
Exogenously Applied Trehalose Augments Cadmium Stress Tolerance and Yield of Mung Bean (Vigna radiata L.) Grown in Soil and Hydroponic Systems through Reducing Cd Uptake and Enhancing Photosynthetic Efficiency and Antioxidant Defense Systems



 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Growth Traits
2.3. Measurement of Photosynthetic Pigments Contents in Soil-Grown Plants
2.4. Determination of Relative Water Content (RWC) in Soil-Grown Plants
2.5. Determination of Electrolyte Leakage (EL), Malondialdehyde (MDA) and Hydrogen Peroxide (H2O2) Contents in Soil-Grown Plants
2.6. Determination of the Total Soluble Proteins (TSP) and Free Amino Acids (FAA) in Soil-Grown Plants
2.7. Antioxidant Activities Assay in Soil-Grown Plants
2.8. Determination of Yield Components in Soil-Grown Plants
2.9. Determination of Cadmium Concentration in Organs of Soil-Grown Plants
2.10. Statistical Analysis
3. Results
3.1. Growth Traits
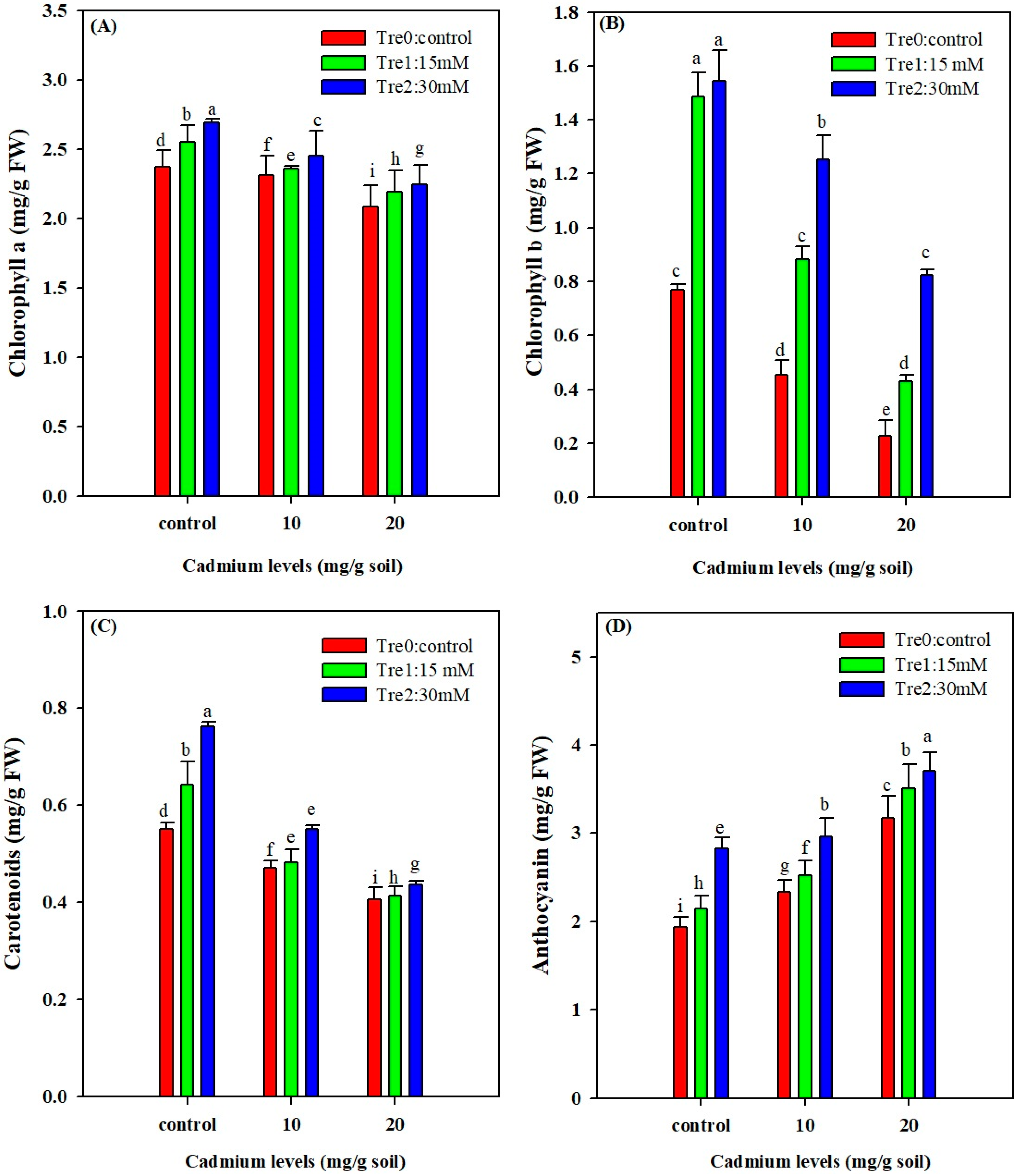
3.2. Photosynthetic Pigments and Anthocyanin Contents
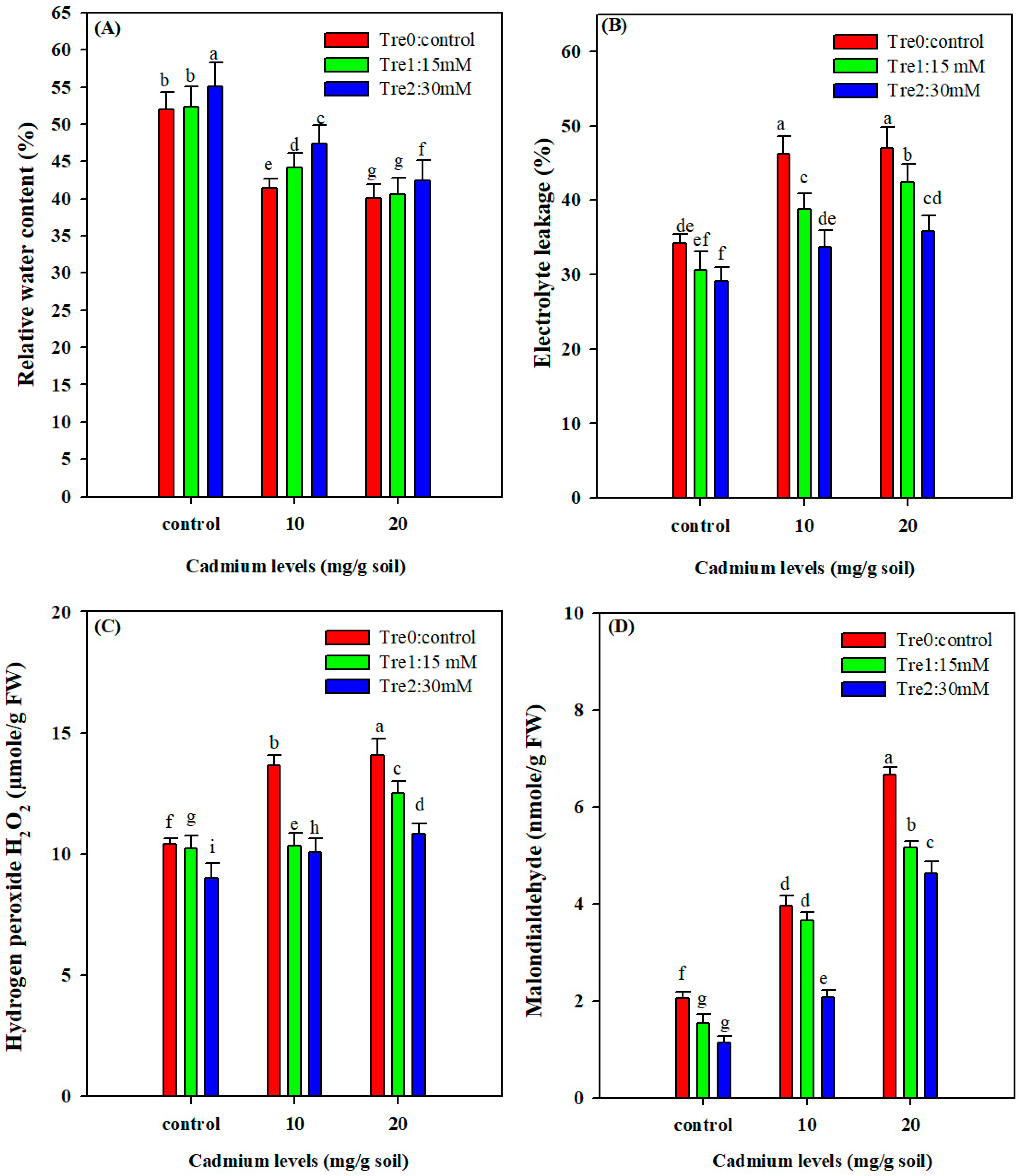
3.3. Relative Water Content
3.4. Electrolyte Leakage, MDA and H2O2
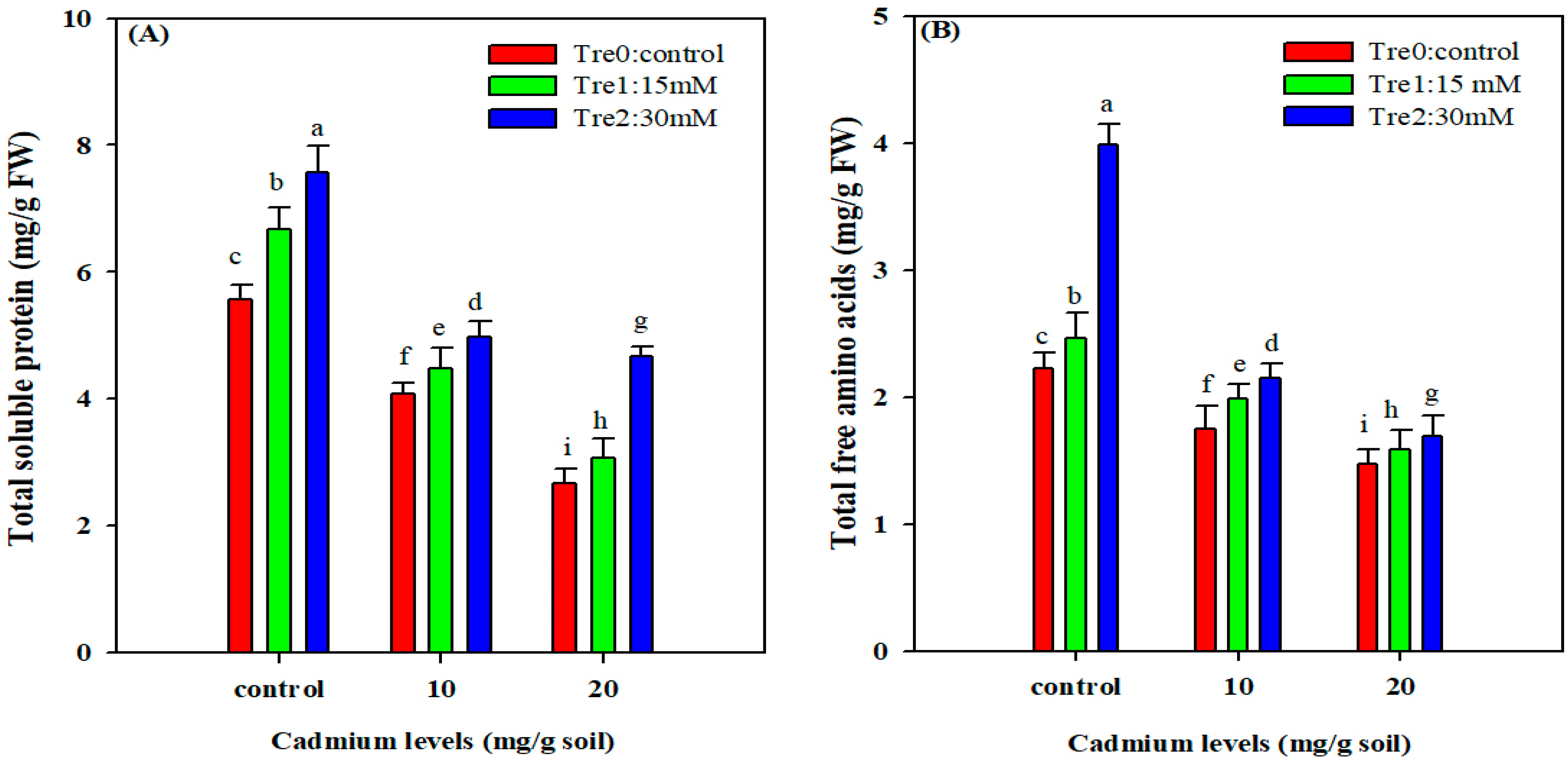
3.5. Total Soluble Proteins and Free Amino Acids
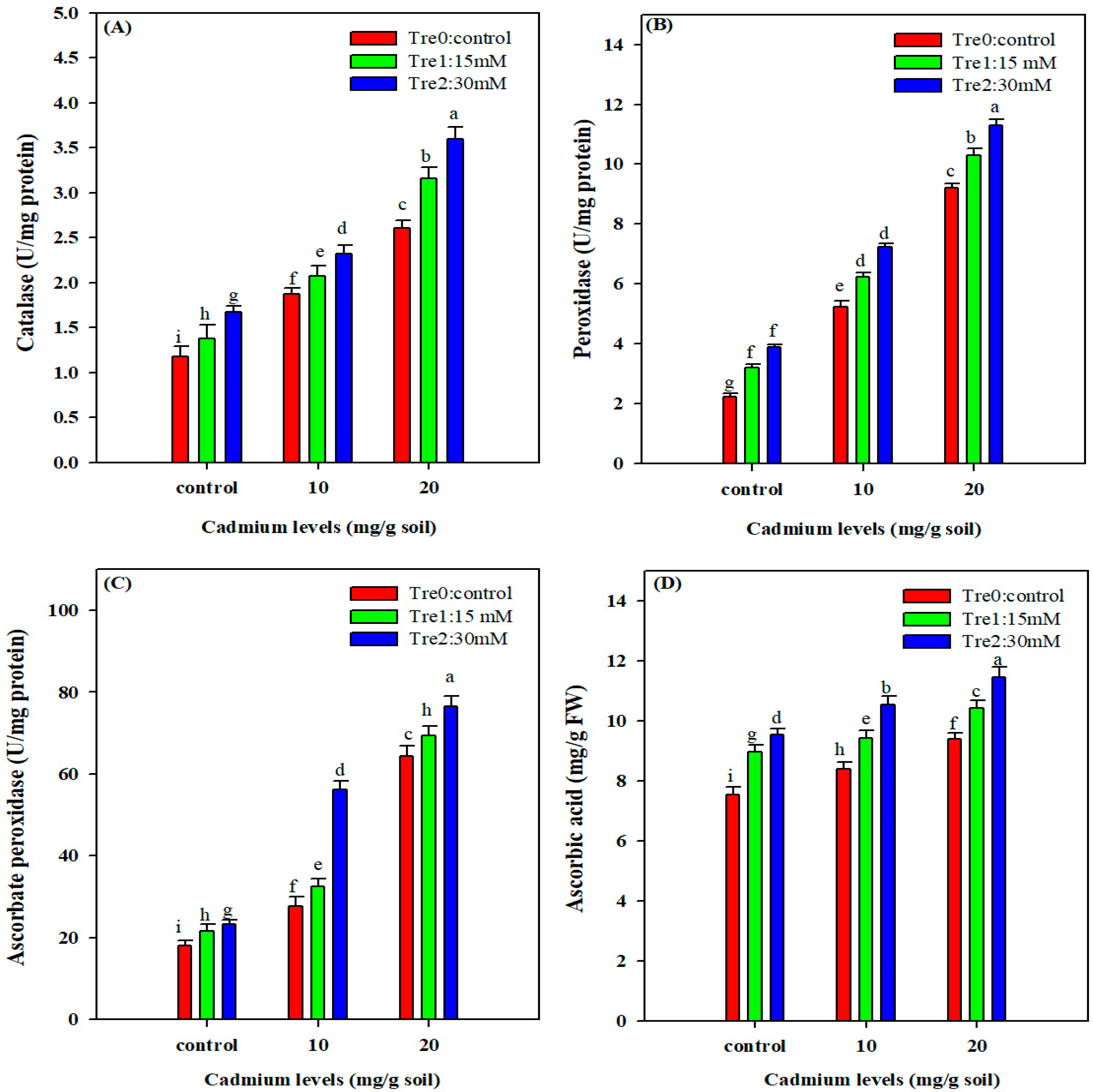
3.6. Antioxidant Enzymes Activities
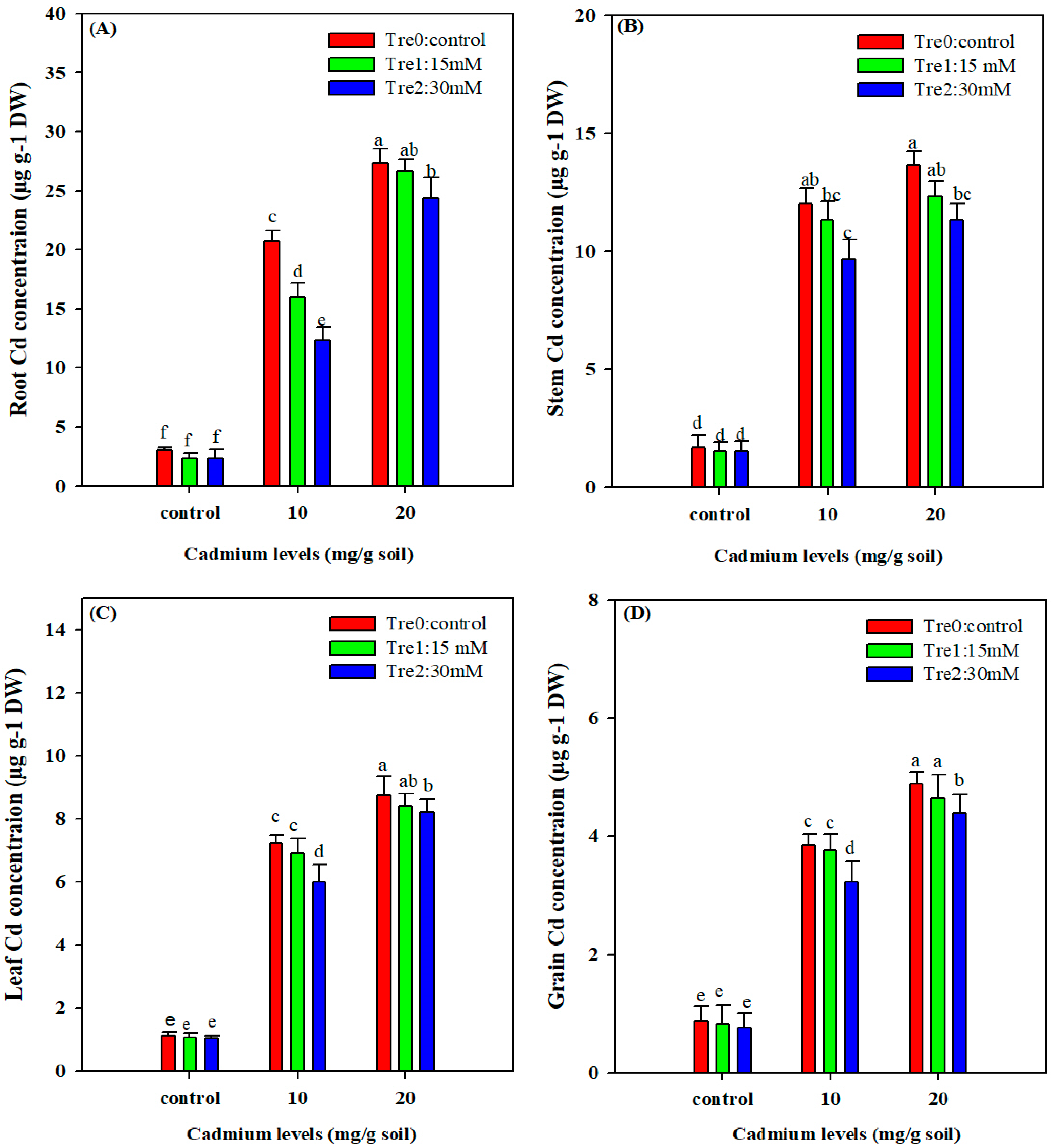
3.7. Cd Concentration in Different Plant Organs
3.8. Yield Components
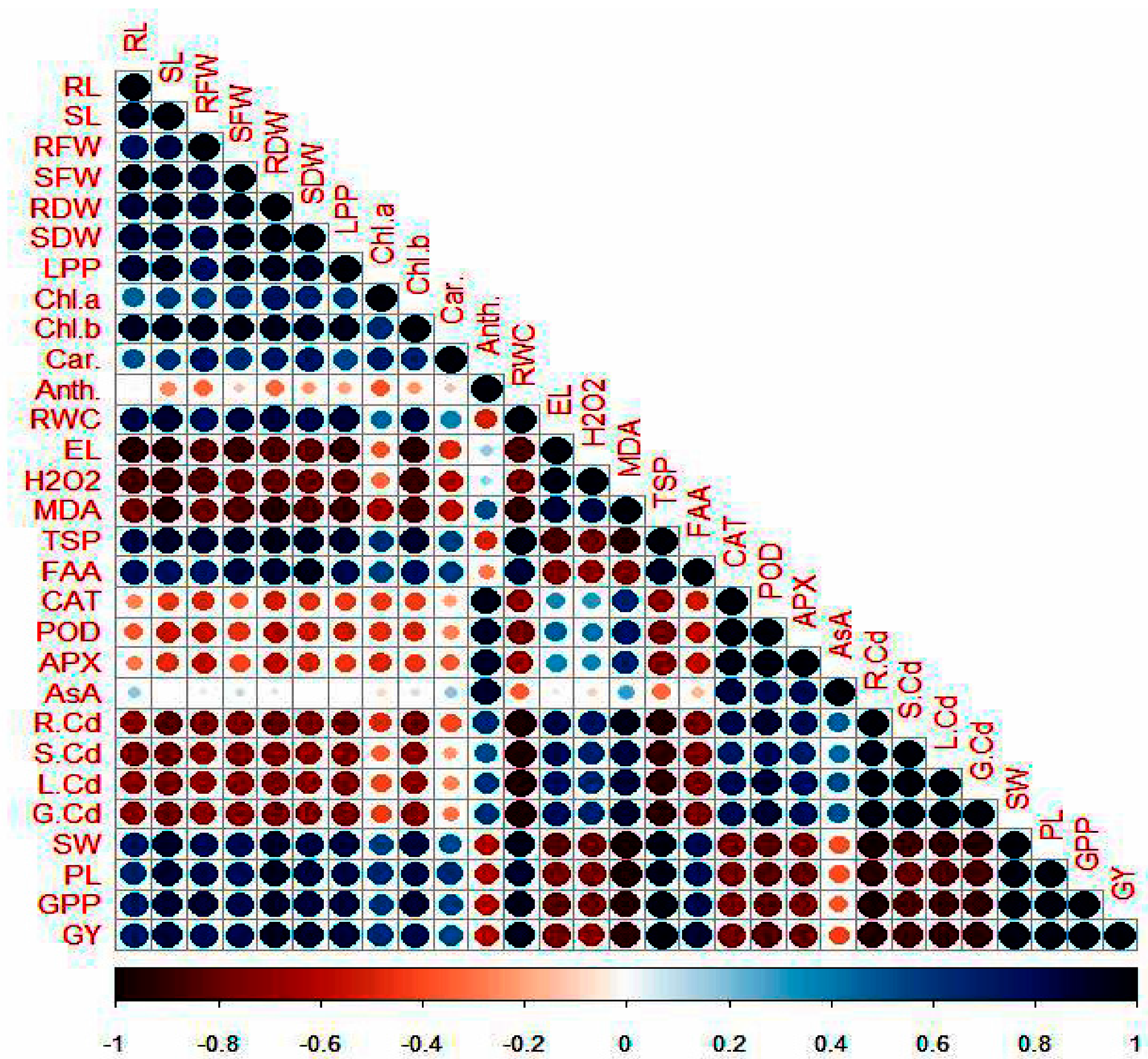
3.9. Pearson’s Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hassan, M.U.; Chattha, M.U.; Khan, I.; Chattha, M.B.; Aamer, M.; Nawaz, M.; Ali, A.; Khan, M.A.U.; Khan, T.A. Nickel toxicity in plants: Reasons, toxic effects, tolerance mechanisms, and remediation possibilities—A review. Environ. Sci. Poll. Res. 2019, 26, 12673–12688. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Guan, J.; Liang, Q.; Zhang, X.; Hu, H.; Zhang, J. Effects of cadmium stress on growth and physiological characteristics of sassafras seedlings. Sci. Rep. 2021, 11, 9913. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.; Qian, P.; Jin, R.; Ali, S.; Khan, M.; Aziz, R.; Tian, T.; Zhou, W. Physiological and ultra-structural changes in Brassica napus seedlings induced by cadmium stress. Biol. Plant. 2014, 58, 131–138. [Google Scholar] [CrossRef]
- Tang, Y.; Xie, Y.; Sun, G.; Tan, H.; Lin, L.; Li, H.; Liao, M.A.; Wang, Z.; Lv, X.; Liang, D.; et al. Cadmium-accumulator straw application alleviates cadmium stress of lettuce (Lactuca sativa) by promoting photosynthetic activity and antioxidative enzyme activities. Environ. Sci. Poll. Res. 2018, 25, 30671–30679. [Google Scholar] [CrossRef]
- Aamer, M.; Muhammad, U.H.; Li, Z.; Abid, A.; Su, Q.; Liu, Y.; Adnan, R.; Muhammad, A.U.K.; Tahir, A.K.; Huang, G. Foliar application of glycinebetaine (GB) alleviates the cadmium (Cd) toxicity in spinach through reducing Cd uptake and improving the activity of anti-oxidant system. Appl. Ecol. Environ. Res. 2018, 16, 7575–7583. [Google Scholar] [CrossRef]
- Chattha, M.U.; Arif, W.; Khan, I.; Soufan, W.; Bilal, M.C.; Hassan, M.U.; Ullah, N.; Sabagh, A.E.; Qari, S.H. Mitigation of cadmium induced oxidative stress by using organic amendments to improve the growth and yield of mash beans [Vigna mungo (L.)]. Agronomy 2021, 11, 2152. [Google Scholar] [CrossRef]
- Rasheed, A.; Fahad, S.; Aamer, M.; Hassan, M.U.; Tahir, M.M.; Wu, Z. Role of genetic factors in regulating cadmium uptake, transport and accumulation mechanisms and quantitative trait loci mapping in rice. a review. Appl. Ecol. Environ. Res. 2020, 18, 4005–4023. [Google Scholar] [CrossRef]
- Hussain, S.; Irfan, M.; Sattar, A.; Hussain, S.; Ullah, S.; Abbas, T.; Ur-Rehman, H.; Nawaz, F.; Al-Hashimi, A.; Elshikh, M.S.; et al. Alleviation of cadmium stress in wheat through the combined application of boron and biochar via regulating morpho-physiological and antioxidant defense mechanisms. Agronomy 2022, 12, 434. [Google Scholar] [CrossRef]
- Chen, H.C.; Zhang, S.L.; Wu, K.J.; Li, R.; He, X.R.; He, D.N.; Huang, C.; Wei, H. The effects of exogenous organic acids on the growth, photosynthesis and cellular ultrastructure of Salix variegata Franch. Under Cd stress. Ecotoxicol. Environ. Saf. 2020, 187, 109790. [Google Scholar] [CrossRef]
- Imran, K.; Seleiman, M.F.; Chattha, M.U.; Jalal, R.S.; Mahmood, F.; Hassan, F.A.; Izzet, W.; Alhammad, B.A.; Rana, R.; Hassan, M.U. Enhancing antioxidant defense system of mung bean with a salicylic acid exogenous application to mitigate cadmium toxicity. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12303. [Google Scholar]
- Rizwan, M.; Ali, S.; Abbas, T.; Zia-ur-Rehman, M.; Hannan, F.; Keller, C.; Al-Wabel, M.I.; Ok, Y.S. Cadmium minimization in wheat: A critical review. Ecotox. Environ. Saf. 2016, 130, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Wang, G.; Ji, J.; Wang, J.; Wu, H.; Ou, Y.; Li, B. Transcriptional, physiological and cytological analysis validated the roles of some key genes linked Cd stress in Salix matsudana Koidz. Environ. Exper. Bot. 2017, 134, 116–129. [Google Scholar] [CrossRef]
- Hassan, M.U.; Aamer, M.; Chattha, M.U.; Haiying, T.; Shahzad, B.; Barbanti, L.; Nawaz, M.; Rasheed, A.; Afzal, A.; Liu, Y.; et al. The critical role of zinc in plants facing the drought stress. Agriculture 2020, 10, 396. [Google Scholar] [CrossRef]
- Hassan, M.U.; Chattha, M.U.; Khan, I.; Chattha, M.B.; Barbanti, L.; Aamer, M.; Iqbal, M.M.; Nawaz, M.; Mahmood, A.; Ali, A.; et al. Heat stress in cultivated plants: Nature, impact, mechanisms, and mitigation strategies—A review. Plant Biosyst. 2021, 15, 211–234. [Google Scholar] [CrossRef]
- Dustgeer, Z.; Seleiman, M.F.; Imran, K.; Chattha, M.U.; Alhammad, B.A.; Jalal, R.S.; Refay, Y.; Hassan, M.U. Glycine-betaine induced salinity tolerance in maize by regulating the physiological attributes, antioxidant defense system and ionic homeostasis. Not. Bot. Horti Agrobot. 2021, 49, 12248. [Google Scholar] [CrossRef]
- Batool, M.; El-Badri, A.M.; Hassan, M.U.; Haiyun, Y.; Chunyun, W.; Zhenkun, Y.; Jie, K.; Wang, B.; Zhou, G. Drought stress in Brassica napus: Effects, tolerance mechanisms, and management strategies. J. Plant Growth Reg. 2022, 1–25. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Aslam, M.T.; Alhammad, B.A.; Hassan, M.U.; Maqbool, R.; Chattha, M.U.; Khan, I.; Gitari, H.I.; Uslu, O.S.; Rana, R.; et al. Salinity stress in wheat: Effects, mechanisms and management strategies. Phyton 2022, 91, 667. [Google Scholar]
- Sultan, I.; Khan, I.; Chattha, M.U.; Hassan, M.U.; Barbanti, L.; Calone, R.; Ali, M.; Majid, S.; Ghani, M.A.; Batool, M.; et al. Improved salinity tolerance in early growth stage of maize through salicylic acid foliar application. Ital. J. Agron. 2021, 16, 1810. [Google Scholar] [CrossRef]
- Li, S.; Yu, J.; Zhu, M.; Zhao, F.; Luan, S. Cadmium impairs ion homeostasis by altering K+ and Ca2+ channel activities in rice root hair cells. Plant Cell Environ. 2012, 35, 1998–2013. [Google Scholar] [CrossRef]
- Xiong, T.; Dumat, C.; Pierart, A.; Shahid, M.; Kang, Y.; Li, N.; Bertoni, G.; Laplanche, C. Measurement of metal bioaccessibility in vegetables to improve human exposure assessments: Field study of soil–plant–atmosphere transfers in urban areas, South China. Environ. Geochem. Health 2016, 38, 1283–1301. [Google Scholar] [CrossRef]
- Qu, J.H.; Meng, X.L.; Jiang, X.Y.; You, H.; Wang, P.; Ye, X.Q. Enhanced removal of Cd (II) from water using sulfur-functionalized rice husk: Characterization, adsorptive performance and mechanism exploration. J. Clean. Prod. 2018, 183, 880–886. [Google Scholar] [CrossRef]
- Subašić, M.; Šamec, D.; Selović, A.; Karalija, E. Phytoremediation of Cadmium Polluted Soils: Current Status and Approaches for Enhancing. Soil Syst. 2022, 6, 3. [Google Scholar] [CrossRef]
- Wang, K.; Li, F.; Gao, M.; Huang, Y.; Song, Z. Mechanisms of trehalose-mediated mitigation of Cd toxicity in rice seedlings. J. Clean. Prod. 2020, 267, 121982. [Google Scholar] [CrossRef]
- Gharaei, A.; Hoseini, S.S.A.; Karimi, M.; Pourjavad, E.; Amjadian, A. An integrated stochastic EPQ model under quality and green policies: Generalized cross decomposition under the separability approach. Int. J. Syst. Sci. Oper. Logist. 2019, 8, 119–131. [Google Scholar] [CrossRef]
- Yang, Y.; Ge, Y.; Tu, P.; Zeng, H.; Zhou, X.; Zou, D.; Wang, K.; Zeng, Q. Phytoextraction of Cd from a contaminated soil by tobacco and safe use of its metal-enriched biomass. J. Hazard Mater. 2019, 363, 385–393. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Ashraf, M.; Siddique, K.H. Role of glycine betaine in the thermotolerance of plants. Agronomy 2022, 12, 276. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Chen, J.; Finnegan, P.M.; Nafees, M.; Younis, A.; Shaukat, N.; Latif, N.; Abideen, Z.; Zaid, A.; Raza, A.; et al. Foliar application of trehalose or 5-aminolevulinic acid improves photosynthesis and biomass production in drought stressed Alpinia zerumbet. Agriculture 2021, 11, 908. [Google Scholar] [CrossRef]
- Shoa, J.; Wu, W.; Fahd, R.; Hassan, M.; Kai, H.; Masood, I.W.; Tasahil, S.A.; Muhammad, A.; Hu, Q.; Huang, G.; et al. Trehalose induced drought tolerance in plants: Physiological and molecular responses. Not. Bot. Horti Agrobot. 2022, 50, 12584. [Google Scholar]
- Duman, F.; Aksoy, A.; Aydin, Z.; Temizgul, R. Effects of exogenous glycinebetaine and trehalose on cadmium accumulation and biological responses of an aquatic plant (Lemna gibba L). Water Air Soil Pollut. 2010, 217, 545–556. [Google Scholar] [CrossRef]
- Ali, Q.; Ashraf, M. Induction of drought tolerance in maize (Zea mays L.) due to exogenous application of trehalose: Growth, photosynthesis, water relations and oxidative defence mechanism. J. Agron. Crop Sci. 2011, 197, 258–271. [Google Scholar] [CrossRef]
- Ponnu, J.; Wahl, V.; Schmid, M. Trehalose-6-phosphate: Connecting plant metabolism and development. Front. Plant Sci. 2011, 2, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blazquez, M.A.; Santos, E.; Floras, C.L.; Martinez-Zapater, J.M.; Salinas, J.; Gancedo, C. Isolation and molecular characterization of the Arabidopsis TPS1 gene, encoding trehalose-6-phosphate synthase. Plant J. 1998, 13, 685–689. [Google Scholar] [CrossRef] [PubMed]
- Zentella, R.; Mascorro-Gallardo, J.O.; Van Dijck, P.; Folch-Mallol, J.; Bonini, B.; Van-Vaeck, C.; Gaxiola, R.; Covarrubias, A.A.; Nieto-Sotelo, J.; Thevelein, J.M.; et al. A Selaginella lepidophylla trehalose-6-phosphate synthase complements growth and stress-tolerance defects in a yeasttps1 mutant. Plant Physiol. 1999, 119, 1473–1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, G.; Aeschbacher, R.A.; Müller, J.; Boller, T.; Wiemken, A. Trehalose-6-phosphate phosphatases from Arabidopsis thaliana: Identification by functional complementation of the yeast tps2 mutant. Plant J. 1998, 13, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, O.; Béthencourt, L.; Quero, A.; Sangwan, R.S.; Clément, C. Trehalose and plant stress responses: Friend or foe? Trends Plant Sci. 2010, 15, 409–417. [Google Scholar] [CrossRef]
- Sadak, M.S. Physiological role of trehalose on enhancing salinity tolerance of wheat plant. Bull. Nat. Res. Cent. 2019, 43, 53. [Google Scholar] [CrossRef] [Green Version]
- Bae, H.; Herman, E.; Bailey, B.; Bae, H.J.; Sicher, R. Exogenous trehalose alters Arabidopsis transcripts involved in cell wall modification, abiotic stress, nitrogen metabolism, and plant defense. Physiol. Plant 2005, 125, 114–126. [Google Scholar] [CrossRef]
- Alemu, I.D. General characteristics and genetic improvement status of mung bean (Vigna radiata L.) in Ethiopia. Intern. J. Agric. Inno. Res. 2016, 5, 232–237. [Google Scholar]
- Muhammad, U.C.; Muhammad, U.H.; Imran, K.; Muhammad, B.C.; Imran, A.; Muhammad, F.; Mina, K. Effect of different nitrogen and phosphorus fertilizer levels in combination with nitrogen and phosphorus solubilizing inoculants on the growth and yield of mung bean. Pak. J. Life Soc. Sci. 2017, 15, 31–36. [Google Scholar]
- Arnon, D.I. Copper enzymes in isolated chloroplasts, polyphenoxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Mostofa, M.G.; Fujita, M. Salicylic acid alleviates copper toxicity in rice (Oryza sativa L.) seedlings by up-regulating antioxidative and glyoxalase systems. Ecotoxicology 2013, 22, 959–973. [Google Scholar] [CrossRef]
- Cakmak, I.; Horst, W.J. Effect of aluminium on lipid peroxidation, superoxide dismutase, catalase, and peroxidase activities in root tips of soybean (Glycine max). Physiol. Plant. 1991, 83, 463–468. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants. Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 2, 248–254. [Google Scholar] [CrossRef]
- Vanslyke, D.D.; Macfadyen, D.A.; Hamilton, P.B. The gasometric determination of amino acids in mine by the ninhydrin-carbon dioxide method. J. Biol. Chem. 1943, 150, 251–258. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. Assay of catalases and peroxidases. Methods Enzymol. 1955, 2, 764–775. [Google Scholar]
- Guan, J.S.; Haggarty, S.J.; Giacometti, E.; Dannenberg, J.H.; Joseph, N.; Gao, J.; Nieland, T.J.; Zhou, Y.; Wang, X.; Mazitschek, R.; et al. HDAC2 negatively regulates memory formation and synaptic plasticity. Nature 2009, 459, 55–60. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Purification of ascorbate peroxidase in spinach chloroplasts; Its inactivation in ascorbate-depleted medium and reactivation by mono-dehydroascorbate radical. Plant Cell Physiol. 1987, 28, 131–140. [Google Scholar]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of water stree-induced changes in the leaves of indigenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant. 1983, 58, 166170. [Google Scholar] [CrossRef]
- Jones, J.B.; Case, V.W. Sampling, handling, and analyzing plant tissue samples. Soil Test. Plant Anal. 1990, 3, 389–427. [Google Scholar]
- Steel, R.G.D.; Torrie, J.H.; Dicky, D.A. Principles and Procedures of Statistics: A Biometrical Approach, 3rd ed.; McGraw Hill, Inc. Book Co.: New York, NY, USA, 1997; pp. 352–358. [Google Scholar]
- Cao, F.; Wang, R.; Cheng, W.; Zeng, F.; Ahmed, I.M.; Hu, X.; Zhang, G.; Wu, F. Genotypic and environmental variation in cadmium, chromium, lead and copper in rice and approaches for reducing the accumulation. Sci. Total Environ. 2014, 496, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Saidi, I.; Ayouni, M.; Dhieb, A.; Chtourou, Y.; Chaïbi, D.W. Oxidative damages induced by short-term exposure to cadmium in bean plants: Protective role of salicylic acid. S. Afr. J. Bot. 2013, 85, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.Z.; Yang, W.Q.; Zhang, J.; Zhou, L.Q. Effects of cadmium stress on growth and nutrient accumulation, distribution and utilization in Osmanthus fragrans var. thunbergii. Chin. J. Plant Ecol. 2010, 34, 1220. [Google Scholar]
- Cengiz, K.; Nudrat, A.; Akram, M.; Ashraf, M.; Nasser, A.; Parvaiz, A. Exogenously supplied silicon (Si) improves cadmium tolerance in pepper (Capsicum annuum L.) by upregulating the synthesis of nitric oxide and hydrogen sulfide. J. Biotechnol. 2020, 3, 316. [Google Scholar]
- Wang, H.; Zhao, S.C.; Xia, W.J. Effects of cadmium stress at different concentrations on photosynthesis, lipid peroxidation and antioxidant enzyme activities in maize seedlings. J. Plant Nutr. Fert. 2008, 14, 36–42. [Google Scholar]
- Dong, L.B.; Yu, D.; Lin, X.T.; Wang, B.; Pan, L. Improving expression of thermostable trehalase from Myceliophthora sepedonium in Aspergillus Niger mediated by the CRISPR/Cas9 tool and its purification, characterization. Protein Expr. Purif. 2020, 165, 105482. [Google Scholar] [CrossRef]
- Zulfiqar, U.; Ayub, A.; Hussain, S.; Waraich, E.A.; El-Esawi, M.A.; Ishfaq, M.; Ahmad, M.; Ali, N.; Maqsood, M.F. Cadmium Toxicity in Plants: Recent Progress on Morpho-physiological Effects and Remediation Strategies. J. Soil Sci. Plant Nutr. 2022, 22, 212–269. [Google Scholar] [CrossRef]
- Chen, X.; Pu, G.; Huang, Y.; Mo, L. Effects of thallium and cadmium stress on growth and photosynthetic characteristics of Arundo donax. Guangxi Zhiwu/Guihaia 2019, 39, 743–751. [Google Scholar]
- Kosar, F.; Akram, N.A.; Ashraf, M.; Sadiq, M.; Al-Qurainy, F. Trehalose-induced improvement in growth, photosynthetic characteristics and levels of some key osmoprotectants in sunflower (Helianthus annuus L.) under drought stress. Pak. J. Bot. 2018, 50, 955–961. [Google Scholar]
- Qin, J.; Dong, W.Y.; He, K.N.; Yu, Y.; Tan, G.D.; Han, L.; Dong, M.; Zhang, Y.Y.; Zhang, D.; Li, A.Z.; et al. NaCl salinity-induced changes in water status, ion contents and photosynthetic properties of Shepherdia argentea (Pursh) Nutt. seedlings. Plant Soil Environ. 2010, 56, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Zeid, I.M. Effect of arginine and urea on polyamines content and growth of bean under salinity stress. Acta Physiol. Plant 2009, 31, 65–70. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Chen, J.; Finnegan, P.M.; Younis, A.; Nafees, M.; Zorrig, W.; Hamed, K.B. Application of trehalose and salicylic acid mitigates drought stress in sweet basil and improves plant growth. Plants 2021, 10, 1078. [Google Scholar] [CrossRef] [PubMed]
- Chaâbene, Z.; Rorat, A.; Hakim, I.R.; Bernard, F.; Douglas, G.C.; Elleuch, A.; Vandenbulcke, F.; Mejdoub, H. Insight into the expression variation of metal-responsive genes in the seedling of date palm (Phoenix dactylifera). Chemosphere 2018, 197, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palma, J.M.; Sandalio, L.M.; Corpas, F.J.; Romero-Puertas, M.C.; McCarthy, I.; Luis, A. Plant proteases, protein degradation, and oxidative stress: Role of peroxisomes. Plant Physiol. Biochem. 2002, 40, 521–530. [Google Scholar] [CrossRef]
- Zemanová, V.; Pavlík, M.; Pavlíková, D.; Tlustoš, P. The significance of methionine, histidine and tryptophan in plant responses and adaptation to cadmium stress. Plant Soil Environ. 2014, 60, 426–432. [Google Scholar] [CrossRef]
- Zoghlami, L.B.; Djebali, W.; Abbes, Z.; Hediji, H.; Maucourt, M.; Moing, A.; Brouquisse, R.; Chaibi, W. Metabolite modifications in Solanum lycopersicum roots and leaves under cadmium stress. Afr. J. Biotechnol. 2011, 10, 567–579. [Google Scholar]
- Metwally, A.; Safronova, V.I.; Belimov, A.A.; Dietz, K.J. Genotypic variation of the response to cadmium toxicity in Pisum sativum L. J. Exp. Bot. 2005, 56, 167–178. [Google Scholar] [CrossRef]
- Akhter, M.; Macfie, S. Species-specific relationship between transpiration and cadmium translocation in lettuce, barley and radish. J. Plant Stud. 2012, 1, 2–13. [Google Scholar]
- Joshi, R.; Sahoo, K.K.; Singh, A.K.; Anwar, K.; Pundir, P.; Gautam, R.K.; Krishnamurthy, S.L.; Sopory, S.K.; Pareek, A.; Singla-Pareek, S.L. Enhancing trehalose biosynthesis improves yield potential in marker-free transgenic rice under drought, saline, and sodic conditions. J. Exp. Bot. 2020, 71, 653–668. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Root Length (cm) | Shoot Length (cm) | Root Fresh Weight (g) | Shoot Fresh Weight (g) | Root Dry Weight (g) | Shoot Dry Weight (g) | Leaves Per Plant | |
|---|---|---|---|---|---|---|---|---|
| Cd0 | Tre0 | 4.89 ± 0.029 cd | 25.40 ± 0.036 c | 0.76 ± 0.004 c | 5.23 ± 0.111 de | 0.27 ± 0.003 c | 0.71 ± 0.007 c | 11.0 ± 0.559 ab |
| Cd0 | Tre1 | 5.90 ± 0.158 b | 27.13 ± 0.059 ab | 1.16 ± 0.044 a | 6.96 ± 0.076 b | 0.34 ± 0.007 b | 0.83 ± 0.005 b | 12.0 ± 0.211 ab |
| Cd0 | Tre2 | 6.67 ± 0.075 a | 28.10 ± 0.073 a | 1.13 ± 0.021 a | 7.67 ± 0.055 a | 0.41 ± 0.004 a | 1.03 ± 0.020 a | 13.0 ± 0.366 a |
| Cd1 | Tre0 | 3.43 ± 0.045 f | 22.29 ± 0.124 e | 0.63 ± 0.004 d | 4.73 ± 0.021 f | 0.26 ± 0.004 c | 0.69 ± 0.005 c | 11.0 ± 0.422 bc |
| Cd1 | Tre1 | 4.13 ± 0.049 e | 23.72 ± 0.039 d | 1.00 ± 0.013 b | 4.93 ± 0.055 ef | 0.26 ± 0.004 c | 0.73 ± 0.007 c | 11.0 ± 0.559 bc |
| Cd1 | Tre2 | 5.33 ± 0.042 c | 26.57 ± 0.055 bc | 0.84 ± 0.004 c | 6.17 ± 0.042 c | 0.32 ± 0.007 b | 0.88 ± 0.009 b | 10.0 ± 0.366 cd |
| Cd2 | Tre0 | 3.17 ± 0.091 f | 19.20 ± 0.036 f | 0.49 ± 0.004 e | 3.83 ± 0.092 g | 0.15 ± 0.004 e | 0.57 ± 0.002 e | 9.0 ± 0.634 d |
| Cd2 | Tre1 | 4.67 ± 0.046 d | 22.97 ± 0.046 de | 0.60 ± 0.005 d | 4.83 ± 0.057 ef | 0.21 ± 0.004 d | 0.64 ± 0.005 d | 9.0 ± 0.332 d |
| Cd2 | Tre2 | 4.97 ± 0.062 cd | 24.13 ± 0.107 d | 0.77 ± 0.006 c | 5.56 ± 0.112 d | 0.25 ± 0.004 c | 0.70 ± 0.004 c | 10.0 ± 0.211 cd |
| Treatments | Root Length (cm) | Shoot Length (cm) | Root Fresh Weight (g) | Shoot Fresh Weight (g) | Root Dry Weight (g) | Shoot Dry Weight (g) | |
|---|---|---|---|---|---|---|---|
| Cd0 | Tre0 | 3.55 ± 0.048 e | 20.56 ± 0.077 c | 0.61 ± 0.005 c | 4.71 ± 0.087 d | 0.21 ± 0.004 d | 0.53 ± 0.006 d |
| Cd0 | Tre1 | 4.88 ± 0.097 b | 21.68 ± 0.062 b | 0.80 ± 0.053 b | 5.11 ± 0.082 c | 0.28 ± 0.008 b | 0.63 ± 0.007 c |
| Cd0 | Tre2 | 5.79 ± 0.112 a | 22.52 ± 0.054 a | 0.96 ± 0.016 a | 6.15 ± 0.075 a | 0.35 ± 0.006 a | 0.91 ± 0.011 a |
| Cd1 | Tre0 | 3.06 ± 0.083 f | 17.11 ± 0.086 f | 0.44 ± 0.006 d | 3.61 ± 0.048 f | 0.17 ± 0.007 e | 0.45 ± 0.008 e |
| Cd1 | Tre1 | 3.95 ± 0.091 d | 18.42 ± 0.104 e | 0.62 ± 0.023 c | 4.62 ± 0.062 d | 0.24 ± 0.005 c | 0.53 ± 0.006 d |
| Cd1 | Tre2 | 4.42 ± 0.065 c | 19.51 ± 0.113 d | 0.79 ± 0.008 b | 5.58 ± 0.051 b | 0.29 ± 0.006 b | 0.69 ± 0.008 b |
| Cd2 | Tre0 | 2.81 ± 0.088 g | 14.01 ± 0.098 h | 0.31 ± 0.005 e | 3.01 ± 0.063 g | 0.12 ± 0.006 f | 0.43 ± 0.004 e |
| Cd2 | Tre1 | 3.56 ± 0.057 e | 15.89 ± 0.087 g | 0.46 ± 0.008 d | 4.19 ± 0.071 e | 0.17 ± 0.005 e | 0.61 ± 0.006 c |
| Cd2 | Tre2 | 4.34 ± 0.091 c | 17.08 ± 0.111 f | 0.60 ± 0.009 c | 5.07 ± 0.092 c | 0.21 ± 0.006 d | 0.63 ± 0.005 c |
| Treatments | Pod Length (cm) | Grains/Pod | 100 Seed Weight (g) | Grain Yield (g pot−1) | |
|---|---|---|---|---|---|
| Cd0 | Tre0 | 11.13 ± 0.14 bc | 11.33 ± 0.27 abc | 6.24 ± 0.034 b | 37.67 ± 1.29 bc |
| Cd0 | Tre1 | 11.46 ± 0.21 b | 12.33 ± 0.29 ab | 6.33 ± 0.053 b | 39.67 ± 1.49 ab |
| Cd0 | Tre2 | 12.20 ± 0.16 a | 12.67 ± 0.54 a | 6.65 ± 0.062 a | 44.00 ± 1.41 a |
| Cd1 | Tre0 | 10.20 ± 0.17 e | 9.67 d ± 0.53 de | 5.03 ± 0.030 e | 33.00 ± 1.25 d |
| Cd1 | Tre1 | 10.62 ± 0.12 de | 10.67 ± 0.27 bcd | 5.47 ± 0.110 d | 32.67 ± 1.18 d |
| Cd1 | Tre2 | 10.87 ± 0.11 cd | 11.00 ± 0.47 bcd | 5.92 ± 0.072 c | 35.00 ± 0.94 cd |
| Cd2 | Tre0 | 8.02 ± 0.03 g | 8.33 ± 0.29 f | 4.13 ± 0.064 g | 25.67 ± 0.98 f |
| Cd2 | Tre1 | 8.60 ± 0.14 f | 8.33 ± 0.30 f | 4.44 ± 0.047 f | 26.67 ± 0.72 e |
| Cd2 | Tre2 | 9.07 ± 0.07 f | 9.00 ± 0.48 e | 4.61 ± 0.072 f | 28.00 ± 1.69 e |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rehman, S.; Chattha, M.U.; Khan, I.; Mahmood, A.; Hassan, M.U.; Al-Huqail, A.A.; Salem, M.Z.M.; Ali, H.M.; Hano, C.; El-Esawi, M.A. Exogenously Applied Trehalose Augments Cadmium Stress Tolerance and Yield of Mung Bean (Vigna radiata L.) Grown in Soil and Hydroponic Systems through Reducing Cd Uptake and Enhancing Photosynthetic Efficiency and Antioxidant Defense Systems. Plants 2022, 11, 822. https://doi.org/10.3390/plants11060822
Rehman S, Chattha MU, Khan I, Mahmood A, Hassan MU, Al-Huqail AA, Salem MZM, Ali HM, Hano C, El-Esawi MA. Exogenously Applied Trehalose Augments Cadmium Stress Tolerance and Yield of Mung Bean (Vigna radiata L.) Grown in Soil and Hydroponic Systems through Reducing Cd Uptake and Enhancing Photosynthetic Efficiency and Antioxidant Defense Systems. Plants. 2022; 11(6):822. https://doi.org/10.3390/plants11060822
Chicago/Turabian StyleRehman, Sadia, Muhammad Umer Chattha, Imran Khan, Athar Mahmood, Muhammad Umair Hassan, Asma A. Al-Huqail, Mohamed Z. M. Salem, Hayssam M. Ali, Christophe Hano, and Mohamed A. El-Esawi. 2022. "Exogenously Applied Trehalose Augments Cadmium Stress Tolerance and Yield of Mung Bean (Vigna radiata L.) Grown in Soil and Hydroponic Systems through Reducing Cd Uptake and Enhancing Photosynthetic Efficiency and Antioxidant Defense Systems" Plants 11, no. 6: 822. https://doi.org/10.3390/plants11060822
APA StyleRehman, S., Chattha, M. U., Khan, I., Mahmood, A., Hassan, M. U., Al-Huqail, A. A., Salem, M. Z. M., Ali, H. M., Hano, C., & El-Esawi, M. A. (2022). Exogenously Applied Trehalose Augments Cadmium Stress Tolerance and Yield of Mung Bean (Vigna radiata L.) Grown in Soil and Hydroponic Systems through Reducing Cd Uptake and Enhancing Photosynthetic Efficiency and Antioxidant Defense Systems. Plants, 11(6), 822. https://doi.org/10.3390/plants11060822