
Codonoblepharonteae, a New Major Lineage among Orthotrichoideae (Orthotrichaceae, Bryophyta)

 ,
,  ,
, 
Abstract
:1. Introduction
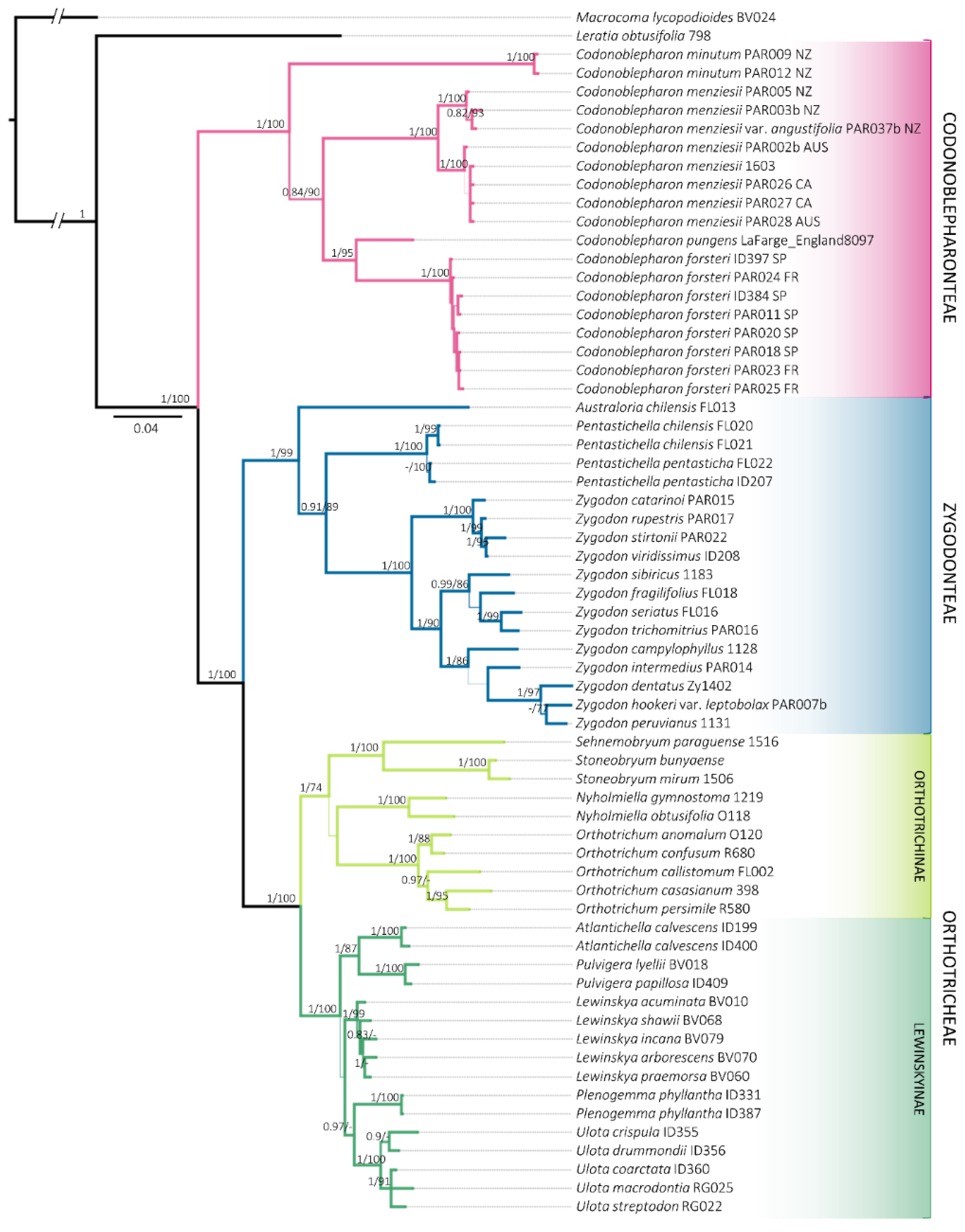
2. Results
3. Discussion
4. Materials and Methods
4.1. Taxon Sampling
4.2. DNA Isolation and Amplification
4.3. Molecular Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Nomenclature
| Nomenclatural Changes: |
| Codonoblepharonteae F. Lara, Garilleti and Draper, tribe nova |
| TYPE: Codonoblepharon Schwägr. Species Muscorum Frondosorum, Supplementum Secundum 2: 142. pl. 137 p.p. 1824. |
References
- Goffinet, B.; Buck, W.R.; Shaw, A.J. Morphology, anatomy, and classification of the Bryophyta. In Bryophyte Biology, 2nd ed.; Goffinet, B., Shaw, A.J., Eds.; Camb: Cambridge, UK, 2009; pp. 55–138. [Google Scholar]
- Medina, N.G.; Draper, I.; Lara, F. Biogeography of Mosses and Allies: Does Size Matter? In Biogeography of Microscopic Organisms; Fontaneto, D., Ed.; Cambridge University Press: Cambridge, UK, 2011; pp. 209–233. [Google Scholar] [CrossRef]
- Chang, Y.; Graham, S.W. Inferring the Higher-Order Phylogeny of Mosses (Bryophyta) and Relatives Using a Large, Multigene Plastid Data Set. Am. J. Bot. 2011, 98, 839–849. [Google Scholar] [CrossRef] [Green Version]
- Cox, C.J.; Goffinet, B.; Wickett, N.J.; Boles, S.B.; Shaw, A.J. Moss Diversity: A Molecular Phylogenetic Analysis of Genera. Phytotaxa 2014, 9, 175–195. [Google Scholar] [CrossRef]
- Liu, Y.; Johnson, M.G.; Cox, C.J.; Medina, R.; Devos, N.; Vanderpoorten, A.; Hedenäs, L.; Bell, N.E.; Shevock, J.R.; Aguero, B.; et al. Resolution of the Ordinal Phylogeny of Mosses Using Targeted Exons from Organellar and Nuclear Genomes. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goffinet, B.; Shaw, A.J.; Cox, C.J.; Wickett, N.J.; Boles, S.B. Phylogenetic Inferences in the Orthotrichoideae (Orthotrichaceae, Bryophyta) Based on Variation in Four Loci from All Genomes. Monogr. Syst. Bot. Missouri Bot. Gard. 2004, 98, 270–289. [Google Scholar]
- Frey, W.; Stech, M. Division of Bryophyta Schimp. (Musci, Mosses). In Syllabus of Plant Families. Adolf Engler’s Syllabus der Pflanzenfamilien, 13th edition. Part 3. Bryophytes and Seedless Vascular Plants; Frey, W., Ed.; Gebrüder Borntraeger: Berlin, Germany, 2009; pp. 116–257. [Google Scholar]
- Goffinet, B.; Vitt, D.H. Revised Generic Classification of the Orthotrichaceae Based on a Molecular Phylogeny and Comparative Morphology. In Bryology for the Twenty-first Century; Bates, J.W., Ashton, N.W., Duckett, J.G., Eds.; Maney Publishing and the British Bryological Society: Leeds, UK, 1998; pp. 143–160. [Google Scholar]
- Lara, F.; Garilleti, R.; Medina, R.; Mazimpaka, V. A New Key to the Genus Orthotrichum Hedw. in Europe and the Mediterranean Region. Cryptogamie Bryol. 2009, 30, 129–142. [Google Scholar]
- Sawicki, J.; Plášek, V.; Szczecińska, M. Molecular Data Do Not Support the Current Division of Orthotrichum (Bryophyta) Species with Immersed Stomata. J. Syst. Evol. 2012, 50, 12–24. [Google Scholar] [CrossRef]
- Lara, F.; Draper, I.; Flagmeier, M.; Calleja, J.A.; Mazimpaka, V.; Garilleti, R. Let’s Make Pulvigera Great Again: Re-Circumscription of a Misunderstood Group of Orthotrichaceae That Diversified in North America. Bot. J. Linn. Soc. 2020, 193, 180–206. [Google Scholar] [CrossRef]
- Draper, I.; Garilleti, R.; Calleja, J.A.; Flagmeier, M.; Mazimpaka, V.; Vigalondo, B.; Lara, F. Insights into the Evolutionary History of the Subfamily Orthotrichoideae (Orthotrichaceae, Bryophyta): New and Former Supra-Specific Taxa So Far Obscured by Prevailing Homoplasy. Front. Plant Sci. 2021, 12, 629035. [Google Scholar] [CrossRef]
- Draper, I.; Villaverde, T.; Garilleti, R.; Burleigh, J.G.; McDaniel, S.F.; Mazimpaka, V.; Calleja, J.A.; Lara, F. An NGS-Based Phylogeny of Orthotricheae (Orthotrichaceae, Bryophyta) With the Proposal of the New Genus Rehubryum from Zealandia. Front. Plant Sci. 2022, 13, 882960. [Google Scholar] [CrossRef]
- Matcham, H.W.; O’Shea, B.J. A Review of the Genus Codonoblepharon Schwägr. (Bryopsida: Orthotrichaceae). J. Bryol. 2005, 27, 129–135. [Google Scholar] [CrossRef]
- Malta, N. Die Gattung Zygodon Hook. et Tayl. Ein Monographischen Studie; Latvijas Universitates: Riga, Latvia, 1926. [Google Scholar]
- Calabrese, G.M. A Taxonomic Revision of Zygodon (Orthotrichaceae) in Southern South America. Bryologist 2006, 109, 453–509. [Google Scholar] [CrossRef]
- Lara, F.; Garilleti; Mazimpaka, V.; Guerra, J. Orthotrichaceae. In Flora Briofítica Ibérica, vol. V. Orthotrichales: Orthotrichaceae; Hedwigiales: Hedwigiaceae; Leucodontales: Fontinalaceae, Climaciaceae, Anomodontaceae, Cryphaeaceae, Leptodontaceae, Leucodontaceae, Neckeraceae; Hookeriales: Hypopterygiaceae, Hookeriaceae, Leucomiaceae, Pilotrichaceae; Guerra, J., Cano, M.J., Brugués, M., Eds.; Universidad de Murcia and Sociedad Española de Briología: Murcia, Spain, 2014; pp. 15–135. [Google Scholar]
- Goffinet, B. A Reconsideration of the Affinities of Kleioweisiopsis, Pleurozygodontopsis, Trigonodictyon, and the Microtheciellaceae (Bryopsida, Orthotrichales). J. Bryol. 1998, 20, 69–81. [Google Scholar] [CrossRef]
- Goffinet, B.; Bayer, R.J.; Vitt, D.H. Circumscription and Phylogeny of the Orthotrichales (Bryopsida) Inferred from RBCL Sequence Analyses. Am. J. Bot. 1998, 85, 1324–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schimper, W.P. Corollarium Bryologiae Europaeae; E. Schweizerbart: Stuttgart, Germany, 1856. [Google Scholar]
- Vitt, D.H. The Genera of Orthotrichaceae. Beih. Nova Hedwigia 1982, 71, 261–268. [Google Scholar]
- Shevock, J.R. Two Mosses New to North America with Southern Hemisphere Affinities. Evansia 2000, 17, 97–98. [Google Scholar] [CrossRef]
- Callaghan, D.A.; Aleffi, M.; Alegro, A.; Bisang, I.; Blockeel, T.L.; Collart, F.; Dragićević, S.; Draper, I.; Erdağ, A.; Erzberger, P.; et al. Global Geographical Range and Population Size of the Habitat Specialist Codonoblepharon forsteri (Dicks.) Goffinet in a Changing Climate. J. Bryol. 2022, 44, 35–50. [Google Scholar] [CrossRef]
- Lewinsky, J. Zygodon Hook. & Tayl. in Australasia: A Taxonomic Revision Including SEM-studies of Peristomes. Lindbergia 1989, 15, 109–139. [Google Scholar]
- Ziolkowski, P.A.; Sadowski, J. FISH-Mapping of RDNAs and Arabidopsis BACs on Pachytene Complements of Selected Brassicas. Genome 2002, 45, 189–197. [Google Scholar] [CrossRef]
- Nadot, S.; Bajon, R.; Lejeune, B. The Chloroplast Gene rps4 as a Tool for the Study of Poaceae Phylogeny. Plant Syst. Evol. 1994, 191, 27–38. [Google Scholar] [CrossRef]
- Souza-Chies, T.T.; Bittar, G.; Nadot, S.; Carter, L.; Besin, E.; Lejeune, B. Phylogenetic Analysis of Iridaceae with Parsimony and Distance Methods Using the Plastid Gene rps4. Plant Syst. Evol. 1997, 204, 109–123. [Google Scholar] [CrossRef]
- Vigalondo, B.; Lara, F.; Draper, I.; Valcárcel, V.; Garilleti, R.; Mazimpaka, V. Is It Really You, Orthotrichum acuminatum? Ascertaining a New Case of Intercontinental Disjunction in Mosses. Bot. J. Linn. Soc. 2016, 180, 30–49. [Google Scholar] [CrossRef] [Green Version]
- Werner, O.; Patiño, J.; González-Mancebo, J.M.; de Almeida Gabriel, R.M.; Ros, R.M. The Taxonomic Status and the Geographical Relationships of the Macaronesian Endemic Moss Fissidens luisieri (Fissidentaceae) Based on DNA Sequence Data. Bryologist 2009, 112, 315–324. [Google Scholar] [CrossRef] [Green Version]
- Pacak, A.; Szweykowska-Kulińska, Z. Molecular Data Concerning Alloploid Character and the Origin of Chloroplast and Mitochondrial Genomes in the Liverwort Pellia borealis. Plant Biotechnol. J. 2000, 2, 101–108. [Google Scholar]
- Müller, K.; Müller, J.; Neinhuis, C.; Quandt, D. PhyDE® (Phylogenetic Data Editor). 2006. Available online: http://www.phyde.de/ (accessed on 10 February 2022).
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI Search and Sequence Analysis Tools APIs in 2019. Nucleic Acids Res. 2019, 47, 636–641. [Google Scholar] [CrossRef] [PubMed]
- Simmons, M.P.; Ochoterena, H. Gaps as Characters in Sequence-Based Phylogenetic Analyses. Syst. Biol. 2000, 49, 369–381. [Google Scholar] [CrossRef] [Green Version]
- Müller, K. SeqState. Appl-Bioinformatics 2005, 4, 65–69. [Google Scholar] [CrossRef]
- Castresana, J. Selection of Conserved Blocks from Multiple Alignments for Their Use in Phylogenetic Analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Talavera, G.; Castresana, J. Improvement of Phylogenies after Removing Divergent and Ambiguously Aligned Blocks from Protein Sequence Alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. PartitionFinder: Combined Selection of Partitioning Schemes and Substitution Models for Phylogenetic Analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef] [Green Version]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New Methods for Selecting Partitioned Models of Evolution for Molecular and Morphological Phylogenetic Analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian Inference of Phylogenetic Trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian Phylogenetic Inference under Mixed Models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: Oxford, UK; New York, NY, USA, 2000. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree 1.4.4. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 3 June 2022).


{kind=link}
| Orthotrichoideae Broth. |
|---|
| Tribe Orthotricheae Engler |
| Subtribe Orthotrichinae F.Lara, Garilleti and Draper |
| Orthotrichum Hedw. |
| Sehnemobryum Lewinsky and Hedenäs |
| Stoneobryum D.H.Norris and H.Rob. |
| Nyholmiella Holmen and E.Warncke |
| Subtribe Lewinskyinae F.Lara, Garilleti and Draper |
| Atlantichella F.Lara, Garilleti and Draper |
| Lewinskya F.Lara, Garilleti and Goffinet |
| Pulvigera Plášek, Sawicki and Ochyra |
| Plenogemma Plášek, Sawicki and Ochyra |
| Rehubryum F.Lara, Garilleti and Draper |
| Ulota D.Mohr |
| Tribe Zygodonteae Engler |
| Australoria F.Lara, Garilleti and Draper |
| Codonoblepharon Schwägr. |
| Pentastichella Müll.Hal. ex Thér. |
| Zygodon Hook. and Taylor |
| ITS2 | rps4 | trnG | trnL-F | Total | ||
|---|---|---|---|---|---|---|
| Ingroup | Variable sites | 115 | 91 | 35 | 43 | 257 |
| Informative sites | 37 | 57 | 11 | 38 | 143 | |
| Indel informative sites | 8 | 5 | 2 | 9 | 24 | |
| Total | Variable sites | 319 | 195 | 146 | 163 | 820 |
| Informative sites | 228 | 135 | 91 | 99 | 550 | |
| Indel informative sites | 81 | 15 | 28 | 29 | 153 | |
| Positions in data matrix | 514 | 647 | 554 | 360 | 2065 | |
| Indels in data matrix | 145 | 28 | 52 | 42 | 267 | |
| Evolutionary models | HKY + I + G | HKY + G | HKY + I + G | HKY + G | GTR + I + G | |
| Malta, 1926 [15] | Goffinet and Vitt, 1998 [8] | Calabrese, 2006 [16] | Goffinet et al., 2004 [6] | Draper et al., 2021 [12] and Current Proposal |
|---|---|---|---|---|
| Zygodon sect. Euzygodon | Zygodon | Zygodon subg. Zygodon sect. Zygodon | Zygodon | Zygodon |
| Leptodontiopsis | -- | |||
| Zygodon sect. Stenomitrium | Stenomitrium1 | Zygodon subg. Zygodon sect. Stenomitrium 4 | Pentastichella | Pentastichella |
| – | Pleurorthotrichum2 | – | Pleurorthotrichum2 | |
| – | -- | – | -- | Australoria |
| Zygodon sect. Obtusifolii | Bryomaltaea | Zygodon subg. Obtusifolii | Leratia ** | Leratia ** |
| – | Leratia | – | ||
| Zygodon sect. Bryoides | Codonoblepharon3 | Zygodon subg. Codonoblepharon | Codonoblepharon | Codonoblepharon ** |
| Region | Primer name | Sequence | Authorship |
|---|---|---|---|
| ITS2 | ITS2-F | CGGATATCTTGGCTCTTG | Ziolkowski and Sadowski [25] |
| ITS2-R | CCGCTTAGTGATATGCTTA | ||
| rps4 | rpsA | ATGTCCCGTTATCGAGGACCT | Nadot et al. [26] |
| trnaS | TACCGAGGGTTCGAATC | Souza-Chies et al. [27] | |
| trnL-F | trnLc-104 | TAAGCAATCCTGAGC | Vigalondo et al. [28] |
| trnFF-425 | CTCTGCTCTACCAACT | ||
| trnG | trnGF-Leu | GGCTAAGGGTTATAGTCGGC | Werner et al. [29] |
| trnGr | GCGGGTATAGTTTAGTGG | Pacak and Szweykowska-Kulinska [30] |
| ITS2 | rps4 | trnG | trnL-F | ||||
|---|---|---|---|---|---|---|---|
| 1 | 94 °C 1 min | 1 | 94 °C 5 min | 1 | 94 °C 5 min | 1 | 95 °C 5 min |
| 30 | 94 °C 1 min | 30 | 95 °C 30 s | 40 | 94 °C 30 s | 38 | 94 °C 30 s |
| 59 °C 1 min | 52 °C 1 min | 52 °C 40 s | 47 °C 1 min | ||||
| 72 °C 1 min 30 s | 68 °C 30 s | 72 °C 1 min 30 s | 72 °C 30 s | ||||
| 94 °C 30 s | |||||||
| 1 | 72 °C 5 min | 1 | 68 °C 7 min | 1 | 72 °C 8 min | 1 | 72 °C 10 min |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguado-Ramsay, P.; Draper, I.; Garilleti, R.; Flagmeier, M.; Lara, F. Codonoblepharonteae, a New Major Lineage among Orthotrichoideae (Orthotrichaceae, Bryophyta). Plants 2022, 11, 3557. https://doi.org/10.3390/plants11243557
Aguado-Ramsay P, Draper I, Garilleti R, Flagmeier M, Lara F. Codonoblepharonteae, a New Major Lineage among Orthotrichoideae (Orthotrichaceae, Bryophyta). Plants. 2022; 11(24):3557. https://doi.org/10.3390/plants11243557
Chicago/Turabian StyleAguado-Ramsay, Pablo, Isabel Draper, Ricardo Garilleti, Maren Flagmeier, and Francisco Lara. 2022. "Codonoblepharonteae, a New Major Lineage among Orthotrichoideae (Orthotrichaceae, Bryophyta)" Plants 11, no. 24: 3557. https://doi.org/10.3390/plants11243557
APA StyleAguado-Ramsay, P., Draper, I., Garilleti, R., Flagmeier, M., & Lara, F. (2022). Codonoblepharonteae, a New Major Lineage among Orthotrichoideae (Orthotrichaceae, Bryophyta). Plants, 11(24), 3557. https://doi.org/10.3390/plants11243557