
Genome Size and Chromosome Number Evaluation of Astragalus L. sect. Hymenostegis Bunge (Fabaceae)


Abstract
:1. Introduction
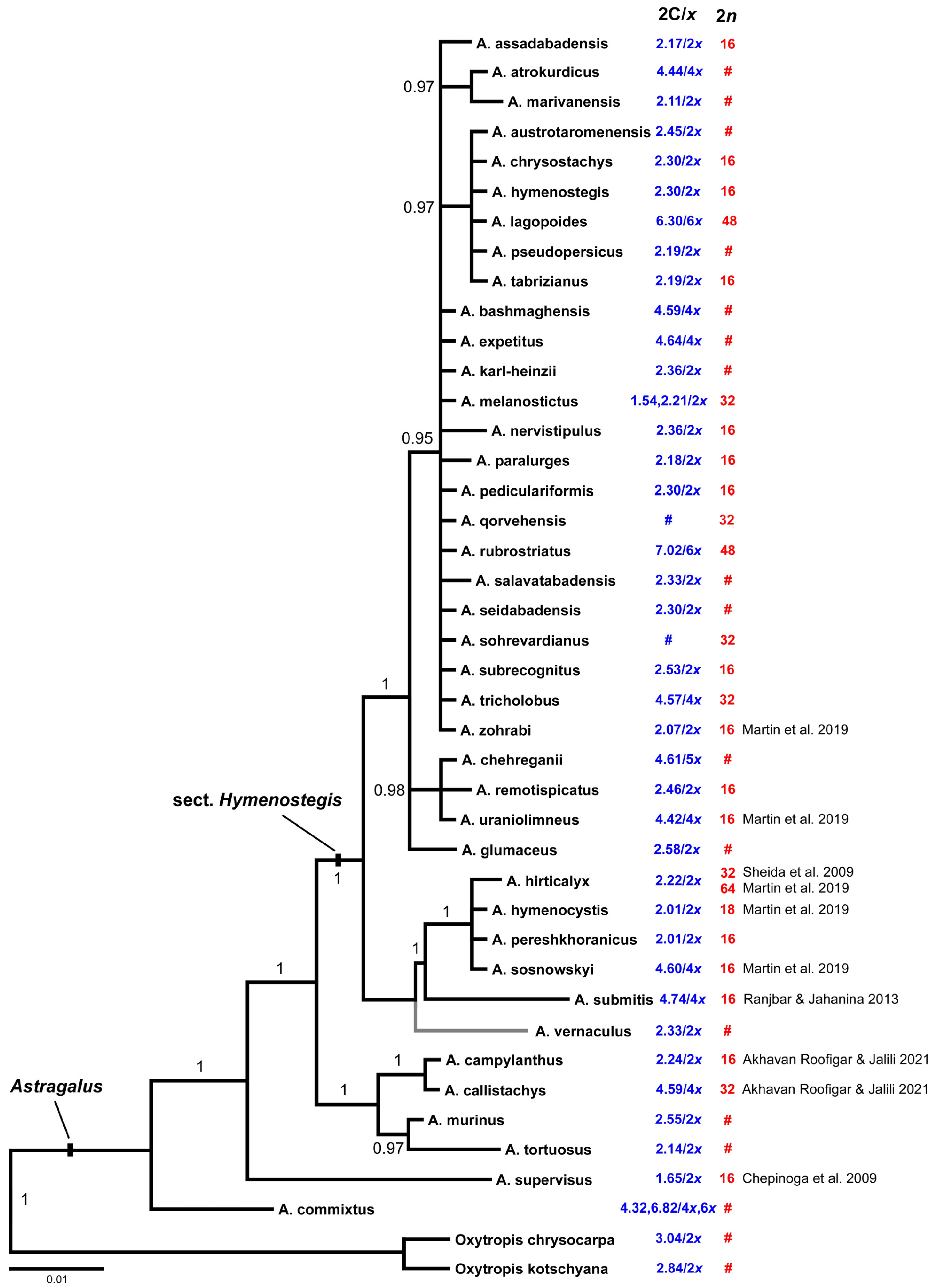
2. Results
2.1. Chromosome Numbers and Karyotypes
2.2. Genome Size Variation
2.3. Distribution of Genome Sizes among Phylogenetic Clades
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Chromosome Number and Karyotype Analysis
4.3. Genome Size Measurement
4.4. Phylogenetic Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lysak, M.A.; Berr, A.; Pecinka, A.; Schmidt, R.; McBreen, K.; Schubert, I. Mechanisms of chromosome number reduction in Arabidopsis thaliana and related Brassicaceae species. Proc. Natl. Acad. Sci. USA 2006, 103, 5224–5229. [Google Scholar] [CrossRef] [Green Version]
- Weiss-Schneeweiss, H.; Schneeweiss, G.M. Karyotype diversity and evolutionary trends in angiosperms. In Plant Genome Diversity 2: Physical Structure, Behaviour and Evolution of Plant Genomes; Springer: Vienna, Austria, 2013; Volume 2, pp. 209–230. [Google Scholar]
- Blattner, F.R. Phylogeny of Hordeum (Poaceae) as inferred by nuclear rDNA ITS sequences. Mol. Phylogenet. Evol. 2004, 33, 289–299. [Google Scholar] [CrossRef]
- Bennett, M.D.; Leitch, I.J. Nuclear DNA amounts in angiosperms: Targets, trends and tomorrow. Ann. Bot. 2011, 107, 467–590. [Google Scholar] [CrossRef]
- Jang, T.S.; Parker, J.S.; Emadzade, K.; Temsch, E.M.; Leitch, A.R.; Weiss-Schneeweiss, H. Multiple origins and nested cycles of hybridization result in high tetraploid diversity in the monocot Prospero. Front. Plant Sci. 2018, 9, 433. [Google Scholar] [CrossRef] [PubMed]
- Greilhuber, J. Intraspecific variation in genome size: A critical reassessment. Ann. Bot. 1998, 82 (Suppl. A), 27–35. [Google Scholar] [CrossRef] [Green Version]
- Soltis, D.E.; Soltis, P.S.; Bennett, M.D.; Leitch, I.J. Evolution of genome size in the angiosperms. Am. J. Bot. 2003, 90, 1596–1603. [Google Scholar] [CrossRef] [PubMed]
- Jakob, S.S.; Meister, A.; Blattner, F.R. The considerable genome size variation in Hordeum species (Poaceae) is linked to phylogeny, life form, ecology, and speciation rates. Mol. Biol. Evol. 2004, 21, 860–869. [Google Scholar] [CrossRef] [PubMed]
- Kellogg, E.A.; Bennetzen, J.L. The evolution of nuclear genome structure in seed plants. Am. J. Bot. 2004, 91, 1709–1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podlech, D.; Zarre, S. A Taxonomic Revision of the Genus Astragalus L. (Leguminosae) in the Old World, 1rd ed.; vols. 1–Naturhistorisches Museum Wien: Vienna, Austria, 2013; 2439p. [Google Scholar]
- Maassoumi, A.A. Old World Check-List of Astragalus, 1rd ed.; Research Institute of Forests and Rangeland: Tehran, Iran, 1998; 617p. [Google Scholar]
- Sanderson, M.J.; Wojciechowski, M.F. Improved bootstrap confidence limits in large-scale phylogenies, with an example from Neo-Astragalus (Leguminosae). Syst. Biol. 2000, 49, 671–685. [Google Scholar] [CrossRef] [Green Version]
- Wojciechowski, M.F. Astragalus (Fabaceae): A molecular phylogenetic perspective. Brittonia 2005, 57, 382–396. [Google Scholar] [CrossRef]
- Kazempour Osaloo, S.; Maassoumi, A.A.; Murakami, N. Molecular systematic of the genus Astragalus L. (Fabaceae): Phylogenetic analyses of nuclear ribosomal DNA internal transcribed spacers and chloroplast gene ndhF sequences. Plant Syst. Evol. 2003, 242, 1–32. [Google Scholar] [CrossRef]
- Kazempour Osaloo, S.; Maassoumi, A.A.; Murakami, N. Molecular systematics of the Old World Astragalus (Fabaceae) as inferred from nrDNA ITS sequence data. Brittonia 2005, 57, 367–381. [Google Scholar] [CrossRef]
- Azani, N.; Bruneau, A.; Wojciechowski, M.F.; Zarre, S. Molecular phylogenetics of annual Astragalus (Fabaceae) and its systematic implications. Bot. J. Linn. Soc. 2017, 184, 347–365. [Google Scholar] [CrossRef]
- Azani, N.; Bruneau, A.; Wojciechowski, M.F.; Zarre, S. Miocene climate change as a driving force for multiple origins of annual species in Astragalus (Fabaceae, Papilionoideae). Mol. Phylogenet. Evol. 2019, 137, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Duan, L.; Liu, P.; Liu, J.; Chang, Z.; Wen, J. Chloroplast phylogenomics and character evolution of eastern Asian Astragalus (Leguminosae): Tackling the phylogenetic structure of the largest genus of flowering plants in Asia. Mol. Phylogenet. Evol. 2021, 156, 107025. [Google Scholar] [CrossRef]
- Bagheri, A.; Maassoumi, A.A.; Rahiminejad, M.R.; Brassac, J.; Blattner, F.R. Molecular phylogeny and divergence times of Astragalus section Hymenostegis: An analysis of a rapidly diversifying species group in Fabaceae. Sci. Rep. 2017, 7, 14033. [Google Scholar] [CrossRef] [Green Version]
- Bagheri, A.; Rahiminejad, M.R.; Maassoumi, A.A. A new species of the genus Astragalus (Leguminosae-Papilionoideae) from Iran. Phytotaxa 2014, 178, 38–42. [Google Scholar] [CrossRef]
- Bagheri, A.; Maassoumi, A.A.; Rahiminejad, M.R.; Blattner, F.R. Molecular phylogeny and morphological analysis support a new species and new synonymy in Iranian Astragalus (Leguminosae). PLoS ONE 2016, 11, e0149726. [Google Scholar] [CrossRef] [Green Version]
- Manandhar, L.; Sakya, S.R. Cytotaxonomic studies in two species of Astragalus. J. Cytol. Genet. 2004, 5, 13–20. [Google Scholar]
- Badr, A.; Sharawy, S.M. Karyotype analysis and systematic relationships in the Egyptian Astragalus L. (Fabaceae). Int. J. Bot. 2007, 3, 147–159. [Google Scholar] [CrossRef] [Green Version]
- Martin, E.; Duran, A.; Dinç, M.; Erişen, S.; Babaoğlu, M. Karyotype analyses of four Astragalus L. (Fabaceae) species from Turkey. Phytologia 2008, 90, 147–159. [Google Scholar]
- Yousefzadeh, K.; Houshmand, S.; Zamani Dadane, G. Karyotype analysis of Astragalus effusus Bunge (Fabaceae). Caryologia 2010, 63, 257–261. [Google Scholar]
- Abdel Samad, F.; Baumel, A.; Juin, M.; Pavon, D.; Siljak-Yakovlev, S.; Médail, F.; Bou Dagher Kharrat, M. Phylogenetic diversity and genome sizes of Astragalus (Fabaceae) in the Lebanon biogeographical crossroad. Plant Syst. Evol. 2014, 300, 819–830. [Google Scholar] [CrossRef]
- Ledingham, G.F.; Fahselt, M.D. Chromosome numbers of some North American species of Astragalus (Leguminosae). Sida 1964, 1, 313–327. [Google Scholar]
- Ledingham, G.F.; Rever, B.M. Chromosome numbers of some Southwest Asian species of Astragalus and Oxytropis (Leguminosae). Canad. J. Genet. Cytol. 1963, 5, 18–32. [Google Scholar] [CrossRef]
- Wojciechowski, M.F.; Sanderson, M.J.; Hu, J.M. Evidence on the monophyly of Astragalus (Fabaceae) and its major subgroups based on nuclear ribosomal DNA ITS and chloroplast DNA trnL intron data. Syst. Bot. 1999, 24, 409–437. [Google Scholar] [CrossRef]
- Ghaffari, S.M. Index to Plant Chromosome Number of Iran, 1rd ed.; Research Institute of Forests and Rangeland: Tehran, Iran, 2020; 336p. [Google Scholar]
- Sheidai, M.; Gharemaninejad, F. New chromosome number and karyotype analysis in four Astragalus L. (Fabaceae) species. Iran. J. Bot. 2008, 15, 21–26. [Google Scholar]
- Sheidai, M.; Zarre, S.; Ismeilzadeh, J. New chromosome number reports in tragacanthic Astragalus species. Caryologia 2009, 62, 30–36. [Google Scholar]
- Bagheri, A.; Erkul, S.K.; Maassoumi, A.A.; Rahiminejad, M.R.; Blattner, F.R. Astragalus trifoliastrum (Fabaceae), a revived species for the flora of Turkey. Nord. J. Bot. 2015, 33, 532–539. [Google Scholar] [CrossRef]
- Martin, E.; Icyer Dogan, G.; Karaman Erkul, S.; Eroglu, H.E. Karyotype analyses of 25 Turkish taxa of Astragalus from the sections Macrophyllum, Hymenostegis, Hymenocoleus, and Anthyllis (Fabaceae). Turk. J. Bot. 2019, 43, 232–242. [Google Scholar] [CrossRef]
- Siljak-Yakovlev, S.; Pustahija, F.; Solic, E.M.; Bogunić, F.; Muratović, E.; Bašić, N.; Brown, S.C. Towards a genome size and chromosome number database of Balkan flora: C-values in 343 taxa with novel values for 242. Adv. Sci. Lett. 2010, 3, 190–213. [Google Scholar] [CrossRef]
- Temsch, E.M.; Temsch, W.; Ehrendorfer-Schratt, L.; Greilhuber, J. Heavy metal pollution, selection, and genome size: The species of the Žerjav Study revisited with flow cytometry. J. Bot. 2010, 15, 596542. [Google Scholar] [CrossRef] [Green Version]
- Bou Dagher-Kharrat, M.; Siljak-Yakovlev, S.; Abdel-Samad, N.; Douaihy, B.C.; Abdel-Samad, F.; Bourge, M.; Brown, S.C. Nuclear DNA C-values for biodiversity screening: Case of the Lebanese flora. Plant Biosyst. 2013, 147, 1228–1237. [Google Scholar] [CrossRef]
- Vallès, J.; Bašić, N.; Bogunić, F.; Bourge, M.; Brown, S.C.; Garnatje, T.; Hajrudinović, A.; Muratović, E.; Pustahija, F.; Šolić, E.M. Contribution to plant genome size knowledge: First assessments in five genera and 30 species of angiosperms from western Balkans. Bot. Serb. 2014, 38, 25–33. [Google Scholar]
- Ranjbar, M.; Assadi, A.; Karamian, R. Systematic study of Astragalus chrysostachys Boiss. (Fabaceae) in Iran, with the description of a new species. Ann. Nat. Hist. Mus. Wien Ser. B Bot. Zool. 2011, 112, 221–237. [Google Scholar]
- Ranjbar, M.; Jahanian, S. Cytotaxonomic study of Astragalus sect. Megalocystis (Fabaceae) in Iran. Cytologia 2013, 78, 181–193. [Google Scholar] [CrossRef] [Green Version]
- Akhavan Roofigar, A.; Jalili, A. A new chromosome number report in five endemic Astragalus L. (Fabaceae) species of Iran. Iran. J. Bot. 2021, 27, 177–181. [Google Scholar]
- Chepinoga, V.V.; Aleksandr, A.G.; Enushchenko, I.V.; Rosbakh, S.A. IAPT/IOPB chromosome data 8. Taxon 2009, 58, 1281–1314. [Google Scholar] [CrossRef]
- Brassac, J.; Blattner, F.R. Species-level phylogeny and polyploid relationships in Hordeum (Poaceae) inferred by next-generation sequencing and in silico cloning of multiple nuclear loci. Syst. Biol. 2015, 64, 792–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soltis, D.E.; Visger, C.J.; Marchant, D.B.; Soltis, P.S. Polyploidy: Pitfalls and paths to a paradigm. Am. J. Bot. 2016, 103, 1146–1166. [Google Scholar] [CrossRef] [Green Version]
- Levan, A.; Fredgra, K.; Sandberg, A.A. Nomenclature for centromeric position on chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Stebbins, G.L. Chromosomal Evolution in Higher Plants; Edward Arnold: London, UK, 1971; 216p. [Google Scholar]
- Arano, H.; Saito, H. Cytological studies in family Umbelliferae 5. Karyotypes of seven species in subtribe Seselinae. La Kromosomo 1980, 2, 471–480. [Google Scholar]
- Lysák, M.A.; Doležel, J. Estimation of nuclear DNA content in Sesleria (Poaceae). Caryologia 1998, 52, 123–132. [Google Scholar] [CrossRef]
- Farhat, P.; Hidalgo, O.; Robert, T.; Siljak-Yakovlev, S.; Leitch, I.J.; Adams, R.P.; Bou Dagher-Kharrat, M. Polyploidy in the conifer genus Juniperus: An unexpectedly high rate. Front. Plant Sci. 2019, 10, 676. [Google Scholar] [CrossRef] [Green Version]
- Farhat, P.; Siljak-Yakovlev, S.; Hidalgo, O.; Rushforth, K.; Bartel, J.A.; Valentin, N.; Leitch, I.; Adams, R.P. Polyploidy in Cupressaceae: Discovery of a new naturally occurring tetraploid, Xanthocyparis vietnamensis. J. Syst. Evol. 2022, 2, 28. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods); Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| No. | Species | Ind./plates | Chr. No. | CL 1 | q 2 | P 3 | TF% 4 | S% 5 | Ask% 6 | Syi% 7 | R 8 | Ci 9 | KF 10 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | A. assadabadensis | 7/11 | 2n = 2x = 16 | 3.4 | 1.44 | 1.95 | 42 | 74 | 57 | 74 | 1.38 | 0.42 | 1M + 6m + 1sm |
| 2 | A. chrysostachys | 3/5 | 2n = 2x = 16 | 1.47 | 0.66 | 0.81 | 45 | 61 | 55 | 81 | 1.26 | 0.43 | 2M + 5m + 1sm |
| 3 | A. hymenostegis | 10/14 | 2n = 2x = 16 | 2.08 | 0.89 | 1.19 | 43 | 69 | 57 | 75 | 1.34 | 0.42 | 8m |
| 4 | A. lagopoides | 5/9 | 2n = 6x = 48 | 1.74 | 0.72 | 1.02 | 41 | 61 | 58 | 70 | 1.44 | 0.41 | 1M + 21m + 2sm |
| 5 | A. melanostictus | 9/8 | 2n = 4x = 32 | 2.27 | 0.93 | 1.34 | 41 | 60 | 59 | 69 | 1.44 | 0.42 | 13m + 3sm |
| 6 | A. nervistipulus | 6/8 | 2n = 2x = 16 | 2.09 | 0.82 | 1.26 | 39 | 69 | 60 | 65 | 1.57 | 0.41 | 5m + 3sm |
| 7 | A. paralurges | 9/11 | 2n = 2x = 16 | 2.06 | 0.86 | 1.2 | 41 | 66 | 58 | 71 | 1.43 | 0.39 | 6m + 2sm |
| 8 | A. pediculariformis | 6/11 | 2n = 2x = 16 | 2.12 | 0.96 | 1.15 | 45 | 71 | 54 | 83 | 1.23 | 0.45 | 2M + 5m + 1sm |
| 9 | A. pereshkhoranicus | 6/9 | 2n = 2x = 16 | 2.32 | 0.88 | 1.43 | 38 | 80 | 62 | 61 | 1.65 | 0.41 | 6m + 2sm |
| 10 | A. qorvehensis | 8/9 | 2n = 4x = 32 | 1.79 | 0.77 | 1.02 | 43 | 52 | 57 | 75 | 1.34 | 0.41 | 2M + 12m + 2sm |
| 11 | A. remotispicatus | 10/8 | 2n = 2x = 16 | 2.26 | 0.95 | 1.3 | 42 | 47 | 58 | 73 | 1.39 | 0.43 | 1M + 6m + 1sm |
| 12 | A. rubrostriatus | 5/9 | 2n = 6x = 48 | 2.01 | 0.85 | 1.16 | 42 | 57 | 57 | 73 | 1.4 | 0.44 | 2M + 18m + 4sm |
| 13 | A. seidabadensis | 10/8 | 2n = 2x = 16 | 2.31 | 0.94 | 1.36 | 41 | 63 | 59 | 69 | 1.46 | 0.38 | 1M + 5m + 2sm |
| 14 | A. sohrevardianus | 7/10 | 2n = 4x = 32 | 1.91 | 0.79 | 1.12 | 41 | 50 | 58 | 71 | 1.40 | 0.36 | 15m + 1sm |
| 15 | A. subrecognitus | 10/7 | 2n = 2x = 16 | 3.5 | 1.41 | 2.8 | 40 | 53 | 59 | 68 | 1.48 | 0.38 | 2M + 3m + 3sm |
| 16 | A. tabrizianus | 5/7 | 2n = 2x = 16 | 2.75 | 0.99 | 1.76 | 36 | 59 | 63 | 56 | 1.77 | 0.38 | 3m + 5sm |
| 17 | A. tricholobus | 8/7 | 2n = 4x = 32 | 2.16 | 0.82 | 1.33 | 38 | 58 | 62 | 61 | 1.66 | 0.43 | 1M + 8m + 7sm |
| Species | Section | Locality | Geographical Coordinates | Elevation | Genome Size Standard | Mean Standard | Mean Sample | CV % Standard | CV % Sample | 2C Genome Size (pg) | GenBank Acc. No. ITS |
|---|---|---|---|---|---|---|---|---|---|---|---|
| A. assadabadensis F.Ghahrem. & Podlech * | Hymenostegis | Hamadan, Assadabad | 34°49′70″ N 48°10′90″ E | 2270 m | Maize | 103.79 | 41.55 | 2.89 | 7.39 | 2.17 | LT622371 |
| A. atrokurdicus Maassoumi, F.Ghahrem., Bagheri & Podlech | Hymenostegis | Zanjan, Zarand | 36°09′18″ N 48°30′78″ E | 2040 m | Maize | 100.46 | 82.66 | 4.11 | 3.93 | 4.47 | LT622374 |
| A. austrotaromensis Maassoumi, F.Ghahrem., Bagheri & Podlech 1 | Hymenostegis | Zanjan, Tarom | 36°45′74″ N 48°49′90″ E | 2150 m | Maize | 97.78 | 44.89 | 5.13 | 13.66 | 2.49 | LT622379 |
| A. austrotaromensis Maassoumi, F.Ghahrem., Bagheri & Podlech 2 | Hymenostegis | Zanjan, Tarom | 36°45′88″ N 48°48′86″ E | 2120 m | Maize | 98.71 | 44.07 | 6 | 9.29 | 2.42 | - |
| A. bashmaghensis Maassoumi & Podlech 1 | Hymenostegis | Kordestan, Marivan to Saqez | 36°12′08″ N 46°31′35″ E | 1800 m | Maize | 99.82 | 82.07 | 2.18 | 5.8 | 4.46 | LT622380 |
| A. bashmaghensis Maassoumi & Podlech 2 | Hymenostegis | Kordestan, Marivan to Saqez | 36°12′08″ N 46°31′35″ E | 1810 m | Maize | 99.76 | 86.54 | 2.32 | 6.3 | 4.71 | - |
| A. callistachys Buhse | Microphysa | Isfahan, Meymeh | 33°36′76″ N 50°59′79″ E | 2110 m | Maize | 105.69 | 89.41 | 2.86 | 4.22 | 4.59 | LT622397 |
| A. campylanthus Boiss. | Campylanthus | Isfahan, Boien | 33°10′65″ N 50°17′06″ E | 2835 m | Maize | 107.48 | 44.4 | 4.62 | 15.56 | 2.24 | LT622398 |
| A. chehreganii Zarre & Podlech | Hymenostegis | West Azerbaijan, Qoshchi | 38°02′14″ N 44°57′84″ E | 1660 m | Maize | 100.12 | 85 | 2.86 | 4.57 | 4.61 | LT622401 |
| A. chrysostachys Boiss. 1 * | Hymenostegis | Zanjan, Mahneshan | 36°49′86″ N 47°25′36″ E | 2090 m | Maize | 96.88 | 40.73 | 2.12 | 5.38 | 2.28 | LT622405 |
| A. chrysostachys Boiss. 2 | Hymenostegis | Zanjan, Mahneshan | 36°49′86″ N 47°25′36″ E | 2095 m | Maize | 95.07 | 39.75 | 2.25 | 3.64 | 2.27 | - |
| A. chrysostachys Boiss. 3 | Hymenostegis | East Azerbaijan, Saeidabad | 37°54′22″ N 46°32′01″ E | 2010 m | Maize | 102.08 | 42.77 | 2.63 | 4.62 | 2.28 | - |
| A. chrysostachys Boiss. 4 | Hymenostegis | Zanjan, Mahneshan | 36°49′86″ N 47°25′36″ E | 2085 m | Maize | 92.23 | 40.35 | 9.1 | 6.22 | 2.38 | - |
| A. commixtus Bunge 1 | Ankylotus | Zanjan, Abhar | 36°07′01″ N 49°03′56″ E | 1910 m | Maize | 71.75 | 90.1 | 5.66 | 6.54 | 6.82 | AB051925 |
| A. commixtus Bunge 2 | Ankylotus | Zanjan, Abhar | 36°07′01″ N 49°03′56″ E | 1912 m | Maize | 90.1 | 71.75 | 6.54 | 5.66 | 4.32 | - |
| A. expetitus Maassoumi | Hymenostegis | Zanjan, Anguran | 36°36′16″ N 47°20′07″ E | 2460 m | Maize | 107.56 | 91.85 | 2.59 | 4.58 | 4.64 | LT622430 |
| A. glumaceus Boiss. 1 | Hymenostegis | Zanjan, Mahneshan | 36°51′43″ N 47°26′17″ E | 1910 m | Maize | 94.97 | 45.5 | 3.2 | 7.18 | 2.60 | LT622439 |
| A. glumaceus Boiss. 2 | Hymenostegis | Kordestan, Sonateh | 36°10′60″ N 46°33′71″ E | 1510 m | Maize | 92.6 | 43.69 | 8.5 | 9.32 | 2.56 | - |
| A. hirticalyx Boiss. & Kotschy 1 | Hymenostegis | West Azerbaijan, Khoy to Chaldoran | 38°40′45″ N 44°41′99″ E | 1915 m | Maize | 98.46 | 41.49 | 3.01 | 14 | 2.29 | LT622447 |
| A. hirticalyx Boiss. & Kotschy 2 | Hymenostegis | West Azerbaijan, Chaldoran | 38°40′45″ N 44°41′99″ E | 1905 m | Maize | 99.01 | 39.38 | 4.06 | 10.32 | 2.16 | - |
| A. hymenocystis Fisch. & C.A.Mey. | Hymenostegis | West Azerbaijan, Qoshchi | 38°02′14″ N 44°57′84″ E | 1660 m | Maize | 98.2 | 36.28 | 4.37 | 9.4 | 2.01 | LT622454 |
| A. hymenostegis Fisch. & C.A.Mey. 1* | Hymenostegis | West Azerbaijan, Hashtian to Salmas | 38°01′39″ N 44°47′67″ E | 1800 m | Maize | 97.38 | 39.64 | 2.81 | 8.86 | 2.21 | LT622455 |
| A. hymenostegis Fisch. & C.A.Mey. 2 | Hymenostegis | West Azerbaijan, Qoshchi | 38°02′14″ N 44°57′84″ E | 1670 m | Maize | 100.09 | 43.8 | 2.91 | 7.73 | 2.38 | - |
| A. karl-heinzii Maassoumi 1 | Hymenostegis | Zanjan, Zarand | 36°09′07″ N 48°30′88″ E | 2090 m | Maize | 98.62 | 42.52 | 2.63 | 5.39 | 2.34 | KT997422 |
| A. karl-heinzii Maassoumi 2 | Hymenostegis | Zanjan, Zarand | 36°09′07″ N 48°30′88″ E | 2092 m | Maize | 98.22 | 42.92 | 3.09 | 12.49 | 2.37 | - |
| A. lagopoides Lam. * | Hymenostegis | East Azerbaijan, Kuhkamar | 38°37′22″ N 45°53′37″ E | 2080 m | Maize | 101.34 | 117.52 | 2.9 | 4.73 | 6.30 | LT622474 |
| A. marivanensis Podlech & Maassoumi | Hymenostegis | Kordestan, Marivan to Saqez | 36°09′44″ N 46°19′30″ E | 1730 m | Maize | 95.93 | 37.21 | 2.71 | 5.35 | 2.11 | LT622496 |
| A. melanostictus Freyn 1 * | Hymenostegis | Zanjan, Mahneshan | 36°39′44″ N 47°36′12″ E | 2160 m | Maize | 96.98 | 27.46 | 2.31 | 6.23 | 1.54 | LT622503 |
| A. melanostictus Freyn 2 | Hymenostegis | Zanjan, Dandi to Tekab | 36°35′26″ N 47°31′28″ E | 1960 m | Maize | 94.33 | 38.41 | 2.74 | 6.74 | 2.21 | - |
| A. murinus Boiss. | Anthylloidei | Isfahan, Boien | 33°10′65″ N 50°17′06″ E | 2800 m | Maize | 109.86 | 51.54 | 4.48 | 8.13 | 2.55 | LT622519 |
| A. nervistipulus Boiss. & Hausskn. * | Hymenostegis | Kordestan, Sanandaj to Marivan | 35°25′03″ N 46°51′34″ E | 2040 m | Maize | 101.86 | 44.28 | 4.09 | 9.96 | 2.36 | LT622524 |
| A. paralurges Bunge 1 * | Hymenostegis | Zanjan, Qeydar | 36°07′19″ N 48°32′48″ E | 2400 m | Maize | 101.33 | 40.97 | 2.89 | 7.52 | 2.20 | LT622534 |
| A. paralurges Bunge 2 | Hymenostegis | Zanjan, Zarand | 36°09′07″ N 48°30′88″ E | 2090 m | Maize | 103 | 41.03 | 2.93 | 8.64 | 2.16 | - |
| A. pediculariformis Maassoumi 1 * | Hymenostegis | Zanjan, Qeydar | 36°07′19″ N 48°32′48″ E | 2402 m | Maize | 99.17 | 42.6 | 2.19 | 15.63 | 2.33 | LT622543 |
| A. pediculariformis Maassoumi 2 | Hymenostegis | Zanjan, Qeydar | 36°07′19″ N 48°32′48″ E | 2405 m | Maize | 101.82 | 42.4 | 2.41 | 5.63 | 2.26 | - |
| A. pereshkhoranicus Maassoumi & F.Ghahrem. * | Hymenostegis | West Azerbaijan, Mavana to Serow | 37°36′74″ N 44°47′66″ E | 1500 m | Maize | 99 | 36.63 | 3.37 | 13.81 | 2.01 | LT622550 |
| A. pseudopersicus Podlech & Maassoumi 1 | Hymenostegis | East Azerbaijan, Sofian | 38°15′78″ N 45°51′11″ E | 1545 m | Maize | 108.18 | 43.46 | 2.35 | 15.41 | 2.18 | LT622560 |
| A. pseudopersicus Podlech & Maassoumi 2 | Hymenostegis | East Azerbaijan, Sofian | 38°15′78″ N 45°51′11″ E | 1550 m | Maize | 108.56 | 43.93 | 2.54 | 6.95 | 2.20 | - |
| A. qorvehensis Podlech * | Hymenostegis | Hamadan, Qorveh | 34°49′70″ N 48°10′90″ E | 2280 m | - | - | - | - | - | - | LT622565 |
| A. remotispicatus Bagheri & Maassoumi * | Hymenostegis | Zanjan, Zarand | 36°09′07″ N 48°30′88″ E | 2085 m | Maize | 101.27 | 45.85 | 2.51 | 10.75 | 2.46 | KT997427 |
| A. rubrostriatus Bunge 1 * | Hymenostegis | Zanjan, Tarom | 36°45′74″ N 48°49′90″ E | 2140 m | Maize | 104.93 | 132.77 | 4.12 | 5.63 | 6.87 | LT622579 |
| A. rubrostriatus Bunge 2 | Hymenostegis | Zanjan, Mahneshan | 36°49′86″ N 47°25′36″ E | 2090 m | Maize | 102.25 | 134.82 | 5.92 | 6.4 | 7.16 | - |
| A. salavatabadensis Podlech 1 | Hymenostegis | Kordestan, Salavatabad | 35°16′50″ N 47°08′40″ E | 2030 m | Maize | 97.82 | 42.51 | 2.58 | 14.68 | 2.36 | LT622589 |
| A. salavatabadensis Podlech 2 | Hymenostegis | Kordestan, Salavatabad | 35°16′50″ N 47°08′40″ E | 2040 m | Maize | 98.35 | 41.68 | 2.67 | 8.35 | 2.30 | - |
| A. seidabadensis Bunge 1 * | Hymenostegis | East Azerbaijan, Kandowan | 37°53′01″ N 46°11′22″ E | 1750 m | Maize | 107.19 | 45.89 | 3.17 | 10.13 | 2.32 | LT622595 |
| A. seidabadensis Bunge 2 | Hymenostegis | East Azerbaijan, Kandowan | 37°53′01″ N 46°11′22″ E | 1758 m | Maize | 100.86 | 43.84 | 3.2 | 12.04 | 2.36 | - |
| A. seidabadensis Bunge 3 | Hymenostegis | East Azerbaijan, Kandowan | 37°53′01″ N 46°11′22″ E | 1755 m | Maize | 99.1 | 40.61 | 6.14 | 7.77 | 2.23 | - |
| A. sohrevardianus Bagheri, Maassoumi & F.Ghahrem. * | Hymenostegis | Zanjan, Sohrevard | 36°05′33″ N 48°28′78″ E | 2020 m | - | - | - | - | - | - | LT622598 |
| A. sosnowskyi Grossh. 1 | Hymenostegis | Kordestan, Bukan to Mahabad | 36°12′50″ N 46°31′40″ E | 1800 m | Maize | 101.12 | 87.27 | 2.45 | 5.66 | 4.69 | LT622602 |
| A. sosnowskyi Grossh. 2 | Hymenostegis | Kordestan, Bukan to Mahabad | 36°12′50″ N 46°31′40″ E | 1810 m | Maize | 97.17 | 80.67 | 2.96 | 4.77 | 4.51 | - |
| A. submitis Boiss. | Hymenostegis/Anthylloidei | Zanjan, Zarand | 36°09′07″ N 48°30′88″ E | 2090 m | Maize | 96.81 | 84.59 | 2.17 | 3.91 | 4.74 | LT622611 |
| A. subrecognitus Bagheri, Maassoumi & F.Ghahrem. 1 * | Hymenostegis | Zanjan, Mahneshan to Pari | 36°51′43″ N 47°26′17″ E | 1900 m | Maize | 103.36 | 49.07 | 2.36 | 7.48 | 2.58 | - |
| A. subrecognitus Bagheri, Maassoumi & F.Ghahrem. 2 | Hymenostegis | Zanjan, Mahneshan to Pari | 36°51′43″ N 47°26′17″ E | 1905 m | Maize | 100.5 | 45.96 | 2.54 | 7.77 | 2.48 | LT622615 |
| A. supervisus (Kuntze) E.Sheld. | Incani | Zanjan, Zarand | 36°09′07″ N 48°30′88″ E | 2080 m | Maize | 96.85 | 29.51 | 3.05 | 5.71 | 1.65 | AB231116 |
| A. tabrizianus Fisch. * | Hymenostegis | Kordestan, Bukan to Mahabad | 36°45′22″ N 45°52′40″ E | 1930 m | Maize | 99.65 | 40.18 | 3.5 | 11.97 | 2.19 | LT622619 |
| A. tortuosus DC. | Anthylloidei | Kordestan, Sanandaj to Marivan | 35°25′03″ N 46°51′34″ E | 2045 m | Maize | 100.76 | 39.76 | 2.72 | 5.77 | 2.14 | AB908451 |
| A. tricholobus DC. 1 * | Hymenostegis | Zanjan, Qeydar | 36°07′19″ N 48°32′48″ E | 2400m | Maize | 100.12 | 83.07 | 2.05 | 3.69 | 4.51 | LT622623 |
| A. tricholobus DC. 2 | Hymenostegis | Zanjan, Mahneshan | 36°49′86″ N 47°25′36″ E | 2085 m | Maize | 106.19 | 91.9 | 2.18 | 6.95 | 4.70 | - |
| A. tricholobus DC. 3 | Hymenostegis | Zanjan, Abhar | 36°07′01″ N 49°03′56″ E | 1910 m | Maize | 99.62 | 81.91 | 2.31 | 5.73 | 4.46 | - |
| A. tricholobus DC. 4 | Hymenostegis | Zanjan, Mahneshan to Pari | 36°51′43″ N 47°26′17″ E | 1900 m | Maize | 105.11 | 89.07 | 2.48 | 4.3 | 4.60 | - |
| A. uraniolimneus Boiss. | Hymenostegis | East Azerbaijan, Kuhkamar | 38°38′58″ N 45°55′14″ E | 2280 m | Maize | 97.09 | 79.09 | 2.42 | 3.29 | 4.42 | LT622627 |
| A. vernaculus Podlech 1 | Hymenostegis | Isfahan, Boien | 33°10′65″ N 50°17′04″ E | 2800 m | Maize | 103.65 | 43.36 | 2.84 | 13.59 | 2.27 | LT622633 |
| A. vernaculus Podlech 2 | Hymenostegis | Isfahan, Dehrajab | 33°09′30″ N 50°16′91″ E | 2730 m | Maize | 96.61 | 42.38 | 3.09 | 6.58 | 2.38 | - |
| A. zohrabi Bunge | Hymenostegis | West Azerbaijan, Hashtian to Salmas | 38°01′39″ N 44°47′67″ E | 1805 m | Maize | 100.14 | 38.14 | 3.4 | 7.73 | 2.07 | LT622637 |
| Oxytropis chrysocarpa Boiss. | - | Isfahan, Fereydunshahr | 32°56′14″ N 50°02′94″ E | 2920 m | Maize | 106.11 | 59.34 | 3.1 | 5.99 | 3.04 | LC213337 |
| Oxytropis kotschyana Boiss. & Hohen. | - | Zanjan, Qeydar | 36°07′19″ N 48°32′48″ E | 2405 m | Maize | 105.35 | 55.09 | 3.8 | 5.49 | 2.84 | LT622640 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bagheri, A.; Akhavan Roofigar, A.; Nemati, Z.; Blattner, F.R. Genome Size and Chromosome Number Evaluation of Astragalus L. sect. Hymenostegis Bunge (Fabaceae). Plants 2022, 11, 435. https://doi.org/10.3390/plants11030435
Bagheri A, Akhavan Roofigar A, Nemati Z, Blattner FR. Genome Size and Chromosome Number Evaluation of Astragalus L. sect. Hymenostegis Bunge (Fabaceae). Plants. 2022; 11(3):435. https://doi.org/10.3390/plants11030435
Chicago/Turabian StyleBagheri, Ali, Azadeh Akhavan Roofigar, Zahra Nemati, and Frank R. Blattner. 2022. "Genome Size and Chromosome Number Evaluation of Astragalus L. sect. Hymenostegis Bunge (Fabaceae)" Plants 11, no. 3: 435. https://doi.org/10.3390/plants11030435
APA StyleBagheri, A., Akhavan Roofigar, A., Nemati, Z., & Blattner, F. R. (2022). Genome Size and Chromosome Number Evaluation of Astragalus L. sect. Hymenostegis Bunge (Fabaceae). Plants, 11(3), 435. https://doi.org/10.3390/plants11030435