt Resistance in Arabidopsis. Front. Plant Sci. 2018, 9, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Dong, S.; Gu, F.; Liu, W.; Yang, G.; Huang, M.; Xiao, W.; Liu, Y.; Guo, T.; Wang, H.; et al. NBS-LRR Protein Pik-H4 Interacts with OsBIHD1 to Balance Rice Blast Resistance and Growth by Coordinating Ethylene-Brassinosteroid Pathway. Front. Plant Sci. 2017, 8, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goggin, F.L.; Jia, L.; Shah, G.; Hebert, S.; Williamson, V.M.; Ullman, D.E. Heterologous Expression of the Mi-1.2 Gene from Tomato Confers Resistance Against Nematodes but Not Aphids in Eggplant. Mol. Plant-Microbe Interact. 2006, 19, 383–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Li, Y.; Vossen, J.H.; Visser, R.G.F.; Jacobsen, E. Functional stacking of three resistance genes against Phytophthora infestans in potato. Transgenic Res. 2012, 21, 89–99. [Google Scholar] [CrossRef] [Green Version]
- Bertioli, D.J.; Cannon, S.B.; Froenicke, L.; Huang, G.; Farmer, A.D.; Cannon, E.K.S.; Liu, X.; Gao, D.; Clevenger, J.; Dash, S.; et al. The genome sequences of Arachis duranensis and Arachis ipaensis, the diploid ancestors of cultivated peanut. Nat. Genet 2016, 48, 438–446. [Google Scholar] [CrossRef]
- Vinson, C.C.; Mota, A.P.Z.; Porto, B.N.; Oliveira, T.N.; Sampaio, I.; Lacerda, A.L.; Danchin, E.G.J.; Guimarães, P.M.; Williams, T.C.R.; Brasileiro, A.C.M. Characterization of raffinose metabolism genes uncovers a wild Arachis galactinol synthase conferring tolerance to abiotic stresses. Sci. Rep. 2020, 10, 15258. [Google Scholar] [CrossRef]
- Morgante, C.V.; Brasileiro, A.C.M.; Roberts, P.A.; Guimaraes, L.A.; Araujo, A.C.G.; Fonseca, L.N.; Leal-Bertioli, S.C.M.; Bertioli, D.J.; Guimaraes, P.M. A survey of genes involved in Arachis stenosperma resistance to Meloidogyne arenaria race 1. Funct. Plant Biol. 2013, 40, 1298–1309. [Google Scholar] [CrossRef] [Green Version]
- Morgante, C.V.; Guimarães, P.M.; Martins, A.; Araújo, A.C.G.; Leal-Bertioli, S.C.M.; Bertioli, D.J.; Brasileiro, A.C.M. Reference genes for quantitative reverse transcription-polymerase chain reaction expression studies in wild and cultivated peanut. BMC Res. Notes 2011, 4, 339. [Google Scholar] [CrossRef] [Green Version]
- Mota, A.P.Z.; Brasileiro, A.C.M.; Vidigal, B.; Oliveira, T.N.; da Cunha Quintana Martins, A.; Saraiva, M.A.d.P.; de Araújo, A.C.G.; Togawa, R.C.; Grossi-de-Sá, M.F.; Guimaraes, P.M. Defining the combined stress response in wild Arachis. Sci. Rep. 2021, 11, 11097. [Google Scholar] [CrossRef]
- Horsch, R.B.; Fry, J.E.; Hoffmann, N.L.; Eichholtz, D.; Rogers, S.G.; Fraley, R.T. A simple and general method for transferring genes into plants. Science 1985, 227, 1229–1231. [Google Scholar] [CrossRef] [PubMed]
- Perchepied, L.; Balagué, C.; Riou, C.; Claudel-Renard, C.; Rivière, N.; Grezes-Besset, B.; Roby, D. Nitric oxide participates in the complex interplay of defense-related signaling pathways controlling disease resistance to Sclerotinia sclerotiorum in Arabidopsis thaliana. Mol. Plant-Microbe Interact. 2010, 23, 846–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Fernald, R.D. Comprehensive algorithm for quantitative real-time polymerase chain reaction. J. Comput. Biol. 2005, 12, 1047–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rancurel, C.; van Tran, T.; Elie, C.; Hilliou, F. SATQPCR: Website for statistical analysis of real-time quantitative PCR data. Mol. Cell. Probes 2019, 46, 101418. [Google Scholar] [CrossRef] [PubMed]
- Brasileiro, A.C.M.; Morgante, C.V.; Araujo, A.C.G.; Leal-Bertioli, S.C.M.; Silva, A.K.; Martins, A.C.Q.; Vinson, C.C.; Santos, C.M.R., C.M.R.; Bonfim, O.; Togawa, R.C.; et al. Proteomics unravels new candidate genes for Meloidogyne resistance in wild Arachis. J. Proteomics 2020, 217, 103690. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.C.Q.; Mehta, A.; Murad, A.M.; Mota, A.P.Z.; Saraiva, M.A.P.; Araújo, A.C.G.; Miller, R.N.G.; Brasileiro, A.C.M.; Guimarães, P.M. Proteomics unravels new candidate genes for Meloidogyne resistance in wild Arachis. J. Proteomics 2020, 217, 103690. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.B.; Monteiro, T.R.; Cabral, G.B.; Aragão, F.J.L. RNAi-mediated resistance to whitefly (Bemisia tabaci) in genetically engineered lettuce (Lactuca sativa). Transgenic Res. 2017, 26, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Han, Y.; Zhang, M.; Zhou, S.; Kong, X.; Wang, W. Overexpression of the wheat expansin gene TaEXPA2 improved seed production and drought tolerance in transgenic tobacco plants. PLoS One 2016, 11, e0153494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Chen, H.; Zhuang, R.-R.; Chen, Y.-T.; Deng, Y.; Cai, T.-C.; Wang, S.-Y.; Liu, Q.-Z.; Tang, R.-H.; Shan, S.-H. Overexpression of the peanut CLAVATA1-like leucine-rich repeat receptor-like kinase AhRLK1 confers increased resistance to bacterial wilt in tobacco. J. Exp. Bot. 2019, 70, 5407–5421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, L.; Cai, G.; Jiang, S.; Sun, L.; Li, D. Response of tobacco to the Pseudomonas syringae pv. tomato DC3000 is mainly dependent on salicylic acid signaling pathway. FEMS Microbiol. Lett. 2013, 344, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Liu, Q.; Liu, Y.; Zhang, L.; Ding, W. Overexpression of NtPR-Q up-regulates multiple defense-related genes in Nicotiana tabacum and enhances plant resistance to Ralstonia solanacearum. Front. Plant Sci. 2017, 8, 1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Zhang, X.; Jia, A.; Xu, G.; Hu, H.; Hu, X.; Hu, L. Jasmonate mediates salt-induced nicotine biosynthesis in tobacco (Nicotiana tabacum L.). Plant Divers. 2016, 38, 118–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alon, M.; Malka, O.; Eakteiman, G.; Elbaz, M.; Zvi, M.M.B.; Vainstein, A.; Morin, S. Activation of the phenylpropanoid pathway in Nicotiana tabacum improves the performance of the whitefly Bemisia tabaci via reduced jasmonate signaling. PLoS One 2013, 8, e76619. [Google Scholar] [CrossRef] [PubMed]
- Dang, F.; Lin, J.; Xue, B.; Chen, Y.; Guan, D.; Wang, Y.; He, S. CaWRKY27 negatively regulates H2O2-mediated thermotolerance in pepper (Capsicum annuum). Front. Plant Sci. 2018, 9, 1633. [Google Scholar] [CrossRef] [PubMed]
- Wi, S.J.; Ji, N.R.; Park, K.Y. Synergistic biosynthesis of biphasic ethylene and reactive oxygen species in response to hemibiotrophic Phytophthora parasitica in tobacco plants. Plant Physiol. 2012, 159, 251–265. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guimaraes, P.M.; Quintana, A.C.; Mota, A.P.Z.; Berbert, P.S.; Ferreira, D.d.S.; de Aguiar, M.N.; Pereira, B.M.; de Araújo, A.C.G.; Brasileiro, A.C.M. Engineering Resistance against Sclerotinia sclerotiorum Using a Truncated NLR (TNx) and a Defense-Priming Gene. Plants 2022, 11, 3483. https://doi.org/10.3390/plants11243483
Guimaraes PM, Quintana AC, Mota APZ, Berbert PS, Ferreira DdS, de Aguiar MN, Pereira BM, de Araújo ACG, Brasileiro ACM. Engineering Resistance against Sclerotinia sclerotiorum Using a Truncated NLR (TNx) and a Defense-Priming Gene. Plants. 2022; 11(24):3483. https://doi.org/10.3390/plants11243483
Chicago/Turabian StyleGuimaraes, Patricia Messenberg, Andressa Cunha Quintana, Ana Paula Zotta Mota, Pedro Souza Berbert, Deziany da Silva Ferreira, Matheus Nascimento de Aguiar, Bruna Medeiros Pereira, Ana Claudia Guerra de Araújo, and Ana Cristina Miranda Brasileiro. 2022. "Engineering Resistance against Sclerotinia sclerotiorum Using a Truncated NLR (TNx) and a Defense-Priming Gene" Plants 11, no. 24: 3483. https://doi.org/10.3390/plants11243483
APA StyleGuimaraes, P. M., Quintana, A. C., Mota, A. P. Z., Berbert, P. S., Ferreira, D. d. S., de Aguiar, M. N., Pereira, B. M., de Araújo, A. C. G., & Brasileiro, A. C. M. (2022). Engineering Resistance against Sclerotinia sclerotiorum Using a Truncated NLR (TNx) and a Defense-Priming Gene. Plants, 11(24), 3483. https://doi.org/10.3390/plants11243483