
Biochemical and Physiological Plant Processes Affected by Seed Treatment with Non-Thermal Plasma



Abstract
:1. Introduction
2. Definitions of Plasma, Low-Temperature Plasma and Description of Different Types of Devices Used for Seed, Plant or Water Treatment
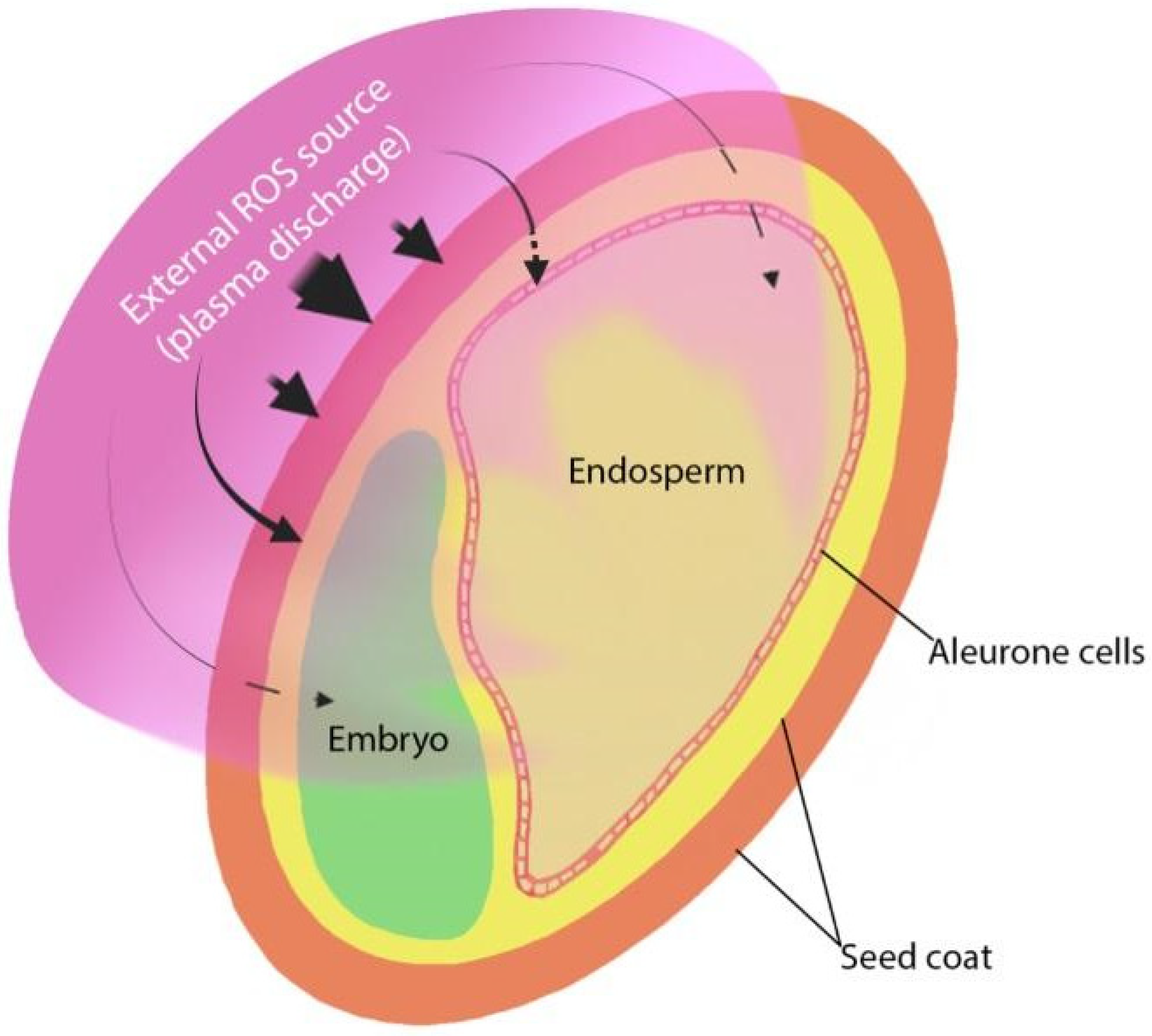
3. NTP Effects on Seed Germination and Early Seedling Growth

{kind=link}
{kind=link}
| Effect | NTP Device | Plant Species (NTP Feeding Gas if Not Air) [References] |
|---|---|---|
| Positive | Low-pressure CP | Ajwain [44], bean [45,46], black mulberry [47]; industrial hemp [48], lamb’s quarters [41,42], lentil [45], maize [46], mung bean [50], wheat [37,45,49], oilseed rape [52], quinoa [53], red clover [54,55], rice [56], soybean [57], sunflower [58], tomato [40,59]; artichoke (nitrogen) [60], common bean (oxygen) [61], safflower (argon) [62], wheat (helium) [63], wheat (argon) [64]. |
| DBD plasma | Barley [65], black pine [66], cotton [67], cucumber, [68], green chiretta [69], Norway spruce [70], quinoa [53]; pea [71,72], pepper [68], radish [73,74,75,76,77,78]; rice [79,80], sunflower [81,82], sweet basil [83], thale cress [84,85,86,87], wheat [51,88,89], zinnia [84]; barley (nitrogen + 0.65% air) [90], carrot (argon) [91], coriander (nitrogen) [92], cotton (argon) [67], rice (argon + air) [93], soybean (oxygen, nitrogen) [94,95], soybean (argon) [96], sweet basil (argon + oxygen) [97], wheat (argon/air, argon/oxygen) [98,99]. | |
| Plasma jet | Common bean (helium) [100], Erythrina velutina (helium) [101], fenugreek (argon) [102] mung bean (air, oxygen) [103], wheat (nitrogen) [104]. | |
| Gliding arc | Buckwheat (air) [105], garden tree-mallow (nitrogen) [106], industrial hemp (air) [107], maize (air) [108]. | |
| Neutral | Low-pressure CP | Blue lupin [49], buckwheat [109], wheat (oxygen) [110]. |
| Gliding arc | Buckwheat [105], industrial hemp [107]. | |
| DBD plasma | Coriander [92], maize [111], Norway spruce [70], radish [76], Scotish pine [112]. | |
| Plasma jet | Mung bean (helium, nitrogen) [64] | |
| Negative | Low-pressure CP | Barley, radish, pea, soybean, corn, bean (fluorocarbon, nitrogen, carbon-containing compounds) [35], Norway spruce [113], rhododendron [47], buckwheat (oxygen) [114], wheat (oxygen) [110]. |
| DBD plasma | Buckwheat [105]; Scots pine [112], pea (nitrogen, oxygen) [72], sunflower [81]. |
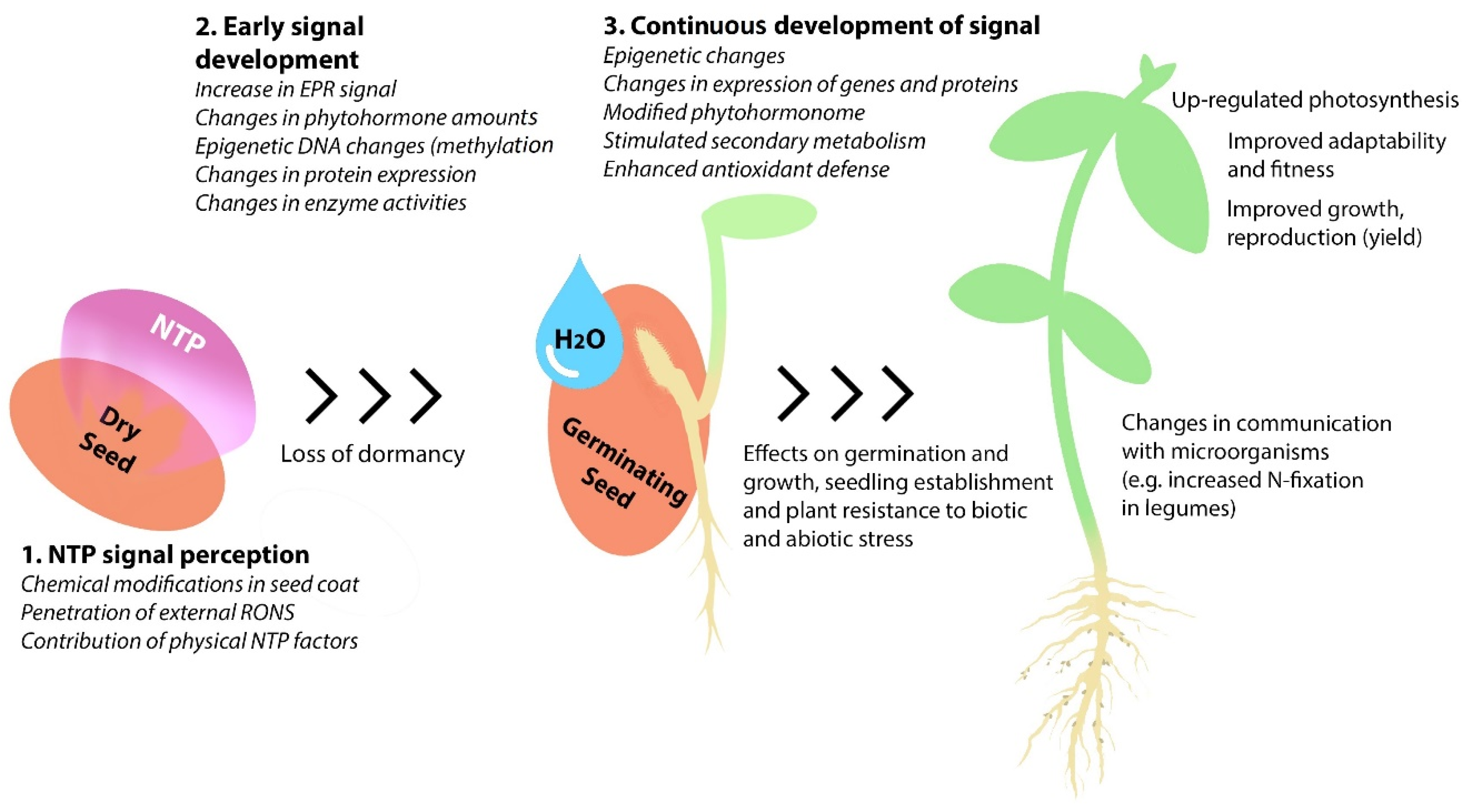
4. The Mechanisms of NTP Effects on Seed Germination
4.1. The Role of NTP-Induced Physical and Chemical Changes in the Seed Coat
4.2. The Impact of Seed Dormancy and Phytohormones
4.3. Involvement of the Internal Generation of Reactive Oxygen Species in the Effects of NTP on Germination and Plant Growth
4.4. Effects of Plasma Treatment on Enzymatic Activities in Dry and Germinating Seeds
5. Effects of Plasma Treatment on Biochemical and Physiological Processes in Growing Seedlings and Plants
5.1. Impact on Plant Epigenetics and Protein Expression
5.2. Changes in Enzyme Activities
5.3. Effects on the Amount of Phytohormones in Plant Tissues
5.4. Effect on Photosynthesis
5.5. Changes in Amount of Secondary Metabolites
5.6. NTP Effects on Plant Adaptability and Stress Resistance
5.7. Effects on Plant Growth and Productivity
6. NTP-Induced Changes in the Seed Microbiome and Plant-Microbial Interactions
7. Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rockström, J.; Williams, J.; Daily, G.; Noble, A.; Matthews, N.; Gordon, L.; Wetterstrand, H.; DeClerck, F.; Shah, M.; Steduto, P.; et al. Sustainable intensification of agriculture for human prosperity and global sustainability. Ambio 2017, 46, 4–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewel, J.J.; Schreeg, L.A.; Sinclair, T.R. Resources for crop production: Accessing the unavailable. Trends Plant Sci. 2019, 24, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Waskow, A.; Howling, A.; Furno, I. Mechanisms of plasma-seed treatments as a potential seed processing technology. Front. Phys. 2021, 9, 617345. [Google Scholar] [CrossRef]
- Araújo, S.S.; Paparella, S.; Dondi, D.; Bentivoglio, A.; Carbonera, D.; Balestrazzi, A. Physical methods for seed invigoration: Advantages and challenges in seed technology. Front. Plant Sci. 2016, 7, 646. [Google Scholar] [CrossRef] [Green Version]
- Adamovich, I.; Baalrud, S.D.; Bogaerts, A.; Bruggeman, P.J.; Cappelli, M.; Colombo, V.; Czarnecki, U.; Ebert, U.; Eden, J.G.; Favia, P.; et al. The 2017 Plasma Roadmap: Low temperature plasma science and technology. J. Phys. D Appl. Phys. 2017, 50, 323001. [Google Scholar] [CrossRef]
- Weltmann, K.; Kolb, J.F.; Holub, M.; Uhrlandt, D.; Šimek, M.; Ostrikov, K.; Hamaguchi, S.; Cvelbar, U.; Černák, M.; Locke, B.; et al. The future for plasma science and technology. Plasma Proc. Polym. 2018, 16, e1800118. [Google Scholar] [CrossRef] [Green Version]
- Straňák, V.; Tichý, M.; Kříha, V.; Scholtz, V.; Šerá, B.; Špatenka, P. Technological applications of sulfatron produced discharge. J. Optoelectron. Adv. Mater. 2007, 9, 852–857. [Google Scholar]
- Laroussi, M.; Lu, X.; Keidar, M. Perspective: The physics, diagnostics, and applications of atmospheric pressure low temperature plasma sources used in plasma medicine. J. Appl. Phys. 2017, 122, 020901. [Google Scholar] [CrossRef]
- Thirumdas, R.; Kothakota, A.; Annapure, U.; Siliveru, K.; Blundell, R.; Gatt, R.; Valdramidis, V.P. Plasma activated water (PAW): Chemistry, physico-chemical properties, applications in food and agriculture. Trends Food Sci. Technol. 2018, 77, 21–31. [Google Scholar] [CrossRef]
- Liao, X.Y.; Cullen, P.J.; Muhammad, A.I.; Jiang, Z.M.; Ye, X.Q.; Liu, D.H.; Ding, T. Cold plasma-based hurdle interventions: New strategies for improving food safety. Food Eng. Rev. 2020, 12, 321–332. [Google Scholar] [CrossRef]
- Liu, D.W.; Zhang, Y.H.; Xu, M.Y.; Chen, H.X.; Lu, X.P.; Ostrikov, K. Cold atmospheric pressure plasmas in dermatology: Sources, reactive agents, and therapeutic effects. Plasma Proc. Polym. 2020, 17, e1900218. [Google Scholar] [CrossRef]
- Zhu, Y.L.; Li, C.Z.; Cui, H.Y.; Lin, L. Feasibility of cold plasma for the control of biofilms in food industry. Trends Food Sci. Technol. 2020, 99, 142–151. [Google Scholar] [CrossRef]
- Šera, B.; Šery, M. Non-thermal plasma treatment as a new biotechnology in relation to seeds, dry fruits, and grains. Plasma Sci. Technol. 2018, 20, 044012. [Google Scholar] [CrossRef] [Green Version]
- Attri, P.; Ishikawa, K.; Okumura, T.; Koga, K.; Shiratani, M. Plasma agriculture from laboratory to farm: A review. Processes 2020, 8, 1002. [Google Scholar] [CrossRef]
- Staric, P.; Vogel-Mikuš, K.; Mozetic, M.; Junkar, I. Effects of nonthermal plasma on morphology, genetics and physiology of seeds: A review. Plants 2020, 9, 1736. [Google Scholar] [CrossRef]
- Adhikari, B.; Adhikari, M.; Park, G. The effects of plasma on plant growth, development, and sustainability. Appl. Sci. 2020, 10, 6045. [Google Scholar] [CrossRef]
- Song, J.-S.; Kim, S.B.; Ryu, S.; Oh, J.; Kim, D.-S. Emerging plasma technology that alleviates crop stress during the early growth stages of plants: A Review. Front. Plant Sci. 2020, 11, 988. [Google Scholar] [CrossRef]
- Holubová, L.; Kyzek, S.; Durovcová, I.; Fabová, J.; Horváthová, E.; Ševcovicová, A.; Gálová, E. Non-thermal plasma—a new green priming agent for plants? Int. J. Mol. Sci. 2020, 21, 9466. [Google Scholar] [CrossRef]
- Waskow, A.; Avino, F.; Howling, A.; Furno, I. Entering the plasma agriculture field: An attempt to standardize protocols for plasma treatment of seeds. Plasma Process Polym. 2021, 18, e2100152. [Google Scholar] [CrossRef]
- Ranieri, P.; Sponsel, N.; Kizer, J.; Rojas-Pierce, M.; Hernández, R.; Gatiboni, L.; Grunden, A.; Stapelmann, K. Plasma agriculture: Review from the perspective of the plant and its ecosystem. Plasma Proc. Polym. 2020, 18, e2000162. [Google Scholar] [CrossRef]
- Braithwaite, N.S.J. Introduction to gas discharges. Plasma Sources Sci. Technol. 2000, 9, 517–527. [Google Scholar]
- Cheruthazhekatt, S.; Cernak, M.; Slavicek, P.; Havel, J. Gas plasmas and plasma modified materials in medicine. J. Appl. Biomed. 2010, 8, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Nehra, V.; Kumar, A.; Dwivedi, H. Atmospheric non-thermal plasma sources. Int. J. Eng. 2008, 2, 53–68. [Google Scholar]
- Piel, A. Plasma Physics. An Introduction to Laboratory, Space, and Fusion Plasmas, 2nd ed.; Springer International Publishing AG: Cham, Switzerland, 2017. [Google Scholar]
- Fridman, G. Non-equilibrium plasmas in biology and medicine. In Biological and Environmental Application of Gas Discharge Plasmas; Brelles, M.G., Ed.; California State Polytechnic University: Pomona, CA, USA, 2012; pp. 95–184. [Google Scholar]
- Misra, N.N.; Schlutter, O.; Cullen, P. Plasma in Food and agriculture. In Cold Plasma in Food and Agriculture: Fundamentals and Applications; Misra, N.N., Schlutter, O., Cullen, P.J.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 1–16. [Google Scholar]
- Conrads, H.; Schmidt, M. Plasma generation and plasma sources. Plasma Sources Sci. Technol. 2000, 9, 441–454. [Google Scholar] [CrossRef] [Green Version]
- Denes, F.S.; Manolache, S. Macromolecular plasma-chemistry: An emerging field of polymer science. Prog. Polym. Sci. 2004, 29, 815–885. [Google Scholar] [CrossRef]
- Randeniya, L.K.; de Groot, G.J.J.B. Non-Thermal Plasma Treatment of Agricultural Seeds for Stimulation of Germination, Removal of Surface Contamination and Other Benefits: A Review. Plasma Proc. Polym. 2015, 12, 608–623. [Google Scholar] [CrossRef]
- Lu, P.; Cullen, P.J.; Ostrikov, K. Atmospheric Pressure Nonthermal Plasma. In Cold Plasma in Food and Agriculture: Fundamentals and Applications; Misra, N.N., Schlutter, O., Culle, P.J.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 83–116. [Google Scholar]
- Turner, M. Physics of cold plasma. In Cold Plasma in Food and Agriculture: Fundamentals and Applications; Misra, N.N., Schlutter, O., Culle, P.J.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 17–50. [Google Scholar]
- Whitehead, J.C. The chemistry of cold plasma. In Cold Plasma in Food and Agriculture: Fundamentals and Applications; Misra, N.N., Schlutter, O., Culle, P.J.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 53–81. [Google Scholar]
- Droge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef]
- Jeevan Kumar, S.P.; Rajendra Prasad, S.; Banerjee, R.; Thammineni, C. Seed birth to death: Dual functions of reactive oxygen species in seed physiology. Ann. Bot. 2015, 116, 663–668. [Google Scholar] [CrossRef] [Green Version]
- Del Rio, L.A. ROS and RNS in plant physiology: An overview. J. Biol. Chem. 2015, 66, 2827–2837. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Zhou, R.; Wang, P.; Xian, Y.; Mai-Prochnow, A.; Lu, X.P.; Cullen, P.J.; Ostrikov, K.; Bazaka, K. Plasma-activated water: Generation, origin of reactive species and biological applications. J. Phys. D Appl. Phys. 2020, 53, 303001. [Google Scholar] [CrossRef]
- Dubinov, A.E.; Lazarenko, E.M.; Selemir, V.D. Effect of glow discharge air plasma on grain crops seed. IEEE Trans. Plasma Sci. 2000, 28, 180–183. [Google Scholar] [CrossRef]
- Volin, J.C.; Denes, F.S.; Young, R.A.; Park, S.M.T. Modification of seed germination performance through cold plasma chemistry technology. Crop Sci. 2000, 40, 1706–1718. [Google Scholar] [CrossRef]
- Živkovic, S.; Puac, N.; Giba, Z.; Grubišic, D.; Petrovic, Z.L. The stimulatory effect of non-equilibrium (low temperature) air plasma pretreatment on light-induced germination of Paulownia tomentosa seeds. Seed Sci. Technol. 2004, 32, 693–701. [Google Scholar] [CrossRef]
- Meiqiang, Y.; Mingjing, H.; Buzhou, M.; Tengcai, M. Stimulating effects of seed treatment by magnetized plasma on tomato growth and yield. Plasma Sci. Technol. 2005, 7, 3143–3147. [Google Scholar] [CrossRef]
- Šerá, B.; Stranák, V.; Šerý, M.; Tichý, M.; Spatenka, P. Germination of Chenopodium album in response to microwave plasma treatment. Plasma Sci. Technol. 2008, 10, 506–511. [Google Scholar] [CrossRef]
- Šerá, B.; Sery, M.; Stranak, V.; Spatenka, P.; Tichy, M. Does cold plasma affect breaking dormancy and seed germination? A study on seeds of Lamb’s Quarters (Chenopodium album agg.). Plasma Sci. Technol. 2009, 11, 750–754. [Google Scholar]
- Bewley, J.D.; Bradford, K.; Hilhorst, H.; Nonogaki, H. Seeds. Physiology of Development, Germination and Dormancy, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 133–181. [Google Scholar]
- Gholami, A.; Safa, N.N.; Khoram, M.; Hadian, J.; Ghomi, H. Effect of low-pressure radio frequency plasma on ajwain seed germination. Plasma Med. 2016, 6, 389–396. [Google Scholar] [CrossRef] [Green Version]
- Bormashenko, E.; Grynyov, R.; Bormashenko, Y.; Drori, E. Cold radiofrequency plasma treatment modifies wettability and germination speed of plant seeds. Sci. Rep. 2012, 2, 741–748. [Google Scholar] [CrossRef]
- Bormashenko, E.; Shapira, Y.; Grynyov, R.; Bormashenko, Y.; Drori, E. Interaction of cold radiofrequency plasma with seeds of beans (Phaseolus vulgaris). J. Exp. Bot. 2015, 66, 4013–4021. [Google Scholar] [CrossRef] [Green Version]
- Mildaziene, V.; Pauzaite, G.; Malakauskiene, A.; Zukiene, R.; Nauciene, Z.; Filatova, I.; Azharonok, V.; Lyushkevich, V. Response of perennial woody plants to seed treatment by electromagnetic field and low-temperature plasma. Bioelectromagnetics 2016, 37, 536–548. [Google Scholar] [CrossRef]
- Ivankov, A.; Nauciene, Z.; Zukiene, R.; Degutyte-Fomins, L.; Malakauskiene, A.; Kraujalis, P.; Venskutonis, P.R.; Filatova, I.; Lyushkevich, V.; Mildaziene, V. Changes in growth and production of non-psychotropic cannabinoids induced by pre-sowing treatment of hemp seeds with cold plasma, vacuum and electromagnetic field. Appl. Sci. 2020, 10, 8519. [Google Scholar]
- Filatova, I.; Azharonok, V.; Lushkevich, V.; Zhukovsky, A.; Gadzhieva, G.; Spasic, K.; Zivkovic, S.; Puac, N.; Lazović, S.; Malović, G.; et al. Plasma Seeds Treatment as a Promising Technique for Seed Germination Improvement. In Proceedings of the 31st International Conference on Phenomena in Ionized Gases, Granada, Spain, 14–19 July 2013; p. PS4-105. Available online: https://www.semanticscholar.org/paper/Plasma-seeds-treatment-as-a-promising-technique-for-Filatova-Azharonok/5e45e5f0a890d8e65a4936292d55173fdddd6c95 (accessed on 15 March 2022).
- Sadhu, S.; Thirumdas, R.; Deshmukh, R.R.; Annapure, U.S. Influence of cold plasma on the enzymatic activity in germinating mung beans (Vigna radiate). LWT-Food Sci. Technol. 2017, 78, 97–104. [Google Scholar] [CrossRef]
- Los, A.; Ziuzina, D.; Boehm, D.; Cullen, P.J.; Bourke, P. Investigation of mechanisms involved in germination enhancement of wheat (Triticum aestivum) by cold plasma: Effects on seed surface chemistry and characteristics. Plasma Proc. Polym. 2019, 16, 1800148. [Google Scholar] [CrossRef]
- Li, L.; Li, J.; Shen, M.; Zhang, C.; Dong, Y. Cold plasma treatment enhances oilseed rape seed germination under drought stress. Sci. Rep. 2015, 5, 13033. [Google Scholar] [CrossRef]
- Gómez-Ramírez, A.; López-Santos, C.; Cantos, M.; García, J.L.; Molina, R.; Cotrino, J.; Espinos, J.P.; González-Elipe, A.R. Surface chemistry and germination improvement of Quinoa seeds subjected to plasma activation. Sci. Rep. 2017, 7, 5924. [Google Scholar] [CrossRef] [Green Version]
- Mildaziene, V.; Pauzaite, G.; Nauciene, Z.; Zukiene, R.; Malakauskiene, A.; Norkeviciene, E.; Stukonis, V.; Slepetiene, A.; Olsauskaite, V.; Padarauskas, A.; et al. Effect of seed treatment with cold plasma and electromagnetic field on red clover germination, growth and content of major isoflavones. J. Phys. D Appl. Phys. 2020, 53, 26. [Google Scholar]
- Ivankov, A.; Zukiene, R.; Nauciene, Z.; Degutyte-Fomins, L.; Filatova, I.; Lyushkevich, V.; Mildaziene, V. The effects of red clover seed treatment with cold plasma and electromagnetic field on germination and seedling growth are dependent on seed color. Appl. Sci. 2021, 11, 4676. [Google Scholar] [CrossRef]
- Chen, H.H.; Chang, H.C.; Chen, Y.K.; Hung, C.L.; Lin, S.Y.; Chen, Y.S. An improved process for high nutrition of germinated brown rice production: Low-pressure plasma. Food Chem. 2016, 191, 120–127. [Google Scholar] [CrossRef]
- Li, L.; Jiang, J.; Li, J.; Shen, M.; Xe, X.; Shao, H.; Yuanhua, D. Effects of cold plasma treatment on seed germination and seedling growth of soybean. Sci. Rep. 2014, 4, 5859–5865. [Google Scholar] [CrossRef] [Green Version]
- Mildažienė, V.; Aleknavičiūtė, V.; Žūkienė, R.; Paužaitė, G.; Naučienė, Z.; Filatova, I.; Lyushkevich, V.; Haimi, P.; Tamošiūnė, I.; Baniulis, D. Treatment of Common sunflower (Helianthus annus L.) seeds with radio-frequency electromagnetic field and cold plasma induces changes in seed phytohormone balance, seedling development and leaf protein expression. Sci. Rep. 2019, 9, 6437. [Google Scholar] [CrossRef]
- Măgureanu, M.; Sîrbu, R.; Dobrin, D.; Gîdea, M. Stimulation of the germination and early growth of tomato seeds by non-thermal plasma. Plasma Chem. Plasma Process. 2018, 38, 989–1001. [Google Scholar] [CrossRef]
- Hosseini, S.I.; Mohsenimehr, S.; Hadian, J.; Ghorbanpour, M.; Shokri, B. Physico-chemical induced modification of seed germination and early development in artichoke (Cynara scolymus L.) using low energy plasma technology. Phys. Plasmas 2018, 25, 013525. [Google Scholar] [CrossRef]
- Recek, N.; Holc, M.; Vesel, A.; Zaplotnik, R.; Gselman, P.; Mozetic, M.; Primc, G. Germination of Phaseolus vulgaris L. seeds after a short treatment with a powerful RF plasma. Int. J. Mol. Sci. 2021, 22, 6672. [Google Scholar] [CrossRef] [PubMed]
- Dhayal, M.; Lee, S.-Y.; Park, S.-U. Using low-pressure plasma for Carthamus tinctorium L. seed surface modification. Vacuum 2006, 80, 499–506. [Google Scholar] [CrossRef]
- Jiang, J.; He, X.; Li, L.; Li, J.; Shao, H.; Xu, Q.; Ye, R.; Dong, Y. Effect of Cold Plasma Treatment on Seed Germination and Growth of Wheat. Plasma Sci. Technol. 2014, 16, 54–58. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, T.; Farooq, M.; Afsheen, S.; Abrar, M.; Yousaf, M.; Ijaz, M. Cold plasma treatment and laser irradiation of Triticum spp. seeds for sterilization and germination. J. Laser Appl. 2019, 31, 042013. [Google Scholar] [CrossRef]
- Feizollahi, E.; Iqdiam, B.; Vasanthan, T.; Thilakarathna, M.S.; Roopesh, M.S. Effects of Atmospheric-Pressure Cold Plasma Treatment on Deoxynivalenol Degradation, Quality Parameters, and Germination of Barley Grains. Appl. Sci. 2020, 10, 3530. [Google Scholar] [CrossRef]
- Šerý, M.; Zaharonova, A.; Kerdi, A.; Šerá, B. Seed germination of black pine (Pinus nigra Arnold) after diffuse coplanar surface barrier discharge plasma treatment. IEEE Trans. Plasma Sci. 2020, 48, 939–945. [Google Scholar] [CrossRef]
- De Groot, G.J.J.B.; Hundt, A.; Murphy, A.B.; Bange, M.P.; Mai-Prochnow, A. Cold plasma treatment for cotton seed germination improvement. Sci. Rep. 2018, 8, 14372. [Google Scholar] [CrossRef] [Green Version]
- Štepánová, V.; Slavícek, P.; Kelar, J.; Prášil, J.; Smékal, M.; Stupavská, M.; Jurmanová, J.; Cernák, M. Atmospheric pressure plasma treatment of agricultural seeds of cucumber (Cucumis sativus L.) and pepper (Capsicum annuum L.) with effect on reduction of diseases and germination improvement. Plasma Process. Polym. 2018, 15, 1700076. [Google Scholar]
- Tong, J.; He, R.; Zhang, X.; Zhan, R.; Chen, W.; Yang, S. Effects of atmospheric pressure air plasma pretreatment on the seed germination and early growth of Andrographis paniculata. Plasma Sci. Technol. 2014, 16, 260–266. [Google Scholar] [CrossRef] [Green Version]
- Sirgedaitė-Šėžienė, V.; Mildažienė, V.; Žemaitis, P.; Ivankov, A.; Koga, K.; Shiratani, M.; Baliuckas, V. Long-term response of Norway spruce to seed treatment with cold plasma: Dependence of the effects on the genotype. Plasma Proc. Polym. 2020, 18, 2000159. [Google Scholar] [CrossRef]
- Stolárik, T.; Henselová, M.; Martinka, M.; Novák, O.; Zahoranová, A.; Černák, M. Effect of Low-Temperature Plasma on the Structure of Seeds, Growth and Metabolism of Endogenous Phytohormones in Pea (Pisum sativum L.). Plasma Chem. Plasma Proc. 2015, 35, 659–676. [Google Scholar] [CrossRef]
- Švubová, R.; Kyzek, S.; Medvecká, V.; Slováková, L.; Gálová, E.; Zahoranová, A. Novel insight at the effect of cold atmospheric pressure plasma on the activity of enzymes essential for the germination of pea (Pisum sativum L. cv. Prophet) seeds. Plasma Chem. Plasma Process. 2020, 40, 1221–1240. [Google Scholar] [CrossRef]
- Sarinont, T.; Amano, T.; Kitazaki, S.; Koga, K.; Uchida, G.; Shiratani, M.; Hayashi, N. Growth enhancement effects of radish sprouts: Atmospheric pressure plasma irradiation vs. heat shock. J. Phys. Conf. Ser. 2014, 518, 012017. [Google Scholar]
- Sarinont, T.; Amano, T.; Attri, P.; Koga, K.; Hayashi, N.; Shiratani, M. Effects of plasma irradiation using various feeding gases on growth of Raphanus sativus L. Arch. Biochem. Biophys. 2016, 605, 129–140. [Google Scholar]
- Degutytė-Fomins, L.; Paužaitė, G.; Žukienė, R.; Mildažienė, V.; Koga, K.; Shiratani, M. Relationship between cold plasma treatment-induced changes in radish seed germination and phytohormone balance. Jpn. J. Appl. Phys. 2020, 59, SH1001. [Google Scholar] [CrossRef]
- Mihai, A.L.; Dobrin, D.; Popa, M.E.; Mihai, A.L.; Dobrin, D.; Măgureanu, M.; Popa, M.E. Positive effect of non-thermal plasma treatment on radish. Rom. Rep. Phys. 2014, 66, 1110–1117. [Google Scholar]
- Koga, K.; Attri, P.; Kamataki, K.; Itakagi, N.; Shiratani, M.; Mildažienė, V. Impact of radish sprouts seeds coat color on the electron paramagnetic resonance signals after plasma treatment. Jpn. J. Appl. Phys. 2020, 59, SHHF01. [Google Scholar] [CrossRef]
- Attri, P.; Ishikawa, K.; Okumura, T.; Koga, K.; Shiratani, M.; Mildaziene, M. Impact of seed color and storage time on the radish seed germination and sprout growth in plasma agriculture. Sci. Rep. 2021, 11, 2539. [Google Scholar] [CrossRef]
- Yodpitak, S.; Mahatheeranont, S.; Boonyawan, D.; Sookwong, P.; Roytrakul, S.; Norkaew, O. Cold plasma treatment to improve germination and enhance the bioactive phytochemical content of germinated brown rice. Food Chem. 2019, 289, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Suriyasak, C.; Hatanaka, K.; Tanaka, H.; Okumura, T.; Yamashita, D.; Attri, P.; Koga, K.; Shiratani, M.; Hamaoka, N.; Ishibashi, Y. Alterations of DNA methylation caused by cold plasma treatment restore delayed germination of heat-stressed rice (Oryza sativa L.) Seeds. ACS Agric. Sci. Technol. 2021, 1, 5–10. [Google Scholar] [CrossRef]
- Zukiene, R.; Nauciene, Z.; Januskaitiene, I.; Pauzaite, G.; Mildaziene, V.; Koga, K.; Shiratani, M. DBD plasma treatment induced changes in sunflower seed germination, phytohormone balance, and seedling growth. Appl. Phys. Express 2019, 126003. [Google Scholar] [CrossRef]
- Yawirach, S.; Sarapirom, S.; Janpong, K. The effects of dielectric barrier discharge atmospheric air plasma treatment to germination and enhancement growth of sunflower seeds. J. Phys. Conf. Ser. 2019, 1380, 012148. [Google Scholar] [CrossRef]
- Ambrico, P.F.; Šimek, M.; Ambrico, M.; Morano, M.; Prukner, V.; Minafra, A.; Allegretta, I.; Porfido, C.; Senesi, G.S.; Terzano, R. On the air atmospheric pressure plasma treatment effect on the physiology, germination and seedlings of basil seeds. J. Phys. D Appl. Phys. 2019, 53, 104001. [Google Scholar] [CrossRef]
- Sarinont, T.; Amano, T.; Koga, K.; Shiratani, M.; Hayashi, N. Multigeneration effects of plasma irradiation to seeds of Arabidopsis Thaliana and Zinnia on their growth. MRS Online Proc. Libr. Arch. 2014, 1723, 7–11. [Google Scholar] [CrossRef]
- Koga, K.; Thapanut, S.; Amano, T.; Seo, H.; Itagaki, N.; Hayashi, N.; Shiratani, M. Simple method of improving harvest by nonthermal air plasma irradiation of seeds of Arabidopsis thaliana (L.). Appl. Phys. Express 2016, 9, 016201. [Google Scholar] [CrossRef]
- Bafoil, M.; Jemmat, A.; Martinez, Y.; Merbahi, N.; Eichwald, O.; Dunand, C.; Yousfi, M. Effects of low temperature plasmas and plasma activated waters on Arabidopsis thaliana germination and growth. PLoS ONE 2018, 13, e0195512. [Google Scholar] [CrossRef]
- Cui, D.; Yin, Y.; Wang, J.; Wang, Z.; Ding, H.; Ma, R.; Jiao, Z. Research on the physio-biochemical mechanism of non-thermal plasma regulated seed germination and early seedling development in Arabidopsis. Front. Plant Sci. 2019, 10, 1322. [Google Scholar] [CrossRef]
- Zahoranová, A.; Henselová, M.; Hudecová, D.; Kaliňáková, B.; Kováčik, D.; Medvecká, V.; Černák, M. Effect of cold atmospheric pressure plasma on the wheat seedlings vigor and on the inactivation of microorganisms on the seeds surface. Plasma Chem. Plasma Process. 2015, 36, 397–414. [Google Scholar] [CrossRef]
- Guo, Q.; Wang, Y.; Zhang, H.; Qu, G.; Wang, T.; Sun, Q.; Liang, D. Alleviation of adverse effects of drought stress on wheat seed germination using atmospheric dielectric barrier discharge plasma treatment. Sci. Rep. 2017, 7, 16680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, Y.; Oh, K.S.; Oh, J.; Seok, D.C.; Kim, S.B.; Yoo, S.J.; Lee, M.-J. The biological effects of surface dielectric barrier discharge on seed germination and plant growth with barley. Plasma Process. Polym. 2016, 15, 1600056. [Google Scholar] [CrossRef]
- Guragain, R.P.; Baniya, H.B.; Pradhan, S.P.; Dhungana, S.; Chhetri, G.K.; Sedhai, B.; Basnet, N.; Panta, G.P.; Joshi, U.M.; Pandey, B.P.; et al. Impact of non-thermal plasma treatment on the seed germination and seedling development of carrot (Daucus carota sativus L.). J. Phys. Commun. 2021, 5, 125011. [Google Scholar]
- Ji, S.H.; Kim, T.; Panngom, K.; Hong, Y.J.; Pengkit, A.; Park, D.H.; Kang, M.H.; Lee, S.H.; Im, J.S.; Kim, J.S.; et al. Assessment of the effects of nitrogen plasma and plasma-generated nitric oxide on early development of Coriandum sativum. Plasma Proc. Polym. 2015, 12, 1164–1173. [Google Scholar] [CrossRef]
- Billah, M.; Karmakar, S.; Mina, F.B.; Haque, M.N.; Hasan, M.F.; Acharjee, U.K. Investigation of mechanisms involved in seed germination enhancement, enzymatic activity and seedling growth of rice (Oryza Sativa L.) using LPDBD (Ar+Air) plasma. Arch. Biochem. Biophys. 2021, 698, 108726. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Pizá, M.C.; Cejas, E.; Zilli, C.; Prevosto, L.; Mancinelli, B.; Santa-Cruz, D.; Yannarelli, G.; Balestrasse, K. Enhancement of soybean nodulation by seed treatment with non–thermal plasmas. Sci. Rep. 2020, 10, 4917. [Google Scholar]
- Pérez-Pizá, M.C.; Prevosto, L.; Zilli, C.; Cejas, E.; Kelly, H.; Balestrasse, K. Effects of non–thermal plasmas on seed-borne Diaporthe/Phomopsis complex and germination parameters of soybean seeds. Innov. Food Sci. Emerg. 2018, 49, 82–91. [Google Scholar] [CrossRef]
- Zhang, J.J.; Jo, J.O.; Huynh, D.L.; Mongre, R.K.; Ghosh, M.; Singh, A.K.; Lee, S.B.; Mok, Y.S.; Hyuk, P.; Jeong, D.K. Growth-inducing effects of argon plasma on soybean sprouts via the regulation of demethylation levels of energy metabolism-related genes. Sci. Rep. 2017, 7, 41917. [Google Scholar]
- Singh, R.; Kishor, R.; Singh, V.; Singh, V.; Prasad, P.; Aulakh, N.S.; Tiwari, U.K.; Kumar, B. Radio-frequency (RF) room temperature plasma treatment of sweet basil seeds (Ocimum basilicum L.) for germination potential enhancement by immaculation. J. Appl. Res. Med. Aromat. Plants 2022, 26, 100350. [Google Scholar] [CrossRef]
- Rahman, M.M.; Sajib, S.A.; Rahi, M.S.; Tahura, S.; Roy, N.C.; Parvez, S.; Reza, M.A.; Talukder, M.R.; Kabir, A.H. Mechanisms and signaling associated with LPDBD plasma mediated growth improvement in wheat. Sci. Rep. 2018, 8, 10498. [Google Scholar] [CrossRef] [Green Version]
- Meng, Y.; Qu, G.; Wang, T.; Sun, Q.; Liang, D.; Hu, S. Enhancement of germination and seedling growth of wheat seed using dielectric barrier discharge plasma with various gas sources. Plasma Chem. Plasma Proc. 2017, 37, 1105–1119. [Google Scholar] [CrossRef]
- Volkov, A.G.; Hairston, J.S.; Marshall, J.; Bookal, A.; Dholichand, A.; Patel, D. Plasma seeds: Cold plasma accelerates Phaseolus vulgaris seed imbibition, germination, and speed of seedling growth. Plasma Med. 2020, 10, 139–158. [Google Scholar]
- Alves, C., Jr.; de Oliveira Vitoriano, J.; da Silva, D.L.; de Lima Farias, M.; de Lima Dantas, N.B. Water uptake mechanism and germination of Erythrina velutina seeds treated with atmospheric plasma. Sci. Rep. 2016, 6, 33722. [Google Scholar] [CrossRef]
- Fadhlalmawla, S.A.; Mohamed, A.A.H.; Almarashi, J.Q.M.; Boutraa, T. The impact of cold atmospheric pressure plasma jet on seed germination and seedlings growth of fenugreek (Trigonella foenum-graecum). Plasma Sci. Technol. 2019, 21, 105503. [Google Scholar] [CrossRef]
- Zhou, R.; Zhou, R.; Zhang, X.; Zhuang, J.; Yang, S.; Bazaka, K.; Ostrikov, K. Effects of atmospheric-pressure N2, He, air, and O2 microplasmas on mung bean seed germination and seedling growth. Sci. Rep. 2016, 6, 32603. [Google Scholar] [CrossRef] [Green Version]
- Lotfy, K.; Al-Harbi, N.A.; Abd El-Raheem, H. Cold atmospheric pressure nitrogen plasma jet for enhancement germination of wheat seeds. Plasma Chem. Plasma Process. 2019, 39, 897–912. [Google Scholar] [CrossRef]
- Šerá, B.; Gajdovâ, I.; Černák, M.; Gavril, B.; Hnatiuc, E.; Kováčik, D.; Kříha, V.; Sláma, J.; Šerý, M.; Špatenka, P. How various plasma sources may affect seed germination and growth. In Proceedings of the 2012 13th International Conference on Optimization of Electrical and Electronic Equipment (OPTIM), Brasov, Romania, 24–26 May 2012; pp. 1365–1370. [Google Scholar]
- Pawlat, J.; Starek, A.; Sujak, A.; Kwiatkowski, M.; Terebun, P.; Budzen, M. Effects of atmospheric pressure plasma generated in GlidArc reactor on Lavatera thuringiaca L. seeds’ germination. Plasma Process. Polym. 2018, 15, 1700064. [Google Scholar]
- Sera, B.; Sery, M.; Gavril, B.; Gajdova, I. Seed Germination and Early Growth Responses to Seed Pre-Treatment by Non-Thermal Plasma in Hemp Cultivars (Cannabis sativa L.). Plasma Chem. Plasma Process. 2017, 37, 207–221. [Google Scholar]
- Šerá, B.; Vanková, R.; Rohácek, K.; Šerý, M. Gliding arc plasma treatment of maize (Zea mays L.) grains promotes seed germination and early growth, affecting hormone pools, but not significantly photosynthetic parameters. Agronomy 2021, 11, 2066. [Google Scholar] [CrossRef]
- Ivankov, A.; Naučienė, Z.; Degutytė-Fomins, L.; Žūkienė, R.; Januškaitienė, I.; Malakauskienė, A.; Jakstas, V.; Ivanauskas, L.; Romanovskaja, D.; Slepetiene, A.; et al. Changes in agricultural performance of common buckwheat induced by seed treatment with cold plasma and electromagnetic Field. Appl. Sci. 2021, 11, 4391. [Google Scholar] [CrossRef]
- Staric, P.; Grobelnik Mlakar, S.; Junkar, I. Response of Two Different Wheat Varieties to Glow and Afterglow Oxygen Plasma. Plants 2021, 10, 1728. [Google Scholar] [CrossRef] [PubMed]
- Zahoranová, A.; Hoppanová, L.; Šimoncicová, J.; Tuceková, Z.; Medvecká, V.; Hudecová, D.; Kalináková, B.; Kovácik, D.; Cernák, M. Effect of Cold Atmospheric Pressure Plasma on Maize Seeds: Enhancement of Seedlings Growth and Surface Microorganisms Inactivation. Plasma Chem. Plasma Process. 2018, 38, 969–988. [Google Scholar]
- Šerá, B.; Šerý, M.; Zahoranová, A.; Tomekova, J. Germination improvement of three pine species (Pinus) after diffuse coplanar surface barrier discharge plasma treatment. Plasma Chem. Plasma Process. 2021, 41, 211–226. [Google Scholar] [CrossRef]
- Paužaitė, G.; Malakauskienė, A.; Naučienė, Z.; Žūkienė, R.; Filatova, I.; Lyushkevich, V.; Azarko, I.; Mildažienė, V. Changes in Norway spruce germination and growth induced by pre-sowing seed treatment with cold plasma and electromagnetic field: Short-term versus long-term effects. Plasma Process. Polym. 2018, 15, 1700068. [Google Scholar] [CrossRef]
- Mravlje, J.; Regvar, M.; Staric, P.; Mozetic, M.; Vogel-Mikuš, K. Cold plasma affects germination and fungal community structure of buckwheat seeds. Plants 2021, 10, 85. [Google Scholar] [CrossRef]
- Ahn, C.; Gill, J.; Ruzic, D.N. Growth of plasma-treated corn seeds under realistic conditions. Sci. Rep. 2019, 9, 4355. [Google Scholar] [CrossRef] [Green Version]
- Šerá, B.; Gajdová, I.; Šerý, M.; Špatenka, P. New physicochemical treatment method of poppy seeds for agriculture and food industries. Plasma Sci. Technol. 2013, 15, 935–938. [Google Scholar]
- Tomeková, J.; Kyzek, S.; Medvecká, V.; Gálová, E.; Zahoranová, A. Influence of cold atmospheric pressure plasma on pea seeds: DNA damage of seedlings and optical diagnostics of plasma. Plasma Chem. Plasma Process. 2020, 40, 1571–1584. [Google Scholar] [CrossRef]
- Maruyama-Nakashita, A.; Ishibashi, Y.; Yamamoto, K.; Liu Zhang, L.; Morikawa-Ichinose, T.; Sun-Ju Kim, S.-J.; Hayashi, N. Oxygen plasma modulates glucosinolate levels without affecting lipid contents and composition in Brassica napus seeds. Biosci. Biotechnol. Biochem. 2021, 85, 2434–2441. [Google Scholar] [CrossRef]
- Iranbakhsh, A.; Ghoranneviss, M.; Oraghi Ardebili, Z.; Oraghi Ardebili, N.; Hesami Tackallou, S.; Nikmaram, H. Non-thermal plasma modified growth and physiology in Triticum aestivum via generated signaling molecules and UV radiation. Biol. Plantarum 2016, 61, 702–708. [Google Scholar] [CrossRef]
- Bußler, S.; Herppich, W.B.; Neugart, S.; Schreiner, M.; Ehlbeck, J.; Rohn, S.; Schlüter, O. Impact of cold atmospheric pressure plasma on physiology and flavonol glycoside profile of peas (Pisum sativum “Salamanca”). Food Res. Int. 2015, 76, 132–141. [Google Scholar] [CrossRef]
- Matilla, A.; Matilla, A.; Gallardo, M.; Puga-Hermida, M.I. Structural, physiological and molecular aspects of heterogeneity in seeds: A review. Seed Sci. Res. 2005, 15, 63–76. [Google Scholar] [CrossRef]
- Bhattacharjee, S. ROS and oxidative stress: Origin and implication. In Reactive Oxygen Species in Plant Biology; Springer Nature India Private Limited: Dheli, India, 2019. [Google Scholar] [CrossRef]
- Wahid, A.; Farooq, M.; Siddique, K.H.M. Implications of Oxidative Stress for Crop Growth and Productivity. In Handbook of Plant and Crop Physiology, 3rd ed.; Pessarakli, M., Ed.; Taylor and Francis Group: Boca Raton, FL, USA, 2014; pp. 549–556. [Google Scholar] [CrossRef]
- Hong, S.-H.; Szili, E.J.; Jenkins, A.T.A.; Short, R.D. Ionized gas (plasma) delivery of reactive oxygen species (ROS) into artificial cells. J. Phys. D Appl. Phys. 2014, 47, 362001. [Google Scholar] [CrossRef] [Green Version]
- Lepiniec, L.; Debeaujon, I.; Routaboul, J.-M.; Baudry, A.; Pourcel, L.; Nesi, N.; Caboche, M. Genetics and biochemistry of seed flavonoids. Ann. Rev. Plant Biochem. 2006, 57, 405–430. [Google Scholar] [CrossRef]
- Zhou, W.; Chen, F.; Luo, X.; Dai, Y.; Yang, Y.; Zheng, C.; Yang, W.; Shu, K. A matter of life and death: Molecular, physiological and environmental regulation of seed longevity. Plant Cell Environ. 2020, 43, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Leprince, O.; Pellizzaro, A.; Berriri, S.; Buitink, J. Late seed maturation: Drying without dying. J. Exp. Bot. 2017, 68, 827–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojtyla, Ł.; Lechowska, K.; Kubala, S.; Garnczarska, M. Different modes of hydrogen peroxide action during seed germination. Front. Plant Sci. 2016, 7, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandiselvam, R.; Mayookha, V.P.; Kothakota, A.; Sharmila, L.; Ramesh, S.V.; Bharathi, C.P.; Gomathy, K.; Srikanth, V. Impact of ozone treatment on seed germination–a systematic review. Ozone: Sci. Eng 2020, 42, 331–346. [Google Scholar] [CrossRef]
- Neyts, E.C. Plasma-surface interactions in plasma catalysis. Plasma Chem Plasma P 2016, 36, 185–212. [Google Scholar] [CrossRef]
- Bazaka, K.; Jacob, M.V.; Crawford, R.J.; Ivanova, E.P. Plasma-assisted surface modification of organic biopolymers to prevent bacterial attachment. Acta Biomater. 2011, 7, 2015–2028. [Google Scholar] [CrossRef]
- Homola, T.; Prukner, V.; Artemenko, A.; Hanuš, J.; Kylián, O.; Šimek, M. Direct treatment of pepper (Capsicum annuum L.) and melon (Cucumis melo) seeds by amplitude-modulated dielectric barrier discharge in air. J. Appl. Phys. 2021, 129, 193303. [Google Scholar] [CrossRef]
- Molina, R.; López-Santos, C.; Gómez-Ramírez, A.; Vílchez, A.; Espinós, J.P.; González-Elipe, A.R. Influence of irrigation conditions in the germination of plasma treated nasturtium seeds. Sci. Rep. 2018, 8, 16442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.H.; Kim, H.-J.; Woo, K.S.; Jo, C.; Kim, J.-K.; Kim, S.H.; Park, H.Y.; Oh, S.-K.; Kim, W.H. Evaluation of cold plasma treatments for improved microbial and physicochemical qualities of brown rice. LWT-Food Sci. Technol. 2016, 73, 442–447. [Google Scholar] [CrossRef]
- Shapira, Y.; Multanen, V.; Whyman, G.; Bormashenko, Y.; Chaniel, G.; Barkay, Z.; Bormashenko, E. Plasma Treatment Switches the Regime of Wetting and Floating of Pepper Seeds. Colloid Surf. B 2017, 157, 417–423. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Nikolaeva, M.G. Factors controlling the seed dormancy pattern. In The Physiology and Biochemistry of Seed Dormancy and Germination; Khan, A.A., Ed.; North-Holland Publishing Co.: Amsterdam, The Netherlands, 1977; pp. 51–74. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: New York, NY, USA, 2014; pp. 37–77. [Google Scholar]
- Graeber, K.; Nakabayashi, K.; Miatton, E.; Leubner-Metzger, G.; Soppe, W.J.J. Molecular mechanisms of seed dormancy. Plant Cell Environ. 2012, 35, 1769–1786. [Google Scholar] [CrossRef] [PubMed]
- Yan, A.; Chen, Z. The pivotal role of abscisic acid signaling during transition from seed maturation to germination. Plant Cell Rep. 2017, 36, 689–703. [Google Scholar] [CrossRef]
- Shu, K.; Liu, X.D.; Xie, Q.; He, Z.H. Two faces of one seed: Hormonal regulation of dormancy and germination. Mol. Plant. 2016, 9, 34–45. [Google Scholar] [CrossRef] [Green Version]
- Seo, M.; Jikumaru, Y.; Kamiya, Y. Profiling of hormones and related metabolites in seed dormancy and germination studies. Methods Mol. Biol. 2011, 773, 99–111. [Google Scholar] [CrossRef]
- Arc, E.; Sechet, J.; Corbineau, F.; Rajjou, L.; Marion-Poll, A. ABA crosstalk with ethylene and nitric oxide in seed dormancy and germination. Front. Plant Sci. 2013, 4, 63. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Yu, D. BRASSINOSTEROID INSENSITIVE2 interacts with ABSCISIC ACID INSENSITIVE5 to mediate the antagonism of brassinosteroids to abscisic acid during seed germination in Arabidopsis. Plant Cell 2014, 26, 4394–4408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohli, A.; Sreenivasulu, N.; Lakshmanan, P.; Kumar, P.P. The phytohormone crosstalk paradigm takes center stage in understanding how plants respond to abiotic stresses. Plant Cell Rep. 2013, 32, 945–957. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Wang, M.; Wang, H.; Zhang, W.-H.; Mao, P. Physiological and proteomic analyses for seed dormancy and release in the perennial grass of Leymus chinensis. Environ. Exp. Bot. 2019, 162, 95–102. [Google Scholar] [CrossRef]
- Ji, S.H.; Choi, K.H.; Pengkit, A.; Im, J.S.; Kim, J.S.; Kim, Y.H.; Park, Y.; Hong, E.J.; Jung, S.K.; Choi, E.H.; et al. Effects of high voltage nanosecond pulsed plasma and micro DBD plasma on seed germination, growth development and physiological activities in spinach. Arch. Biochem. Biophys. 2016, 605, 117–128. [Google Scholar] [CrossRef]
- Mildažienė, V.; Ivankov, A.; Paužaitė, G.; Naučienė, Z.; Žūkienė, R.; Degutytė-Fomins, L.; Pukalskas, A.; Venskutonis, P.R.; Filatova, I.; Lyuskevich, V. Seed treatment with cold plasma and electromagnetic field induces changes in red clover root growth dynamics, flavonoid exudation, and activates nodulation. Plasma Proc. Polym. 2020, 18, 2000160. [Google Scholar]
- Hansen, C.C.; Nelson, D.R.; Møller, B.L.; Werck-Reichhart, D. Plant cytochrome P450 plasticity and evolution. Molecular Plant 2021, 14, 1244–1265. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Waszczak, C.; Carmody, M.; Kangasjärvi, J. Reactive Oxygen Species in Plant Signaling. Ann. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef] [Green Version]
- Skalak, J.; Nicolas, K.L.; Vankova, R.; Hejatko, J. Signal integration in plant abiotic stress responses via multistep phosphorelay signaling. Front. Plant Sci. 2021, 12, 644823. [Google Scholar] [CrossRef]
- Arasimowicz, M.; Floryszak-Wieczorek, J. Nitric oxide as a bioactive signalling molecule in plant stress responses. Plant Sci. 2007, 172, 876–887. [Google Scholar]
- Santisree, P.; Adimulam, S.S.; Sharma, K.; Bhatnagar-Mathur, P.; Sharma, K.K. Insights into the Nitric Oxide Mediated Stress Tolerance in Plants. In Plant Signaling Molecules; Khan, M.I.R., Reddy, P.S., Ferrante, A., Khan, N.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 385–406. [Google Scholar]
- Bailly, C. The signalling role of ROS in the regulation of seed germination and dormancy. Biochem. J. 2019, 476, 3019–3032. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhu, J.-K.; Lang, Z. Nitric oxide suppresses the inhibitory effect of abscisic acid on seed germination by S-nitrosylation of SnRK2 proteins. Plant Sign. Behav. 2015, 10, e1031939. [Google Scholar] [CrossRef] [Green Version]
- Bahin, E.; Bailly, C.; Sotta, B.; Kranner, I.; Corbineau, F.; Leymarie, J. Crosstalk between reactive oxygen species and hormonal signalling pathways regulates grain dormancy in barley. Plant Cell Environ. 2011, 34, 980–993. [Google Scholar] [CrossRef] [PubMed]
- Oracz, K.; Karpinski, S. Phytohormones signaling pathways and ROS involvement in seed germination. Front. Plant Sci. 2016, 7, 864. [Google Scholar] [CrossRef] [Green Version]
- Zandalinas, S.I.; Mittler, R. ROS-induced ROS release in plant and animal cells. Free Rad. Biol. Med. 2017, 122, 21–27. [Google Scholar] [CrossRef]
- Khan, M.M.; Hendry, G.A.F.; Atherton, N.M.; Vertucci-Walters, C.W. Free radical accumulation and lipid peroxidation in testas of rapidly aged soybean seeds: A light-promoted process. Seed Sci. Res. 1996, 6, 101–107. [Google Scholar] [CrossRef]
- Hepburn, H.A.; Goodman, B.A.; McPhail, D.B.; Matthews, S.; Powell, A.A. An Evaluation of EPR Measurements of the Organic Free Radical Content of Individual Seeds in the Non-destructive Testing of Seed Viability. J. Exp. Bot. 1986, 37, 1675–1684. [Google Scholar] [CrossRef]
- Zhang, H.; Lang, Z.; Zhu, J.-K. Dynamics and function of DNA methylation in plants. Nat. Rev. Mol. Cell Biol. 2018, 19, 489–506. [Google Scholar] [CrossRef]
- Wang, J.; Cui, D.; Wang, L.; Du, M.; Yin, Y.; Ma, R.; Sun, H.; Jiao, Z. Atmospheric pressure plasma treatment induces abscisic acid production, reduces stomatal aperture and improves seedling growth in Arabidopsis thaliana. Plant Biol. 2021, 23, 564–573. [Google Scholar] [CrossRef]
- Cui, D.J.; Yin, Y.; Li, H.D.; Hu, X.X.; Zhuang, J.; Ma, R.N.; Jiao, Z. Comparative transcriptome analysis of atmospheric pressure cold plasma enhanced early seedling growth in Arabidopsis thaliana. Plasma Sci. Technol. 2021, 23, 085502. [Google Scholar] [CrossRef]
- Seddighinia, F.S.; Iranbakhsh, A.; Ardebili, Z.O.; Soleimanpour, S. Seed-priming with cold plasma and supplementation of nutrient solution with carbon nanotube enhanced carotenoid contents and the expression of psy and pds in Bitter melon (Momordica charantia). J. Appl. Bot. Food Qual. 2021, 94, 7–14. [Google Scholar] [CrossRef]
- Ghaemi, M.; Majd, A.; Iranbakhsh, A. Transcriptional responses following seed priming with cold plasma and electromagnetic field in Salvia nemorosa L. J. Theor. Appl. Phys. 2020, 14, 323–328. [Google Scholar] [CrossRef]
- Iranbakhsh, A.; Ardebili, Z.O.; Molaei, H.; Ardebili, N.O.; Amini, M. Cold plasma up-regulated expressions of WRKY1 transcription factor and genes involved in biosynthesis of cannabinoids in hemp (Cannabis sativa L.). Plasma Chem Plasma Process. 2020, 40, 527–537. [Google Scholar] [CrossRef]
- Holubova, L.; Svubova, R.; Slovakova, L.; Bokor, B.; Krockova, V.C.; Rencko, J.; Uhrin, F.; Medvecka, V.; Zahoranova, A.; Galova, E. Cold atmospheric pressure plasma treatment of maize grains-induction of growth, enzyme activities and heat shock proteins. Int. J. Mol. Sci. 2021, 22, 8509. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Yu, N.N.; Zheng, W.; Zhang, L.N.; Liu, Y.; Yu, J.B.; Zhang, Y.Q.; Park, G.; Sun, H.N.; Kwon, T. Effect of non-thermal plasma (NTP) on common sunflower (Helianthus annus L.) seed growth via upregulation of antioxidant activity and energy metabolism-related gene expression. Plant Growth Regul. 2021, 95, 271–281. [Google Scholar] [CrossRef]
- Tamosiune, I.; Gelvonauskiene, D.; Haimi, P.; Mildaziene, V.; Koga, K.; Shiratani, M.; Baniulis, D. Cold plasma treatment of sunflower seeds modulates plant-associated microbiome and stimulates root and lateral organ growth. Front. Plant Sci. 2020, 11, 1347. [Google Scholar] [CrossRef]
- Li, K.; Zhong, C.S.; Shi, Q.H.; Bi, H.G.; Gong, B. Cold plasma seed treatment improves chilling resistance of tomato plants through hydrogen peroxide and abscisic acid signaling pathway. Free Rad. Biol. Med. 2021, 172, 286–297. [Google Scholar] [CrossRef]
- Adhikari, B.; Adhikari, M.; Ghimire, B.; Adhikari, B.C.; Park, G.; Choi, E.H. Cold plasma seed priming modulates growth, redox homeostasis and stress response by inducing reactive species in tomato (Solanum lycopersicum). Free Radical Bio. Med. 2020, 156, 57–69. [Google Scholar] [CrossRef]
- Li, K.; Zhang, L.; Shao, C.; Zhong, C.; Cao, B.; Shi, Q.; Gong, B. Utilising cold plasma seed treatment technologies to delay cotyledon senescence in tomato seedlings. Sci. Horticult. 2021, 281, 109911. [Google Scholar] [CrossRef]
- Iranbakhsh, A.; Ardebili, N.O.; Ardebili, Z.O.; Shafaati, M.; Ghoranneviss, M. Non-thermal plasma Induced expression of heat shock factor A4A and improved wheat (Triticum aestivum L.) growth and resistance against salt stress. Plasma Chem. Plasma Process. 2018, 38, 29–44. [Google Scholar] [CrossRef]
- Hasan, M.; Sohan, M.; Sajib, S.A.; Hossain, M.; Miah, M.; Maruf, M.; Khalid-Bin-Ferdaus, K.M.; Kabir, A.H.; Talukder, M.R.; Rashid, M.M.; et al. The effect of low-pressure dielectric barrier discharge (LPDBD) plasma in boosting fermination, growth, and nutritional properties in wheat. Plasma Chem. Plasma Process. 2022, 42, 339–362. [Google Scholar] [CrossRef]
- Moghanloo, M.; Iranbakhsh, A.; Ebadi, M.; Satari, T.N.; Ardebili, Z.O. Seed priming with cold plasma and supplementation of culture medium with silicon nanoparticle modified growth, physiology, and anatomy in Astragalus Fridae as an endangered species. Acta Physiol. Plant 2019, 41, 54. [Google Scholar] [CrossRef]
- Babajani, A.; Iranbakhsh, A.; Ardebili, Z.O.; Eslami, B. Seed priming with non-thermal plasma modified plant reactions to selenium or zinc oxide nanoparticles: Cold plasma as a novel emerging tool for plant science. Plasma Chem. Plasma Prosess. 2019, 39, 21–34. [Google Scholar] [CrossRef]
- Wu, Z.H.; Chi, L.H.; Bian, S.F.; Xu, K.Z. Effects of plasma treatment on maize seeding resistance. J. Maize Sci. 2007, 15, 111–113. [Google Scholar]
- Henselová, M.; Slováková, Ľ.; Martinka, M.; Zahoranová, A. Growth, anatomy and enzyme activity changes in maize roots induced by treatment of seeds with low-temperature plasma. Biologia 2012, 67, 490–497. [Google Scholar] [CrossRef]
- Karmakar, S.; Billah, M.; Hasan, M.; Sohan, S.R.; Hossain, M.F.; Hoque, K.M.F.; Kabir, A.H.; Rashid, M.M.; Talukder, M.R.; Reza, M.A. Impact of LFGD (Ar+O2) plasma on seed surface, germination, plant growth, productivity and nutritional composition of maize (Zea mays L.). Heliyon 2021, 7, e06458. [Google Scholar] [CrossRef]
- Iranbakhsh, A.; Ardebili, Z.O.; Ardebili, N.O.; Ghoranneviss, M.; Safari, N. Cold plasma relieved toxicity signs of nano zinc oxide in Capsicum Annuum Cayenne via modifying growth, differentiation, and physiology. Acta Physiol. Plant 2018, 40, 154. [Google Scholar] [CrossRef]
- Sheteiwy, M.S.; An, J.; Yin, M.; Jia, X.; Guan, Y.; He, F.; Hu, J. Cold plasma treatment and exogenous salicylic acid priming enhances salinity tolerance of Oryza sativa seedlings. Protoplasma 2019, 256, 79–99. [Google Scholar] [CrossRef]
- Singh, R.; Prasad, P.; Mohan, R.; Verma, M.K.; Kumar, B. Radiofrequency cold plasma treatment enhances seed germination and seedling growth in variety CIM-Saumya of sweet basil (Ocimum basilicum L.). J. Appl. Res. Med. Arom. Plants 2019, 12, 78–81. [Google Scholar] [CrossRef]
- Zhang, B.; Li, R.; Yan, J. Study on activation and improvement of crop seeds by the application of plasma treating seeds equipment. Arch. Biochem. Biophys. 2018, 655, 37–42. [Google Scholar] [CrossRef]
- Kabir, A.H.; Rahman, M.M.; Das, U.; Sarkar, U.; Roy, N.C.; Reza, M.A.; Talukder, M.R.; Uddin, M.A. Reduction of cadmium toxicity in wheat through plasma technology. PLoS ONE 2019, 14, e0214509. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, T.; Meng, Y.; Qu, G.; Sun, Q.; Liang, D.; Hu, S. Air atmospheric dielectric barrier discharge plasma induced germination and growth enhancement of wheat seed. Plasma Chem. Plasma Process. 2017, 37, 1621–1634. [Google Scholar] [CrossRef]
- Saravana Kumar, R.M.; Wang, Y.; Zhang, X.; Cheng, H.; Sun, L.; He, S.; Hao, F. Redox components: Key regulators of epigenetic modifications in plants. Int. J. Mol. Sci. 2020, 21, 1419. [Google Scholar] [CrossRef] [Green Version]
- Sera, B.; Spatenka, P.; Sery, M.; Vrchotova, N.; Hruskova, I. Influence of plasma treatment on wheat and oat germination and early growth. IEEE Transact. Plasma Sci. 2010, 38, 2963–2968. [Google Scholar] [CrossRef]
- Pérez-Pizá, M.C.; Prevosto, L.; Grijalba, P.E.; Zilli, C.G.; Cejas, E.; Mancinelli, B.; Balestrasse, K.B. Improvement of growth and yield of soybean plants through the application of non-thermal plasmas to seeds with different health status. Heliyon 2019, 54, e01495. [Google Scholar] [CrossRef] [Green Version]
- Mildažienė, V.; Paužaitė, G.; Naučienė, Z.; Malakauskiene, A.; Žūkienė, R.; Januškaitienė, I.; Jakštas, V.; Ivanauskas, L.; Filatova, I.; Lyushkevich, V. Pre-sowing seed treatment with cold plasma and des electromagnetic field increases secondary metabolite content in purple coneflower (Echinacea purpurea) leaves. Plasma Process. Polym. 2018, 14, 1700059. [Google Scholar]
- Saberi, M.; Modarres-Sanavy, S.A.M.; Zare, R.; Ghomi, H. Amelioration of photosynthesis and quality of wheat under non-thermal radio frequency plasma treatment. Sci. Rep. 2018, 8, 11655. [Google Scholar] [CrossRef] [Green Version]
- Filatova, I.; Lyushkevich, V.; Goncharik, S.; Zhukovsky, A.; Krupenko, N.; Kalatskaja, J. The effect of low-pressure plasma treatment of seeds on the plant resistance to pathogens pathogens and crop yields. J. Phys. D Appl. Phys. 2020, 53, 244001. [Google Scholar] [CrossRef]
- Nedved, H.; Kalatskaja, J.; Kopylova, N.; Herasimovich, K.; Rybinskaya, E.; Vusik, N.; Filatova, I.; Lyushkevich, V.; Laman, N. Short-term and long-term effects of plasma and radio wave seed treatments on red clover plants. IOP Conf. Ser. Earth Environ. Sci. 2021, 937, 022137. [Google Scholar] [CrossRef]
- Tamosiune, I.; Gelvonauskiene, D.; Ragauskaite, L.; Koga, K.; Shiratani, M.; Baniulis, D. Cold plasma treatment of Arabidopsis thaliana (L.) seeds modulates plant-associated microbiome composition. Appl. Phys. Express 2020, 13, 076001. [Google Scholar] [CrossRef]
- Holc, M.; Primc, G.; Iskra, J.; Titan, P.; Kovač, J.; Mozetič, M.; Junkar, I. Effect of oxygen plasma on sprout and root growth, surface morphology and yield of garlic. Plants 2019, 8, 462. [Google Scholar] [CrossRef] [Green Version]
- Cianioti, S.; Katsenios, N.; Efthimiadou, A.; Stergiou, P.; Xanthou, Z.; Giannoglou, M.; Dimitrakellis, P.; Gogolides, E.; Katsaros, G. Pre-sowing treatment of maize seeds by cold atmospheric plasma and pulsed electromagnetic fields: Effect on plant and kernels characteristics. Aust. J. Crop Sci. 2021, 15, 251–259. [Google Scholar] [CrossRef]
- Roy, N.C.; Hasan, M.M.; Talukder, M.R.; Hossain, M.D.; Chowdhury, A.N. Prospective applications of low frequency glow discharge plasmas on enhanced germination, growth and yield of wheat. Plasma Chem. Plasma Process. 2018, 38, 13–28. [Google Scholar] [CrossRef]
- Li, L.; Li, J.; Shen, M.; Hou, J.; Shao, H.; Dong, Y.; Jiang, J. Improving seed germination and peanut yields by cold plasma treatment. Plasma Sci. Technol. 2016, 18, 1027–1033. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Huang, Y.; Yang, S.; Chen, W. Introduction of a new atmospheric pressure plasma device and application on tomato seeds. Agricult. Sci. 2011, 2, 23–27. [Google Scholar] [CrossRef] [Green Version]
- Yoon, Y.; Seo, D.H.; Shin, H.; Kim, H.J.; Kim, C.M.; Jang, G. The role of stress-responsive transcription factors in modulating abiotic stress tolerance in plants. Agronomy 2020, 10, 788. [Google Scholar] [CrossRef]
- Heldt, H.-W.; Piechulla, B. Phenylpropanoids comprise a multitude of plant-specialized metabolites and cell wall components. In Plant Biochemistry, 5th ed.; Academic Press: London, UK, 2021; pp. 411–429. [Google Scholar]
- Bai, Y.; Sunarti, S.; Kissoudis, C.; Visser, R.G.F.; van der Linden, C.G. The role of tomato WRKY genes in plant responses to combined abiotic and biotic stresses. Front. Plant Sci. 2018, 9, 801. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T.; Fujita, Y.; Sayama, H.; Kidokoro, S.; Maruyama, K.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AREB1, AREB2, and ABF3 are master transcription factors that cooperatively regulate ABRE-dependent ABA signaling involved in drought stress tolerance and require ABA for full activation. Plant J. 2010, 61, 672–685. [Google Scholar] [CrossRef]
- Lee, Y.K.; Lim, J.; Hong, E.J.; Kim, S.B. Plasma-activated water regulates root hairs and cotyledon size dependent on cell elongation in Nicotiana tabacum L. Plant Biotechnol. Rep. 2020, 14, 663–672. [Google Scholar] [CrossRef]
- Ghasempour, M.; Iranbakhsh, A.; Ebadi, M.; Oraghi Ardebili, Z. Seed priming with cold plasma improved seedling performance, secondary metabolism, and expression of deacetylvindoline O-acetyltransferase gene in Catharanthus roseus. Contrib. Plasm. Phys. 2020, 60, e201900159. [Google Scholar] [CrossRef]
- Dezest, M.; Chavatte, L.; Bourdens, M.; Quinton, D.; Camus, M.; Garrigues, L.; Descargues, P.; Arbault, S.; Burlet-Schiltz, O.; Casteilla, L.; et al. Mechanistic insights into the impact of cold atmospheric pressure plasma on human epithelial cell lines. Sci. Rep. 2017, 7, 41163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scharf, C.; Eymann, C.; Emicke, P.; Bernhardt, J.Ã.; Wilhelm, M.; Görries, F.; Winter, J.; von Woedtke, T.; Darm, K.; Daeschlein, G.; et al. Improved wound healing of airway epithelial cells is mediated by cold atmospheric plasma: A time course-related proteome Analysis. Oxid. Med. Cell. Longev. 2019, 2019, 7071536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hideg, Ã.V.; Jansen, M.A.K.; Strid, Ã.K. UV-B exposure, ROS, and stress: Inseparable companions or loosely linked associates? Trends Plant Sci. 2013, 18, 107–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Suh, D.; Tang, T.; Lee, H.J.; Roe, J.S.; Kim, G.C.; Han, S.; Song, K. Non-thermal atmospheric pressure plasma induces epigenetic modifications that activate the expression of various cytokines and growth factors in human mesoderm-derived stem cells. Free Rad. Biol. Med. 2020, 148, 108–122. [Google Scholar] [CrossRef] [PubMed]
- Socco, S.; Bovee, R.C.; Palczewski, M.B.; Hickok, J.R.; Thomas, D.D. Epigenetics: The third pillar of nitric oxide signaling. Pharmacol. Res. 2017, 121, 52–58. [Google Scholar] [CrossRef]
- Lindermayr, C.; Rudolf, E.E.; Durner, J.Ã.; Groth, M. Interactions between metabolism and chromatin in plant models. Mol. Metab. 2020, 38, 100951. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, B.; Pangomm, K.; Veerana, M.; Mitra, S.; Park, G. Plant disease control by non-thermal atmospheric-pressure plasma. Front. Plant Sci. 2020, 11, 77. [Google Scholar] [CrossRef]
- Lacchini, E.; Goossens, A. Combinatorial control of plant specialized metabolism: Mechanisms, functions, and consequences. Ann. Rev. Cell Dev. Bi. 2020, 36, 291–313. [Google Scholar] [CrossRef]
- Wink, M. Secondary metabolites, the role in plant diversification of. In Encyclopedia of Evolutionary Biology, 1st ed.; Kliman, R., Ed.; Elsevier Inc.: Oxford, UK, 2016; Volume 4, pp. 1–9. [Google Scholar]
- Selmar, D.; Kleinwächter, M. Stress enhances the synthesis of secondary plant products: The impact of stress-related over-reduction on the accumulation of natural products. Plant Cell Physiol. 2013, 54, 817. [Google Scholar]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Env. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Sewelam, N.; Kazan, K.; Schenk, P.M. Global plant stress signaling: Reactive oxygen species at the cross-road. Front. Plant Sci. 2016, 7, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant salinity stress: Many unanswered questions remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Dubey, S.; Shri, M.; Gupta, A.; Rani, V.; Chakrabarty, D. Toxicity and detoxification of heavy metals during plant growth and metabolism. Environ. Chem. Lett. 2018, 16, 1169–1192. [Google Scholar] [CrossRef]
- Feng, J.; Wang, D.; Shao, C.; Zhang, L.; Tang, X. Effects of cold plasma treatment on alfalfa seed growth under simulated drought stress. Plasma Sci. Technol. 2018, 20, 035505. [Google Scholar] [CrossRef] [Green Version]
- Kyzek, S.; Holubová, Ľ.; Medvecká, V.; Tomeková, J.; Gálová, E.; Zahoranová, A. Cold atmospheric pressure plasma can induce adaptive response in pea seeds. Plasma Chem Plasma Processing 2019, 39, 475–486. [Google Scholar] [CrossRef]
- Jiang, J.; Lu, Y.; Li, J.; Li, L.; He, X.; Shao, H.; Dong, Y. Effect of seed treatment by cold plasma on the resistance of tomato to Ralstonia solanacearum (Bacterial Wilt). PLoS ONE 2014, 9, e97753. [Google Scholar] [CrossRef] [Green Version]
- De Geyter, N.; Morent, R. Nonthermal plasma sterilization of living and nonliving surfaces. Annu. Rev. Biomed. Eng. 2012, 14, 255–274. [Google Scholar] [CrossRef]
- Patil, S.; Bourke, P.; Cullen, P.J. Principles of nonthermal plasma decontamination. In Cold Plasma in Food and Agriculture: Fundamentals and Applications; Misra, N.N., Schlutter, O., Culle, P.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 143–177. [Google Scholar]
- Bourke, P.; Ziuzina, D.; Han, L.; Cullen, P.J.; Gilmore, B.F. Microbiological interactions with cold plasma. J. Appl. Microbiol. 2017, 123, 308–324. [Google Scholar] [CrossRef] [Green Version]
- Niedzwiedz, I.; Wasko, A.; Pawlat, J.; Polak-Berecka, M. The state of research on antimicrobial activity of cold plasma. Pol. J. Microbiol. 2019, 68, 153–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van, I.J.; Smet, C.; Tiwari, B.; Greiner, R.; Ojha, S.; Stulic, V.; Vukušić, T.; Režek Jambrak, A. State of the art of nonthermal and thermal processing for inactivation of micro-organisms. J. Appl. Microbiol. 2018, 125, 16–35. [Google Scholar] [CrossRef] [Green Version]
- Khamsen, N.; Onwimol, D.; Teerakawanich, N.; Dechanupaprittha, S.; Kanokbannakorn, W.; Hongesombut, K.; Srisonphan, S. Rice (Oryza sativa L.) seed sterilization and germination enhancement via atmospheric hybrid nonthermal discharge plasma. ACS Appl. Mater. Interfaces 2016, 8, 19268–19275. [Google Scholar] [CrossRef] [PubMed]
- Simoncicova, J.; Krystofova, S.; Medvecka, V.; Durisova, K.; Kalinakova, B. Technical applications of plasma treatments: Current state and perspectives. Appl. Microbiol. Biotechnol. 2019, 103, 5117–5129. [Google Scholar] [CrossRef]
- Waskow, A.; Betschart, J.; Butscher, D.; Oberbossel, G.; Kloti, D.; Buttner-Mainik, A.; Adamcik, J.; von Rohr, P.R.; Schuppler, M. Characterization of efficiency and mechanisms of cold atmospheric pressure plasma decontamination of seeds for sprout production. Front. Microbiol. 2018, 9, 3164. [Google Scholar] [CrossRef] [Green Version]
- Nishioka, T.; Takai, Y.; Kawaradani, M.; Okada, K.; Tanimoto, H.; Misawa, T.; Kusakari, S. Seed disinfection effect of atmospheric pressure plasma and low pressure plasma on Rhizoctonia solani. Biocontrol. Sci. 2014, 19, 99–102. [Google Scholar] [CrossRef] [Green Version]
- Nishioka, T.; Takai, Y.; Mishima, T.; Kawaradani, M.; Tanimoto, H.; Okada, K.; Misawa, T.; Kusakari, S. Low-pressure plasma application for the inactivation of the seed-borne pathogen Xanthomonas campestris. Biocontrol. Sci. 2016, 21, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Kordas, L.; Pusz, W.; Czapka, T.; Kacprzyk, R. The effect of low-temperature plasma on fungus colonization of winter wheat grain and seed quality. Pol. J. Environ. Stud. 2015, 24, 433–438. [Google Scholar]
- Barret, M.; Briand, M.; Bonneau, S.; Preveaux, A.; Valiere, S.; Bouchez, O.; Hunault, G.; Simoneau, P.; Jacques, M.-A. Emergence shapes the structure of the seed microbiota. Appl. Environ. Microbiol. 2015, 81, 1257. [Google Scholar] [CrossRef] [Green Version]
- Klaedtke, S.; Jacques, M.A.; Raggi, L.; Preveaux, A.; Bonneau, S.; Negri, V.; Chable, V.; Barret, M. Terroir is a key driver of seed-associated microbial assemblages. Environ. Microbiol. 2016, 18, 1792–1804. [Google Scholar] [CrossRef] [Green Version]
- Nelson, E.B. The seed microbiome: Origins, interactions, and impacts. Plant Soil. 2018, 422, 7–34. [Google Scholar] [CrossRef]
- Ji, S.H.; Kim, J.S.; Lee, C.H.; Seo, H.S.; Chun, S.C.; Oh, J.; Choi, E.-H.; Park, G. Enhancement of vitality and activity of a plant growth-promoting bacteria (PGPB) by atmospheric pressure non-thermal plasma. Sci. Rep. 2019, 9, 1044. [Google Scholar] [CrossRef] [PubMed]
- Mitra, A.; Li, Y.F.; Klämpfl, T.G.; Shimizu, T.; Jeon, J.; Morfill, G.E.; Zimmermann, J.L. Inactivation of surface-borne microorganisms and increased germination of seed specimen by cold atmospheric plasma. Food Bioprocess Technol. 2014, 7, 645–653. [Google Scholar] [CrossRef] [Green Version]
- Butscher, D.; Van, L.H.; Waskow, A.; Rudolf von Rohr, P.; Schuppler, M. Plasma inactivation of microorganisms on sprout seeds in a dielectric barrier discharge. Int. J. Food. Microbiol. 2016, 238, 222–232. [Google Scholar] [CrossRef]
- Selcuk, M.; Oksuz, L.; Basaran, P. Decontamination of grains and legumes infected with Aspergillus spp. and Penicillum spp. by cold plasma treatment. Bioresour. Technol. 2008, 99, 5104–5109. [Google Scholar] [CrossRef]
- Hong, Y.F.; Kang, J.G.; Lee, H.Y.; Uhm, H.S.; Moon, E.; Park, Y.H. Sterilization effect of atmospheric plasma on Escherichia coli and Bacillus subtilis endospores. Lett. Appl. Microbiol. 2009, 48, 33–37. [Google Scholar] [CrossRef]
- Reineke, K.; Langer, K.; Hertwig, C.; Ehlbeck, J.; Schluter, O. The impact of different process gas compositions on the inactivation effect of an atmospheric pressure plasma jet on Bacillus spores. Innov. Food Sci. Emerg. 2015, 30, 112–118. [Google Scholar] [CrossRef]
- Liao, X.; Muhammad, A.I.; Chen, S.; Hu, Y.; Ye, X.; Liu, D.; Ding, T. Bacterial spore inactivation induced by cold plasma. Crit. Rev. Food Sci. Nutr. 2019, 59, 2562–2572. [Google Scholar] [CrossRef]
- Schnabel, U.; Niquet, R.; Krohmann, U.; Winter, J.; Schluter, O.; Weltmann, K.D.; Ehlbeck, J. Decontamination of microbiologically contaminated specimen by direct and indirect plasma treatment. Plasma Process Polym. 2012, 9, 569–575. [Google Scholar] [CrossRef]
- Butscher, D.; Zimmermann, D.; Schuppler, M.; Rudolf von Rohr, P. Plasma inactivation of bacterial endospores on wheat grains and polymeric model substrates in a dielectric barrier discharge. Food Control 2016, 60, 636–645. [Google Scholar] [CrossRef]
- Han, L.; Patil, S.; Boehm, D.; Milosavljevic, V.; Cullen, P.J.; Bourke, P. Mechanisms of inactivation by high-voltage atmospheric cold plasma differ for Escherichia coli and Staphylococcus aureus. Appl. Environ. Microbiol. 2016, 82, 450–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Q.S.; Huang, C.; Hsieh, F.H.; Huff, H.; Duan, Y. Bacterial inactivation using a low-temperature atmospheric plasma brush sustained with argon gas. J. Biomed. Mater. Res. 2007, 80B, 211–219. [Google Scholar] [CrossRef]
- Boudam, M.K.; Moisan, M.; Saoudi, B.; Popovici, C.; Gherardi, N.; Massines, F. Bacterial spore inactivation by atmospheric-pressure plasmas in the presence or absence of UV photons as obtained with the same gas mixture. J. Phys. D Appl. Phys. 2006, 39, 3494–3507. [Google Scholar] [CrossRef]
- Moisan, M.; Barbeau, J.; Moreau, S.; Pelletier, J.; Tabrizian, M.; Yahia, L. Low-temperature sterilization using gas plasmas: A review of the experiments and an analysis of the inactivation mechanisms. Int. J. Pharm. 2001, 226, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Moreau, S.; Moisan, M.; Tabrizian, M.; Barbeau, J.; Pelletier, J.; Ricard, A.; Yahia, L. Using the flowing afterglow of a plasma to inactivate Bacillus subtilis spores: Influence of the operating conditions. J. Appl. Phys. 2000, 88, 1166–1174. [Google Scholar] [CrossRef]
- Ritter, A.C.; Santi, L.; Vannini, L.; Beys-da-Silva, W.O.; Gozzi, G.; Yates, J., 3rd; Ragni, L.; Brandelli, A. Comparative proteomic analysis of foodborne Salmonella Enteritidis SE86 subjected to cold plasma treatment. Food Microbiol. 2018, 76, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Yau, K.P.S.; Murphy, A.B.; Zhong, L.; Mai-Prochnow, A. Cold plasma effect on the proteome of Pseudomonas aeruginosa-Role for bacterioferritin. PLoS ONE 2018, 13, e0206530. [Google Scholar] [CrossRef]
- Sharma, A.; Collins, G.; Pruden, A. Differential gene expression in Escherichia coli following exposure to nonthermal atmospheric pressure plasma. J. Appl. Microbiol. 2009, 107, 1440–1449. [Google Scholar] [CrossRef]
- Roth, S.; Feichtinger, J.; Hertel, C. Response of Deinococcus radiodurans to low-pressure low-temperature plasma sterilization processes. J. Appl. Microbiol. 2010, 109, 1521–1530. [Google Scholar] [CrossRef]
- Winter, T.; Bernhardt, J.Ã.; Winter, J.Ã.; Mader, U.; Schluter, R.; Weltmann, K.D.; Hecker, M.; Kusch, H. Common versus noble Bacillus subtilis differentially responds to air and argon gas plasma. Proteomics 2013, 13, 2608–2621. [Google Scholar] [CrossRef]
- Winter, T.; Winter, J.Ã.; Polak, M.; Kusch, K.; Mader, U.; Sietmann, R.; Ehlbeck, J.; van Hijum, S.; Weltmann, K.D.; Hecker, M.; et al. Characterization of the global impact of low temperature gas plasma on vegetative microorganisms. Proteomics 2011, 11, 3518–3530. [Google Scholar] [CrossRef] [PubMed]
- Krewing, M.; Jarzina, F.; Dirks, T.; Schubert, B.; Benedikt, J.; Lackmann, J.W.; Bandow, J.E. Plasma-sensitive Escherichia coli mutants reveal plasma resistance mechanisms. J. R. Soc. Interface 2019, 16, 20180846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mols, M.; Mastwijk, H.; Nierop Groot, M.; Abee, T. Physiological and transcriptional response of Bacillus cereus treated with low-temperature nitrogen gas plasma. J. Appl. Microbiol. 2013, 115, 689–702. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Koberl, M.; Rybakova, D.; Muller, H.; Grosch, R.; Smalla, K. Plant microbial diversity is suggested as the key to future biocontrol and health trends. FEMS Microbiol. Ecol. 2017, 93, fix050. [Google Scholar] [CrossRef]
- Sessitsch, A.; Mitter, B. 21st century agriculture: Integration of plant microbiomes for improved crop production and food security. Microb. Biotechnol. 2015, 8, 32–33. [Google Scholar] [CrossRef]
- Ferguson, B.J.; Mens, C.; Hastwell, A.H.; Zhang, M.; Su, H.; Jones, C.H.; Chu, X.; Gresshoff, P.M. Legume nodulation: The host controls the party. Plant Cell Environ. 2019, 42, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Kawase, M.; Chen, W.; Kawaguchi, K.; Nyasha, M.R.; Sasaki, S.; Hatakeyama, H.; Kaneko, T.; Kanzaki, M. TRPA1 and TRPV1 channels participate in atmospheric-pressure plasma-induced [Ca2+]i response. Sci. Rep. 2020, 10, 9687. [Google Scholar] [CrossRef]
- Himmel, N.J.; Cox, D.N. Transient receptor potential channels: Current perspectives on evolution, structure, function and nomenclature. Proc. R. Soc. B 2020, 287, 20201309. [Google Scholar] [CrossRef]
- Demidchik, V.; Shabala, S.; Isayenkov, S.; Cuin, T.A.; Pottosin, I. Calcium transport across plant membranes: Mechanisms and functions. New Phytologist. 2018, 220, 49–69. [Google Scholar] [CrossRef] [Green Version]
- Sano, N.; Rajjou, L.; North, H.M.; Debeaujon, I.; Marion-Poll, A.; Seo, M. Staying alive: Molecular Aspects of Seed Longevity. Plant Cell Physiol. 2015, 57, 660–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]


| Dormancy Type | Key Determinant | NTP Effects Due to |
|---|---|---|
| Physical, PY | Permeability is limited by the seed coat | Changes in the surface and improved permeability of the seed coat |
| Physiological, PD | Phytohormone balance (low GA/ABA) | Shift in the balance of phytohormones (GA/ABA increase) |
| Morphological, MD Morphophysiological, MPD | Under-developed or immature embryo Under-developed embryo and phytohormones | NTP not effective |
| Combinational: physical and physiological, PY + PD | Germination is limited by the seed coat and inhibited by phytohormones | Combination of the involved factors (both coat and phytohormonal changes) |
| Non dormant seeds, ND | Seeds are germination competent | Negligible effects on germination |
| NTP Induced Change | Plant Species [Reference] | Implication |
|---|---|---|
| Increased number of paramagnetic centers (EPR signal) in seeds | Norway spruce [113], radish [77,78] | Increased production of stable organic radicals indicates the interaction of seed components with ROS (NTP generated NTP or internally produced) |
| Increased ROS amount in dry and in germinating seeds | Wheat [89], A. thaliana [87], Norway spruce [113], soybean [95] | Induced internal RONS production; RONS involved in NTP effects |
| Change in the balance of phytohormones involved in the control of germination | Radish [78] | NTP effects on germination are related to induced shift in GA/ABA |
| Gene expression and expression or activities of proteins (including enzymes) | Mung bean [50], rice [56,80], A. thaliana [87], spinach [147], soybean [95], wheat [98] | Induced changes in the expression or activities of proteins/enzymes involved in mobilisation of resources for germination and antioxidative defense |
| DNA methylation | Rice [80] | NTP induces changes in gene expression through changes in DNA methylation. |
| NTP Induced Change in | Plant Species [Reference] | Implication: |
|---|---|---|
| DNA methylation | Soybean [96] | Impact on gene expression through DNA methylation |
| Gene and protein expression, including proteins involved in photosynthesis, stress response, secondary metabolism | Arabidopsis [163,164], bitter melon [165], blue sage [166], industrial hemp [167], maize [168], soybean [94,96], sunflower [58,169,170], tomato [171,172,173], wheat [81,174,175] | Changed expression and amounts of proteins in growing plants |
| Enzyme activities | Arabidopsis [87], artichoke [60], A. fridae [176], green chiretta [69], lemon balm [177], maize [178,179,180], pea [18], pepper [181], rice [182], soybean [94,96], sweet basil [183], tomato [172,184], wheat [89,98,185,186] | Changes in plant metabolism and antioxidant defense |
| Amount of phytohormones in plants | Maize [108], pea [71], tomato [173] | |
| Content of photosynthetic pigments | carrot [91], wheat [63,98,185,187,188], Norway spruce [70], maize [108,180], rice [93,178], soybean [189], spinach [147], tomato [172] | Improved growth due to up-regulated photosynthesis. |
| Activity or efficiency of photosynthesis | common buckwheat [109], maize [179], rice [182], pea [121], purple coneflower [190], soybean [189], sunflower [81], wheat [191] | |
| Secondary metabolism | coriander [92], brown rice [56,79], buckwheat [109], industrial hemp [48], maize [180,192], Norway spruce [70], purple coneflower [190], rapeseed [118], red clover [54,148,193], soybean [94], spinach [25], wheat [188,192] | Increased content of secondary metabolites is important for establishment of seedlings, plant fitness, stress resistance, communication with microorganisms. |
| Communication with microorganisms | Arabidopsis [194], sunflower [170], soybean [94], red clover [148] | Changed interactions with pathogens and beneficial microorganisms |
| Plant growth for the entire vegetation period and production yield | Arabidopsis [85], black mulberry [47], common buckwheat [109], garlic [195], industrial hemp [48], maize [179,196,197], Norway spruce [113], purple coneflower [190], peanut [198], red clover [54], rhododendron [47], tomato [40,171,199], wheat [63,191,192,197] | Improved plant growth for longer period of time. Persistent effects show the potential of NTP treatment for Plasma in Agriculture |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mildaziene, V.; Ivankov, A.; Sera, B.; Baniulis, D. Biochemical and Physiological Plant Processes Affected by Seed Treatment with Non-Thermal Plasma. Plants 2022, 11, 856. https://doi.org/10.3390/plants11070856
Mildaziene V, Ivankov A, Sera B, Baniulis D. Biochemical and Physiological Plant Processes Affected by Seed Treatment with Non-Thermal Plasma. Plants. 2022; 11(7):856. https://doi.org/10.3390/plants11070856
Chicago/Turabian StyleMildaziene, Vida, Anatolii Ivankov, Bozena Sera, and Danas Baniulis. 2022. "Biochemical and Physiological Plant Processes Affected by Seed Treatment with Non-Thermal Plasma" Plants 11, no. 7: 856. https://doi.org/10.3390/plants11070856
APA StyleMildaziene, V., Ivankov, A., Sera, B., & Baniulis, D. (2022). Biochemical and Physiological Plant Processes Affected by Seed Treatment with Non-Thermal Plasma. Plants, 11(7), 856. https://doi.org/10.3390/plants11070856