
Quinoa Response to Application of Phosphogypsum and Plant Growth-Promoting Rhizobacteria under Water Stress Associated with Salt-Affected Soil


 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Exchangeable Na % in the Soil and Soil Enzymes’ Activity
2.2. Concentration of Na+, K+, and K+/Na+ in the Leaves
2.3. Photosynthetic Pigments (Chlorophyll a, Chlorophyll b, and Total Carotenoids)
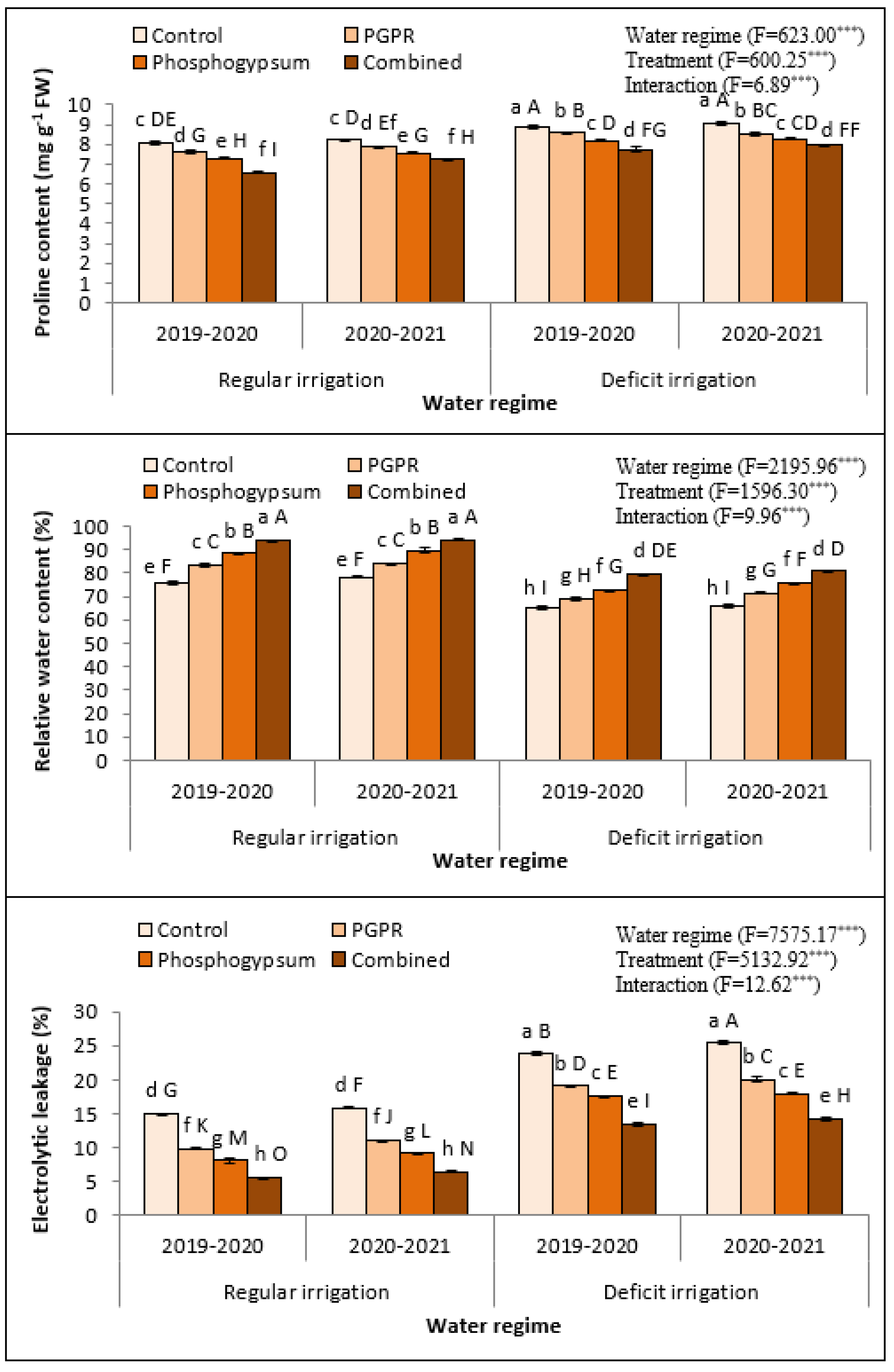
2.4. Proline Content, Relative Water Content, and Electrolytic Leakage
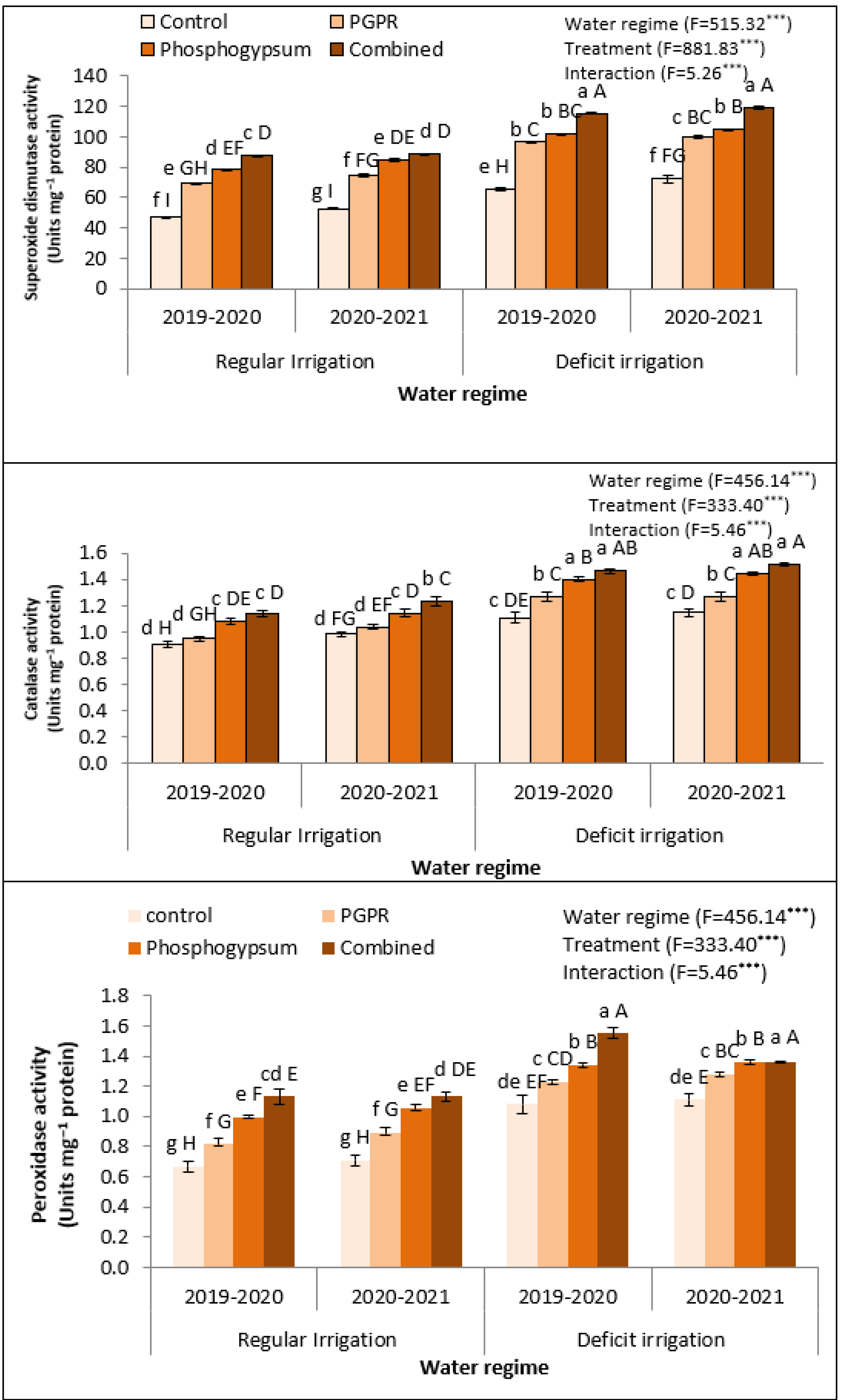
2.5. Phosphogypsum and PGPR Improved the Antioxidant-Related Enzymatic Activity in Deficiently Irrigated Quinoa Plant under Saline Soil
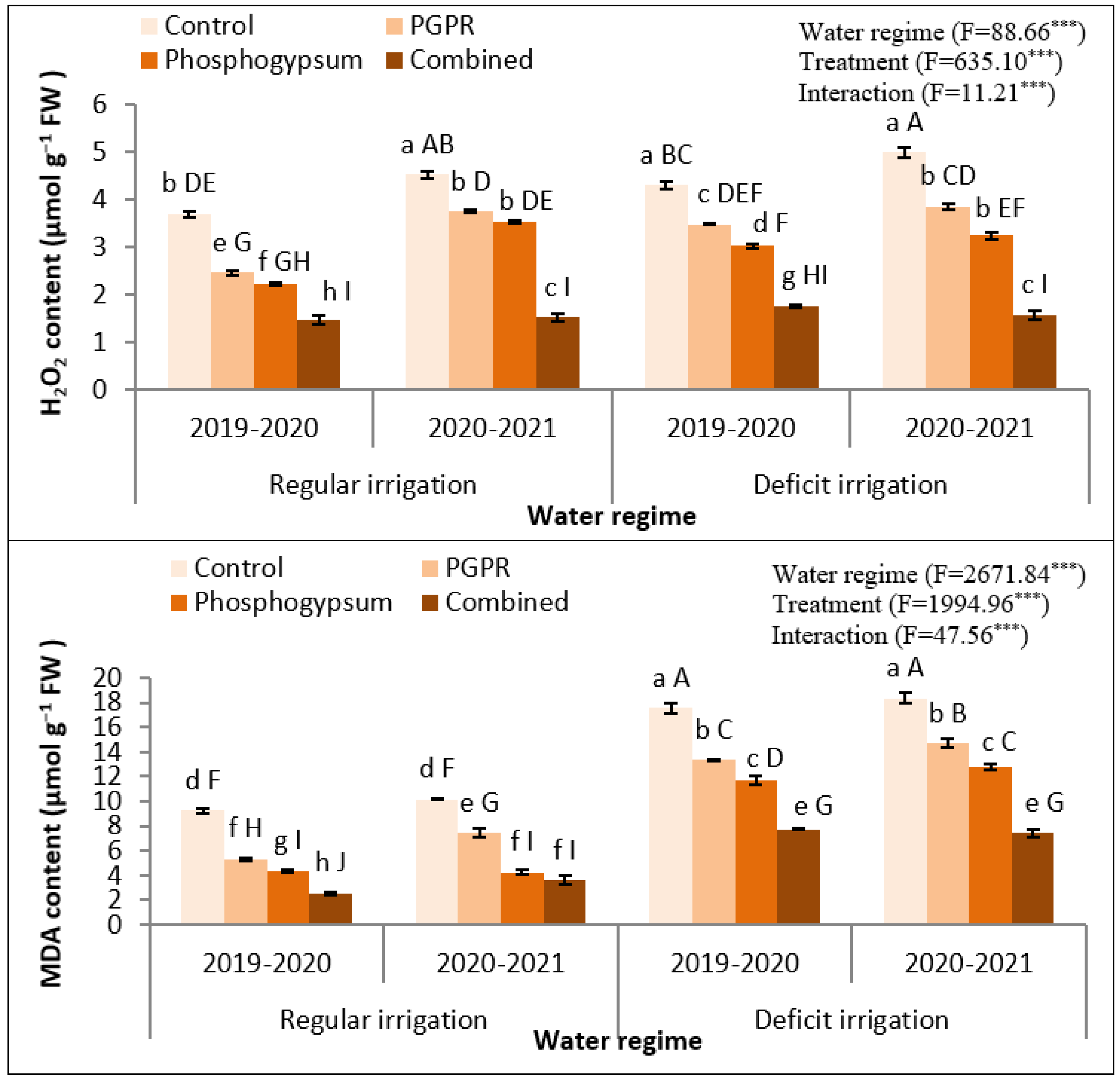
2.6. Phosphogypsum and PGPR Application Deflate-Stress Biochemical Indicators in Stressed Quinoa Plants
2.7. Phosphogypsum and PGPR Application Enhanced the Yield Traits in Stressed Quinoa Plants
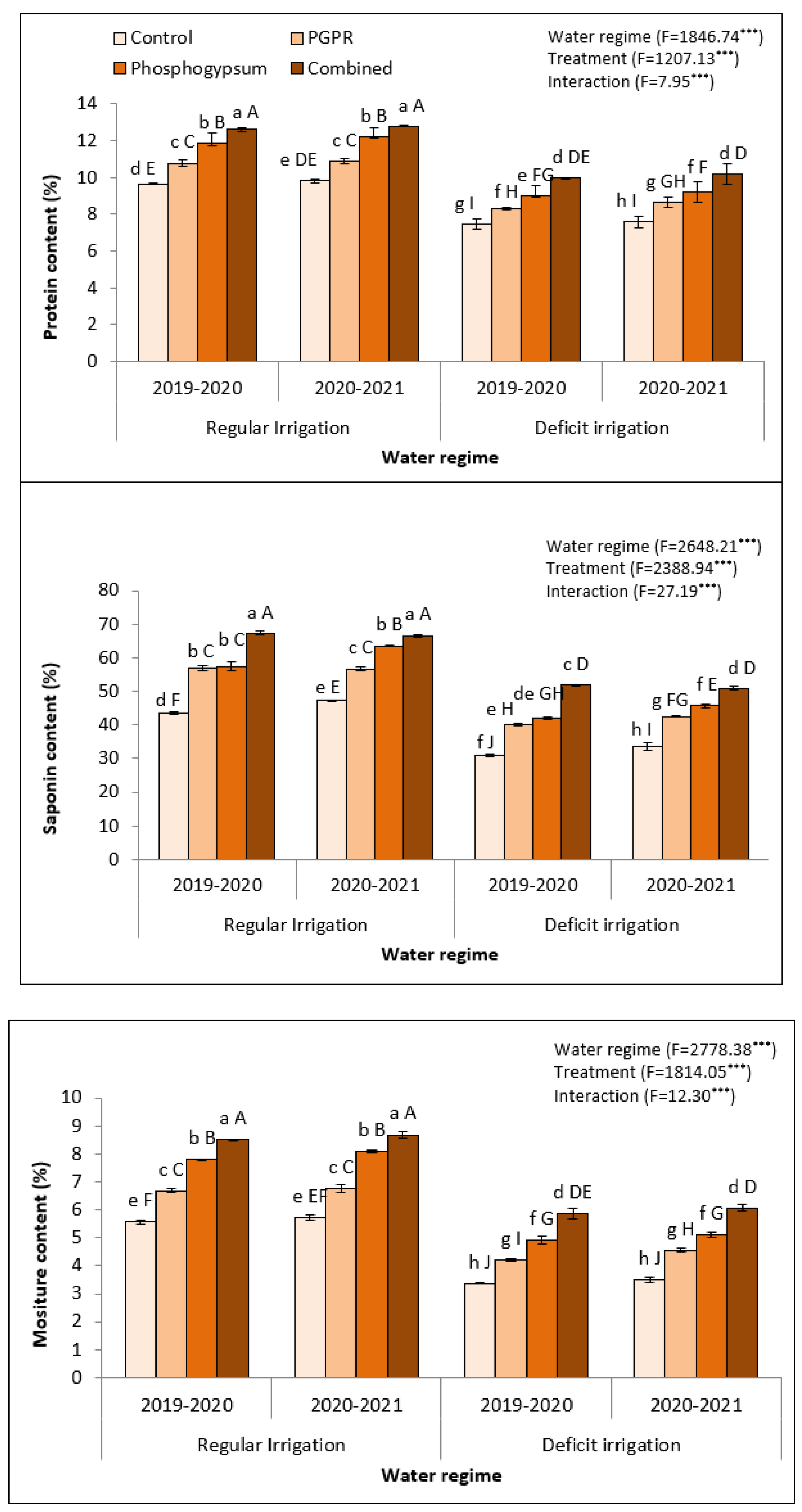
2.8. Phosphogypsum and PGPR Application Enhanced the Seed Quality in Stressed Quinoa Plants
3. Discussion
3.1. Effect of Soil Treatments on Soil Exchangeable Na Percentage, Soil Enzymes’ Activity (Urease and Dehydrogenase), in Addition to Na+ and K+ Ions in Leaves under Abiotic Stress
3.2. Effect of Soil Treatments on Photosynthetic Pigment Contents and Indicators of Plant Responses to Stresses under Abiotic Stress
3.3. Effect of Soil Treatments on Antioxidant Enzymes’ Activity and Oxidative Stress under Abiotic Stress
3.4. Effect of Soil Amendments on Quinoa Yield Related Traits, Productivity, and Seed Quality under Abiotic Stress
4. Materials and Methods
4.1. Phosphogypsum (PG) Characterization
4.1.1. Microorganisms and Culture Conditions
4.1.2. Study Site Description and Plant Material
4.1.3. Experimental Design and Treatments
4.1.4. Soil Measurements
Exchangeable Sodium Percentage (ESP) in Soil
Soil Enzymes’ Activity
4.2. Physiological Attributes
4.2.1. Measurements of Na+ and K+ Ions in Leaves
4.2.2. Photosynthetic Pigment Contents
4.2.3. Indicators of Plant Responses to Stresses
Leaf Water Relations
Proline Content
Electrolyte Leakage
4.3. Antioxidant Enzyme Activity
4.3.1. Superoxide Dismutase Activity
4.3.2. Catalase Activity
4.3.3. Peroxidase Activity
4.4. Oxidative Stress Markers
4.4.1. Hydrogen Peroxide (H2O2)
4.4.2. Lipid Peroxidation (MDA)
4.5. Plant Growth and Yield Parameters
4.6. Phytochemical Screening of the Extracts
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adolf, V.I.; Shabala, S.; Andersen, M.N.; Razzaghi, F.; Jacobsen, S.-E. Varietal differences of quinoa’s tolerance to saline conditions. Plant Soil 2012, 357, 117–129. [Google Scholar] [CrossRef]
- Jacobsen, S.E.; Mujica, A.; Jensen, C.R. The resistance of quinoa (Chenopodium quinoa Willd.) to adverse abiotic factors. Food Rev. Int. 2003, 19, 99–109. [Google Scholar]
- Bazile, D.; Jacobsen, S.-E.; Verniau, A. The global expansion of quinoa: Trends and limits. Front. Plant Sci. 2016, 7, 622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazile, D.; Bertero, H.D.; Nieto, C. State of the Art Report on Quinoa around the World in 2013; FAO: Rome, Italy, 2015; Available online: https://agritrop.cirad.fr/575489/1/document_575489.pdf (accessed on 1 August 2021).
- Jaikishun, S.; Li, W.; Yang, Z.; Song, S. Quinoa: In perspective of global challenges. Agronomy 2019, 9, 176. [Google Scholar] [CrossRef] [Green Version]
- Maradini-Filho, A. Quinoa: Nutritional aspects. J. Nutraceuticals Food Sci. 2017, 2, 1–5. [Google Scholar]
- Hinojosa, L.; González, J.A.; Barrios-Masias, F.H.; Fuentes, F.; Murphy, K.M. Quinoa abiotic stress responses: A review. Plants 2018, 7, 106. [Google Scholar] [CrossRef] [Green Version]
- FAO. Quinoa: An Ancient Crop to Contribute to World Food Security; Food and Agriculture Organization of the United Nations: Rome, Italy, 2011. [Google Scholar]
- FAO. The Future of Food and Agriculture: Trends and Challenges; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020. [Google Scholar]
- Richards, L.A. Diagnosis and Improvement of Saline and Alkali Soils; Agriculture Handbook No. 60; US Department of Agriculture: Washington, DC, USA, 1954.
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Flowers, T.J.; Galal, H.K.; Bromham, L. Evolution of halophytes: Multiple origins of salt tolerance in land plants. Funct. Plant Biol. 2010, 37, 604–612. [Google Scholar] [CrossRef]
- Abd El-Moneim, D.; ELsarag, E.I.S.; Aloufi, S.; El-Azraq, A.M.; ALshamrani, S.M.; Safhi, F.A.A.; Ibrahim, A.A. Quinoa (Chenopodium quinoa Willd.): Genetic diversity according to ISSR and SCoT markers, relative gene expression, and morpho-physiological variation under salinity stress. Plants 2021, 10, 2802. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A.; Läuchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar]
- Killi, D.; Haworth, M. Diffusive and metabolic constraints to photosynthesis in quinoa during drought and salt stress. Plants 2017, 6, 49. [Google Scholar] [CrossRef] [Green Version]
- Saddiq, M.S.; Wang, X.; Iqbal, S.; Hafeez, M.B.; Khan, S.; Raza, A.; Iqbal, J.; Maqbool, M.M.; Fiaz, S.; Qazi, M.A.; et al. Effect of water stress on grain yield and physiological characters of quinoa genotypes. Agronomy 2021, 11, 1934. [Google Scholar] [CrossRef]
- Valdivia-Cea, W.; Bustamante, L.; Jara, J.; Fischer, S.; Holzapfel, E.; Wilckens, R. Effect of soil water availability on physio-logical parameters, yield, and seed quality in four quinoa genotypes (Chenopodium quinoa Willd.). Agronomy 2021, 11, 1012. [Google Scholar]
- Alvar-Beltrán, J.; Dao, A.; Dalla, M.A.; Saturnin, C.; Casini, P.; Sanou, J.; Orlandini, S. Effect of drought, nitrogen fertilization, temperature and photoperiodicity on quinoa plant growth and development in the Sahel. Agronomy 2019, 9, 607. [Google Scholar]
- Zhu, J.-K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Leal, L.D.S.G.; Pessoa, L.G.M.; de Oliveira, J.P.; Santos, N.A.; Silva, L.F.D.S.; Júnior, G.B.; Freire, M.B.G.D.S.; de Souza, E.S. Do applications of soil conditioner mixtures improve the salt extraction ability of Atriplex nummularia at early growth stage? Int. J. Phytoremediat. 2020, 22, 482–489. [Google Scholar]
- Saadaoui, E.; Ghazel, N.; Ben Romdhane, C.; Massoudi, N. Phosphogypsum: Potential uses and problems—A review. Int. J. Environ. Stud. 2017, 74, 558–567. [Google Scholar]
- Mazzilli, B.; Palmiro, V.; Saueia, C.; Nisti, M. Radiochemical characterization of Brazilian phosphogypsum. J. Environ. Radioact. 2000, 49, 113–122. [Google Scholar] [CrossRef]
- Olivé, F.D.; Bosch-Serra, À.D.; Yagüe, M.R.; Poch, R.M.; Boixadera, J. Long term application of dairy cattle manure and pig slurry to winter cereals improves soil quality. Nutr. Cycl. Agroecosyst. 2016, 104, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Mesić, M.; Brezinščak, L.; Zgorelec, Ž.; Perčin, A.; Šestak, I.; Bilandžija, D.; Trdenić, M.; Lisac, H. The application of phosphogypsum in agriculture. Agric. Conspec. Sci. 2016, 81, 7–13. [Google Scholar]
- Al-Enazy, A.R.; Al-Oud, S.S.; Al-Barakah, F.N.; Usman, A.R. Role of microbial inoculation and industrial by-product phos-phogypsum in growth and nutrient uptake of maize (Zea mays L.) grown in calcareous soil. J. Sci. Food Agric. 2017, 97, 3665–3674. [Google Scholar] [PubMed]
- Lee, C.H.; Ha, B.Y.; Lee, Y.B.; Kim, P.J. Effect of alkalized phosphogypsum on soil chemical and biological properties. Commun. Soil Sci. Plant Anal. 2009, 40, 2072–2086. [Google Scholar] [CrossRef]
- Mahmoud, E.; Ghoneim, A.; El Baroudy, A.; Abd El-Kader, N.; Aldhumri, S.A.; Othman, S.; El Khamisy, R. Effects of phos-phogypsum and water treatment residual application on key chemical and biological properties of clay soil and maize yield. Soil Use Manag. 2020, 37, 494–503. [Google Scholar]
- Aslam, M.U.; Raza, M.A.S.; Saleem, M.F.; Waqas, M.; Iqbal, R.; Ahmad, S.; Haider, I. Improving strategic growth stage-based drought tolerance in quinoa by rhizobacterial inoculation. Commun. Soil Sci. Plant Anal. 2020, 51, 853–868. [Google Scholar] [CrossRef]
- Fghire, R.; Anaya, F.; Ali, O.I.; Benlhabib, O.; Ragab, R.; Wahbi, S. Physiological and photosynthetic response of quinoa to drought stress. Chil. J. Agric. Res. 2015, 75, 174–183. [Google Scholar] [CrossRef] [Green Version]
- Ilangumaran, G.; Smith, D.L. Plant growth promoting rhizobacteria in amelioration of salinity stress: A systems biology perspective. Front. Plant Sci. 2017, 8, 1768. [Google Scholar] [CrossRef]
- Ayuso-Calles, M.; Flores-Félix, J.D.; Rivas, R. Overview of the role of rhizobacteria in plant salt stress tolerance. Agronomy 2021, 11, 1759. [Google Scholar] [CrossRef]
- Sushanto, G.; Rout, G.K.; Gitishree, D.; Spiros, P.; Han-Seung, S.; Jayanta, K.P.; Yang, A.Z.; Akhtar, S.S.; Iqbal, S.; Amjad, M.; et al. Enhancing salt tolerance in quinoa by halotolerant bacterial inoculation. Funct. Plant Biol. 2016, 43, 632–642. [Google Scholar]
- Martínez, E.A.; Veas, E.; Jorquera, C.; Martín, R.S.; Jara, P. Re-introduction of quínoa into arid Chile: Cultivation of two lowland races under extremely low irrigation. J. Agron. Crop Sci. 2009, 195, 1–10. [Google Scholar] [CrossRef]
- Hafez, E.E.-D.M.; El Hassan, W.H.A.; Gaafar, I.A.; Seleiman, M.F. Effect of gypsum application and irrigation intervals on clay saline-sodic soil characterization, rice water use efficiency, growth, and yield. J. Agric. Sci. 2015, 7, 208. [Google Scholar] [CrossRef] [Green Version]
- Kamara, M.M.; Rehan, M.; Ibrahim, K.M.; Alsohim, A.S.; Elsharkawy, M.M.; Kheir, A.M.S.; Hafez, E.M.; El-Esawi, M.A. Genetic diversity and combining ability of white maize inbred lines under different plant densities. Plants 2020, 9, 1140. [Google Scholar] [CrossRef]
- Nehela, Y.; Mazrou, Y.S.A.; Alshaal, T.; Rady, A.M.S.; El-Sherif, A.M.A.; Omara, A.E.-D.; El-Monem, A.M.A.; Hafez, E.M. The integrated amendment of sodic-saline soils using biochar and plant growth-promoting rhizobacteria enhances maize (Zea mays L.) resilience to water salinity. Plants 2021, 10, 1960. [Google Scholar] [PubMed]
- Garcia, C.; Hernandez, T.; Costa, F. Potential use of dehydrogenase activity as an index of microbial activity in degraded soils. Commun. Soil Sci. Plant Anal. 1997, 28, 123–134. [Google Scholar] [CrossRef]
- Kheir, A.M.S.; Abouelsoud, H.M.; Hafez, E.M.; Ali, O.A.M. Integrated effect of nano-Zn, nano-Si, and drainage using crop straw–filled ditches on saline sodic soil properties and rice productivity. Arab. J. Geosci. 2019, 12, 471. [Google Scholar]
- Hafez, E.M.; Alsohim, A.S.; Farig, M.; Omara, A.E.-D.; Rashwan, E.; Kamara, M.M. Synergistic effect of biochar and plant growth promoting rhizobacteria on alleviation of water deficit in rice plants under salt-affected soil. Agronomy 2019, 9, 847. [Google Scholar] [CrossRef] [Green Version]
- Osman, H.S.; Rady, A.M.S.; Awadalla, A.; Omara, A.E.-D.; Hafez, E.M. Improving the antioxidants system, growth, and sugar beet quality subjected to long-term osmotic stress by phosphate solubilizing bacteria and compost tea. Int. J. Plant Prod. 2021, 1–17. [Google Scholar] [CrossRef]
- Abdelrasheed, K.G.; Mazrou, Y.; Omara, A.E.-D.; Osman, H.S.; Nehela, Y.; Hafez, E.M.; Rady, A.M.S.; El-Moneim, D.A.; Alowaiesh, B.F.; Gowayed, S.M. Soil amendment using biochar and application of K-humate enhance the growth, productivity, and nutritional value of onion (Allium cepa L.) under deficit irrigation conditions. Plants 2021, 10, 2598. [Google Scholar] [CrossRef]
- Osman, H.; Gowayed, S.; Elbagory, M.; Omara, A.; El-Monem, A.; El-Razek, U.A.; Hafez, E. Interactive impacts of beneficial microbes and Si-Zn nanocomposite on growth and productivity of soybean subjected to water deficit under salt-affected soil conditions. Plants 2021, 10, 1396. [Google Scholar] [CrossRef]
- Ding, Z.; Kheir, A.M.S.; Ali, O.A.M.; Hafez, E.M.; ElShamey, E.A.; Zhou, Z.; Wang, B.; Lin, X.; Ge, Y.; Fahmy, A.E.; et al. A vermicompost and deep tillage system to improve saline-sodic soil quality and wheat productivity. J. Environ. Manag. 2021, 277, 111388. [Google Scholar] [CrossRef]
- Hafez, E.; Farig, M. Efficacy of salicylic acid as a cofactor for ameliorating effects of water stress and enhancing wheat yield and water use efficiency in saline soil. Int. J. Plant Prod. 2019, 13, 163–176. [Google Scholar] [CrossRef]
- Hafez, E.E.-D.M. Influence of salicylic acid on ion distribution, enzymatic activity and some agromorphological characteristics of wheat under salt-affected soil. Egypt. J. Agron. 2016, 38, 455–469. [Google Scholar] [CrossRef] [Green Version]
- Seleiman, M.F.; Refay, Y.; Al-Suhaibani, N.; Al-Ashkar, I.; El-Hendawy, S.; Hafez, E.M.; Suhaibani, A.; Ashkar, A.; Hendawy, E. Integrative effects of rice-straw biochar and silicon on oil and seed quality, yield and physiological traits of Helianthus annuus L. grown under water deficit stress. Agronomy 2019, 9, 637. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharjee, S. ROS and oxidative stress: Origin and implication. In Reactive Oxygen Species in Plant Biology; Springer: New Delhi, India, 2019; pp. 1–31. [Google Scholar]
- Abbas, G.; Amjad, M.; Saqib, M.; Murtaza, B.; Naeem, M.A.; Shabbir, A.; Murtaza, G. Soil sodicity is more detrimental than salinity for quinoa (Chenopodium quinoa Willd.): A multivariate comparison of physiological, biochemical and nutritional quality attributes. J. Agron. Crop Sci. 2021, 207, 59–73. [Google Scholar] [CrossRef]
- Nafees, M.; Fahad, S.; Shah, A.N.; Bukhari, M.A.; Maryam, I.; Ahmad, S.; Hussain, S. Reactive oxygen species signaling in plants. In Plant Abiotic Stress Tolerance; Springer: Cham, Switzerland, 2019; pp. 259–272. [Google Scholar]
- Chung, W.-H. Unraveling new functions of superoxide dismutase using yeast model system: Beyond its conventional role in superoxide radical scavenging. J. Microbiol. 2017, 55, 409–416. [Google Scholar] [CrossRef]
- Ahmad, R.; Hussain, S.; Anjum, M.A.; Khalid, M.F.; Saqib, M.; Zakir, I.; Hassan, A.; Fahad, S.; Ahmad, S. Oxidative stress and antioxidant defense mechanisms in plants under salt stress. In Plant Abiotic Stress Tolerance; Springer: Cham, Switzerland, 2019; pp. 191–205. [Google Scholar] [CrossRef]
- Alam, M.Z.; Carpenter-Boggs, L.; Hoque, A.; Ahammed, G.J. Effect of soil amendments on antioxidant activity and photo-synthetic pigments in pea crops grown in arsenic contaminated soil. Heliyon 2020, 6, e05475. [Google Scholar]
- Hasanuzzaman, M.; Parvin, K.; Bardhan, K.; Nahar, K.; Anee, T.I.; Masud, A.A.C.; Fotopoulos, V. Biostimulants for the reg-ulation of reactive oxygen species metabolism in plants under abiotic stress. Cells 2021, 10, 2537. [Google Scholar]
- Hafez, E.M.; Gowayed, S.M.; Nehela, Y.; Sakran, R.M.; Rady, A.M.S.; Awadalla, A.; Omara, A.E.-D.; Alowaiesh, B.F. incorporated biochar-based soil amendment and exogenous glycine betaine foliar application ameliorate rice (Oryza sativa L.) tolerance and resilience to osmotic stress. Plants 2021, 10, 1930. [Google Scholar]
- Hafez, E.M.; Omara, A.E.D.; Alhumaydhi, F.A.; El-Esawi, M.A. Minimizing hazard impacts of soil salinity and water stress on wheat plants by soil application of vermicompost and biochar. Physiol. Plant. 2020, 172, 587–602. [Google Scholar] [CrossRef]
- Hafez, E.; Osman, H.; El-Razek, U.; Elbagory, M.; Omara, A.; Eid, M.; Gowayed, S. Foliar-applied potassium silicate coupled with plant growth-promoting rhizobacteria improves growth, physiology, nutrient uptake and productivity of faba bean (Vicia faba L.) irrigated with saline water in salt-affected soil. Plants 2021, 10, 894. [Google Scholar] [CrossRef]
- Yan, J.M.; Smith, M.D.; Glick, B.R.; Liang, Y. Effects of ACC deaminase containing rhizobacteria on plant growth and expression of Toc GTPases in tomato (Solanum lycopersicum) under salt stress. Botany 2014, 92, 775–781. [Google Scholar]
- Sokol, N.W.; Slessarev, E.; Marschmann, G.L.; Nicolas, A.; Blazewicz, S.J.; Brodie, E.L.; Firestone, M.K.; Foley, M.M.; Hestrin, R.; Hungate, B.A.; et al. Life and death in the soil microbiome: How ecological processes influence biogeochemistry. Nat. Rev. Microbiol. 2022, 1–16. [Google Scholar] [CrossRef]
- King, E.O.; Ward, M.K.; Raney, D.E. Two simple media for the determination of pyocianine and fluorescein. J. Lab. Clin. Med. 1954, 44, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Jensen, H.L. Notes on the biology of azotobacter. Proc. Soc. Appl. Bacteriol. 1951, 14, 89–94. [Google Scholar] [CrossRef]
- Arshad, M.; Lowery, B.; Grossman, B. Physical tests for monitoring soil quality. In Methods for Assessing SoilQuality; Doran, J.W., Jones, A.J., Eds.; Special Publication 49; Soil Science Society of America: Madison, WI, USA, 1997; pp. 123–141. [Google Scholar]
- Öhlinger, R. Dehydrogenase activity with the substrate TTC. In Methods in Soil Biology; Schinner, F., Öhlinger, R., Kandeler, E., Margesin, R., Eds.; Springer: Berlin/Heidelberg, Germany, 1996. [Google Scholar]
- Alef, K.; Nannipieri, P. Urease activity. In Methods in Applied Soil Microbiology and Biochemistry; Alef, K., Nannipieri, P., Eds.; Academic Press: London, UK, 1998; pp. 316–320. [Google Scholar]
- Jackson, M.L.A. Simplified assay for milled rice analysis. J. Cereal Sci. 1971, 16, 334–340. [Google Scholar]
- Anaraki, Z.; Shariati, M.; Hosseini Tafreshi, S. Transient silencing of phytoene desaturase reveals critical roles on plant response to salinity stress. Acta Physiol. Plant 2017, 39, 161. [Google Scholar]
- Weatherley, P.E. Studies in the water relations of the cotton plant. The field measurement of water deficits in leaves. New Phytol. 1950, 49, 81–97. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Bajji, M.; Kinet, J.-M.; Lutts, S. The use of the electrolyte leakage method for assessing cell membrane stability as a water stress tolerance test in durum wheat. Plant Growth Regul. 2002, 36, 61–70. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Vetter, J.L.; Steinberg, M.P.; Nelson, A.I. Enzyme assay, quantitative determination of peroxidase in sweet corn. J. Agric. Food Chem. 1958, 6, 39–41. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Pro-tective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar]
- Du, Z.; Bramlage, W.J. Modified thiobarbituric acid assay for measuring lipid oxidation in sugar-rich plant tissue extracts. J. Agric. Food Chem. 1992, 40, 1566–1570. [Google Scholar] [CrossRef]
- AOAC (Association of Official Analytical Chemists). Official Methods of Analysis Chemists, 17th ed.; AOAC: Washington, DC, USA, 2000. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ESP (%) | Urease (mg TPF g−1 Dry Soil d−1) | Dehydrogenase (mg NH4+ g−1 Dry Soil d−1) | |||
|---|---|---|---|---|---|
| Water regime | |||||
| Regular Irrigation_2019–2020 | 14.7 ± 0.37 c | 139.4 ± 4.2 b | 90.9 ± 3.6 b | ||
| Deficit irrigation_2019–2020 | 21.1 ± 0.32 a | 94.1 ± 2.8 d | 66.1 ± 3.3 d | ||
| Regular Irrigation_2020–2021 | 13.6 ± 0.31 d | 150.3 ± 4.2 a | 96.5 ± 3.8 a | ||
| Deficit irrigation_2020–2021 | 19.8 ± 0.36 b | 100.7 ± 2.7 c | 75.1 ± 3.0 c | ||
| Treatment | |||||
| Control | 21.3 ± 0.35 a | 82.3 ± 1.8 d | 41.6 ± 1.2 d | ||
| PGPR | 15.9 ± 0.36 c | 125.4 ± 3.2 b | 90.1 ± 1.5 b | ||
| Phosphogypsum | 18.6 ± 0.41 b | 111.3 ± 2.5 c | 72.9 ± 1.2 c | ||
| Combined | 13.5 ± 0.37 d | 165.6 ± 3.7 a | 123.9 ± 1.9 a | ||
| Interaction | |||||
| Year | Water regime | Treatment | |||
| 2019–2020 | Regular Irrig. | Control | 19.1 ± 0.16 c, D | 93.0 ± 0.14 f, K | 48.8 ± 0.10 g, M |
| PGPR | 13.0 ± 0.12 f, J | 145.0 ± 0.42 b, D | 99.6 ± 0.16 c, F | ||
| Phosphogypsum | 15.9 ± 0.20 e, G | 127.0 ± 0.19 d, G | 79.8 ± 0.21 d, H | ||
| Combined | 10.7 ± 0.12 g, L | 192.8 ± 0.27 a, B | 135.2 ± 0.33 a, B | ||
| Deficit Irrig. | Control | 24.9 ± 0.21 a, A | 62.9 ± 0.21 h, N | 26.5 ± 0.41 h, O | |
| PGPR | 19.4 ± 0.17 c, D | 96.2 ± 0.25 e, J | 72.8 ± 0.27 e, I | ||
| Phosphogypsum | 22.3 ± 0.21 b, C | 86.3 ± 0.44 g, L | 58.7 ± 0.25 f, K | ||
| Combined | 17.7 ± 0.18 d, F | 130.7 ± 0.43 c, F | 106.3 ± 0.28 b, D | ||
| 2020–2021 | Regular Irrig. | Control | 17.6 ± 0.12 d, F | 101.5 ± 0.21 d, H | 52.7 ± 0.02 f, L |
| PGPR | 12.5 ± 0.17 g, K | 160.0 ± 0.26 b, C | 104.4 ± 0.36 c, E | ||
| Phosphogypsum | 14.3 ± 0.04 f, I | 137.9 ± 0.24 c, E | 84.3 ± 0.45 d, G | ||
| Combined | 10.2 ± 0.18 h, M | 201.9 ± 0.58 a, A | 144.5 ± 0.39 a, A | ||
| Deficit Irrig. | Control | 23.5 ± 0.08 a, B | 71.9 ± 0.63 g, M | 38.3 ± 0.46 g, N | |
| PGPR | 18.5 ± 0.04 c, E | 100.3 ± 0.02 e, I | 83.6 ± 0.32 d, G | ||
| Phosphogypsum | 21.8 ± 0.17 b, C | 94.0 ± 0.05 f, K | 69.1 ± 1.00 e, J | ||
| Combined | 15.5 ± 0.12 e, H | 136.8 ± 0.43 c, E | 109.5 ± 0.64 b, C | ||
| F-value (Two-way) | |||||
| Water regime | 6999.15 *** | 79,968.43 *** | 13,608.91 *** | ||
| Treatment | 5815.84 *** | 123,147.72 *** | 81,585.23 *** | ||
| Interaction | 32.53 *** | 1891.50 *** | 305.76 *** | ||
| Na+ (mg g−1 DW) | K+ (mg g−1 DW) | K+/Na+ | |||
|---|---|---|---|---|---|
| Water regime | |||||
| Regular Irrigation_2019–2020 | 3.15 ± 0.04 b | 3.81 ± 0.05 a | 1.24 ± 0.03 a | ||
| Deficit irrigation_2019–2020 | 3.90 ± 0.04 a | 3.00 ± 0.03 c | 0.78 ± 0.02 b | ||
| Regular Irrigation_2020–2021 | 3.23 ± 0.04 b | 3.87 ± 0.05 a | 1.23 ± 0.03 a | ||
| Deficit irrigation_2020–2021 | 3.92 ± 0.04 a | 3.11 ± 0.03 b | 0.81 ± 0.02 b | ||
| Treatment | |||||
| Control | 4.01 ± 0.04 a | 2.97 ± 0.03 d | 0.75 ± 0.02 d | ||
| PGPR | 3.68 ± 0.04 b | 3.32 ± 0.05 c | 0.92 ± 0.02 c | ||
| Phosphogypsum | 3.43 ± 0.04 c | 3.53 ± 0.05 b | 1.06 ± 0.03 b | ||
| Combined | 3.09 ± 0.04 d | 3.97 ± 0.06 a | 1.32 ± 0.03 a | ||
| Interaction | |||||
| Year | Water regime | Treatment | |||
| 2019–2020 | Regular Irrig. | Control | 3.59 ± 0.10 c, DE | 3.18 ± 0.07 e, FG | 0.89 ± 0.04 e, E |
| PGPR | 3.27 ± 0.04 d, FG | 3.69 ± 0.10 c, CD | 1.13 ± 0.04 c, C | ||
| Phosphogypsum | 3.00 ± 0.06 e, HI | 3.97 ± 0.10 b, B | 1.33 ± 0.02 b, B | ||
| Combined | 2.73 ± 0.06 f, J | 4.39 ± 0.02 a, A | 1.61 ± 0.03 a, A | ||
| Deficit Irrig. | Control | 4.36 ± 0.07 a, A | 2.68 ± 0.03 f, J | 0.62 ± 0.01 h, I | |
| PGPR | 4.02 ± 0.11 b, BC | 2.83 ± 0.01 f, IJ | 0.71 ± 0.02 g, GHI | ||
| Phosphogypsum | 3.81 ± 0.06 b, CD | 3.05 ± 0.04 e, GH | 0.80 ± 0.02 f, EFG | ||
| Combined | 3.40 ± 0.08 cd, ED | 3.44 ± 0.04 d, E | 1.01 ± 0.01 d, D | ||
| 2020–2021 | Regular Irrig. | Control | 3.69 ± 0.09 c, D | 3.24 ± 0.03 e, F | 0.88 ± 0.01 d, E |
| PGPR | 3.36 ± 0.06 de, EFG | 3.77 ± 0.12 c, C | 1.12 ± 0.05 c, C | ||
| Phosphogypsum | 3.12 ± 0.07 e, GH | 3.99 ± 0.02 b, B | 1.28 ± 0.03 b, B | ||
| Combined | 2.77 ± 0.11 f, IJ | 4.49 ± 0.09 a, A | 1.62 ± 0.09 a, A | ||
| Deficit Irrig. | Control | 4.39 ± 0.08 a, A | 2.77 ± 0.04 g, J | 0.63 ± 0.02 f, HI | |
| PGPR | 4.08 ± 0.09 b, B | 2.99 ± 0.02 f, HI | 0.73 ± 0.01 ef, FGH | ||
| Phosphogypsum | 3.79 ± 0.09 c, CD | 3.13 ± 0.02 ef, FGH | 0.83 ± 0.02 de, EF | ||
| Combined | 3.43 ± 0.08 d, EF | 3.55 ± 0.04 d, DE | 1.04 ± 0.02 c, CD | ||
| F-value (Two-way) | |||||
| Water regime | 329.31 *** | 710.52 *** | 562.93 *** | ||
| Treatment | 287.38 *** | 596.16 *** | 504.28 *** | ||
| Interaction | 0.42 ns | 12.40 *** | 14.80 *** | ||
| chl_a (mg g−1 FW) | chl_b (mg g−1 FW) | Carotenoids (mg g−1 FW) | |||
|---|---|---|---|---|---|
| Water regime | |||||
| Regular Irrigation_2019–2020 | 0.480 ± 0.06 b | 0.722 ± 0.05 b | 0.913 ± 0.01 b | ||
| Deficit irrigation_2019–2020 | 0.370 ± 0.05 d | 0.620 ± 0.05 d | 0.638 ± 0.01 d | ||
| Regular Irrigation_2020–2021 | 0.494 ± 0.07 a | 0.744 ± 0.07 a | 0.966 ± 0.01 a | ||
| Deficit irrigation_2020–2021 | 0.384 ± 0.06 c | 0.633 ± 0.06 c | 0.684 ± 0.02 c | ||
| Treatment | |||||
| Control | 0.365 ± 0.05 d | 0.615 ± 0.05 d | 0.634 ± 0.02 d | ||
| PGPR | 0.410 ± 0.07 c | 0.659 ± 0.07 c | 0.788 ± 0.02 c | ||
| Phosphogypsum | 0.449 ± 0.07 b | 0.699 ± 0.07 b | 0.843 ± 0.02 b | ||
| Combined | 0.504 ± 0.07 a | 0.746 ± 0.07 a | 0.936 ± 0.01 a | ||
| Interaction | |||||
| Year | Water regime | Treatment | |||
| 2019–2020 | Regular Irrig. | Control | 0.402 ± 0.004 e, H | 0.653 ± 0.004 e, G | 0.763 ± 0.012 c, GH |
| PGPR | 0.469 ± 0.006 c, E | 0.719 ± 0.006 c, DE | 0.913 ± 0.007 b, DE | ||
| Phosphogypsum | 0.495 ± 0.007 b, D | 0.745 ± 0.005 b, C | 0.953 ± 0.051 b, CD | ||
| Combined | 0.554 ± 0.007 a, B | 0.770 ± 0.009 a, B | 1.023 ± 0.015 a, AB | ||
| Deficit Irrig. | Control | 0.317 ± 0.001 h, K | 0.567 ± 0.001 h, J | 0.477 ± 0.005 e, J | |
| PGPR | 0.347 ± 0.004 g, J | 0.597 ± 0.004 g, I | 0.623 ± 0.012 d, I | ||
| Phosphogypsum | 0.382 ± 0.001 f, I | 0.632 ± 0.001 f, H | 0.670 ± 0.026 d, I | ||
| Combined | 0.434 ± 0.004 d, G | 0.684 ± 0.004 d, F | 0.780 ± 0.017 c, GH | ||
| 2020–2021 | Regular Irrig. | Control | 0.413 ± 0.001 e, H | 0.663 ± 0.001 e, G | 0.820 ± 0.017 d, FG |
| PGPR | 0.473 ± 0.002 c, E | 0.723 ± 0.002 c, D | 0.960 ± 0.018 c, CD | ||
| Phosphogypsum | 0.515 ± 0.004 b, C | 0.765 ± 0.004 b, B | 1.003 ± 0.021 b, BC | ||
| Combined | 0.577 ± 0.009 a, A | 0.827 ± 0.009 a, A | 1.080 ± 0.010 a, A | ||
| Deficit Irrig. | Control | 0.327 ± 0.006 g, K | 0.577 ± 0.006 f, J | 0.477 ± 0.012 g, J | |
| PGPR | 0.353 ± 0.004 f, J | 0.596 ± 0.013 f, I | 0.657 ± 0.015 f, I | ||
| Phosphogypsum | 0.405 ± 0.005 e, H | 0.655 ± 0.005 e, G | 0.743 ± 0.015 e, H | ||
| Combined | 0.453 ± 0.007 d, F | 0.703 ± 0.007 d, E | 0.860 ± 0.010 d, EF | ||
| F-value (Two-way) | |||||
| Water regime | 2012.95 *** | 1300.20 *** | 850.41 *** | ||
| Treatment | 1708.64 *** | 1040.72 *** | 508.04 *** | ||
| Interaction | 14.62 *** | 16.48 *** | 5.33 *** | ||
| Plant Height, cm | 1000-Seed Weight (gm) | Seed Yield (t ha−1) | Foliage Yield (t ha−1) | Biological Yield ‡ (t ha−1) | Harvest Index (%) | |||
|---|---|---|---|---|---|---|---|---|
| Water regime | ||||||||
| Regular Irrigation_2019–2020 | 142.4 ± 3.94 b | 3.49 ± 0.11 a | 2.93 ± 0.06 b | 4.43 ± 0.06 b | 7.36 ± 0.11 b | 39.6 ± 0.16 b | ||
| Deficit irrigation_2019–2020 | 83.3 ± 2.84 d | 1.91 ± 0.05 b | 1.98 ± 0.05 d | 3.48 ± 0.05 d | 5.47 ± 0.09 d | 36.0 ± 0.24 d | ||
| Regular Irrigation_2020–2021 | 151.4 ± 4.65 a | 3.44 ± 0.14 a | 3.10 ± 0.06 a | 4.60 ± 0.06 a | 7.71 ± 0.12 a | 40.1 ± 0.16 a | ||
| Deficit irrigation_2020–2021 | 89.0 ± 2.69 c | 1.84 ± 0.04 c | 2.17 ± 0.05 c | 3.67 ± 0.05 c | 5.85 ± 0.09 c | 36.9 ± 0.20 c | ||
| Treatment | ||||||||
| Control | 80.6 ± 2.72 d | 1.74 ± 0.05 d | 1.94 ± 0.04 d | 3.44 ± 0.04 d | 5.39 ± 0.09 d | 35.8 ± 0.23 d | ||
| PGPR | 122.9 ± 4.28 b | 2.94 ± 0.10 b | 2.38 ± 0.06 c | 3.88 ± 0.06 c | 6.25 ± 0.12 c | 37.7 ± 0.24 c | ||
| Phosphogypsum | 100.5 ± 2.64 c | 2.25 ± 0.07 c | 2.67 ± 0.06 b | 4.17 ± 0.06 b | 6.83 ± 0.12 b | 38.8 ± 0.19 b | ||
| Combined | 162.2 ± 4.55 a | 3.75 ± 0.15 a | 3.21 ± 0.06 a | 4.71 ± 0.06 a | 7.91 ± 0.12 a | 40.3 ± 0.15 a | ||
| Interaction | ||||||||
| Year | Water regime | Treatment | ||||||
| 2019–2020 | Regular Irrig. | Control | 101.1 ± 0.37 e, J | 2.34 ± 0.051 de, GH | 2.25 ± 0.026 d, G | 3.75 ± 0.026 d, G | 6.00 ± 0.053 e, F | 37.5 ± 0.11 d, F |
| PGPR | 151.7 ± 0.26 b, D | 3.70 ± 0.064 b, D | 2.79 ± 0.040 c, EF | 4.29 ± 0.040 c, EF | 7.08 ± 0.080 c, E | 39.4 ± 0.12 c, DE | ||
| Phosphogypsum | 124.2 ± 0.11 c, F | 3.01 ± 0.103 c, E | 3.09 ± 0.068 b, CD | 4.59 ± 0.068 b, CD | 7.67 ± 0.050 b, CD | 40.2 ± 0.17 b, BC | ||
| Combined | 192.7 ± 0.12 a, B | 4.91 ± 0.104 a, B | 3.58 ± 0.084 a, B | 5.08 ± 0.090 a, B | 8.67 ± 0.050 a, B | 41.3 ± 0.33 a, A | ||
| Deficit Irrig. | Control | 52.7 ± 0.30 h, O | 1.28 ± 0.041 g, L | 1.49 ± 0.078 g, J | 2.99 ± 0.078 g, J | 4.49 ± 0.155 h, I | 33.3 ± 0.27 g, I | |
| PGPR | 86.0 ± 0.10 f, K | 2.18 ± 0.042 e, HI | 1.75 ± 0.031 f, I | 3.25 ± 0.031 f, I | 4.99 ± 0.061 g, H | 35.0 ± 0.18 f, H | ||
| Phosphogypsum | 74.2 ± 0.20 g, M | 1.69 ± 0.036 f, K | 2.05 ± 0.061 e, H | 3.55 ± 0.061 e, H | 5.60 ± 0.122 f, G | 36.6 ± 0.29 e, G | ||
| Combined | 120.5 ± 0.44 d, H | 2.50 ± 0.012 d, FG | 2.65 ± 0.049 c, F | 4.15 ± 0.049 c, F | 6.79 ± 0.099 d, E | 39.0 ± 0.16 c, E | ||
| 2020–2021 | Regular Irrig. | Control | 106.7 ± 0.05 e, I | 2.02 ± 0.016 e, IJ | 2.37 ± 0.017 e, G | 3.87 ± 0.017 e, G | 6.24 ± 0.035 e, F | 38.0 ± 0.07 e, F |
| PGPR | 167.3 ± 0.43 b, C | 3.88 ± 0.073 b, C | 2.95 ± 0.061 c, DE | 4.45 ± 0.061 c, DE | 7.40 ± 0.122 c, D | 39.9 ± 0.17 c, CD | ||
| Phosphogypsum | 122.1 ± 0.10 d, G | 2.63 ± 0.029 c, F | 3.23 ± 0.036 b, C | 4.73 ± 0.036 b, C | 7.96 ± 0.072 b, C | 40.6 ± 0.09 b, B | ||
| Combined | 209.5 ± 0.27 a, A | 5.21 ± 0.027 a, A | 3.87 ± 0.015 a, A | 5.37 ± 0.015 a, A | 9.23 ± 0.031 a, A | 41.9 ± 0.03 a, A | ||
| Deficit Irrig. | Control | 62.0 ± 0.10 h, N | 1.34 ± 0.069 g, L | 1.66 ± 0.049 g, I | 3.16 ± 0.049 g, I | 4.83 ± 0.099 g, H | 34.5 ± 0.31 g, H | |
| PGPR | 86.6 ± 0.17 f, K | 1.99 ± 0.064 e, J | 2.01 ± 0.049 f, H | 3.51 ± 0.049 f, H | 5.53 ± 0.099 f, G | 36.4 ± 0.24 f, G | ||
| Phosphogypsum | 81.4 ± 0.38 g, L | 1.67 ± 0.053 f, K | 2.29 ± 0.045 e, G | 3.79 ± 0.045 e, G | 6.09 ± 0.090 e, F | 37.7 ± 0.18 e, F | ||
| Combined | 126.1 ± 0.38 c, E | 2.35 ± 0.035 d, G | 2.73 ± 0.085 d, F | 4.23 ± 0.085 d, F | 6.95 ± 0.170 d, E | 39.2 ± 0.24 d, E | ||
| F-value (Two-way) | ||||||||
| Water regime | 204,669.72 *** | 3043.54 *** | 1247.59 *** | 1221.45 *** | 1604.70 *** | 1146.44 *** | ||
| Treatment | 200,957.53 *** | 2722.49 *** | 1151.69 *** | 1127.57 *** | 1481.35 *** | 1033.50 *** | ||
| Interaction | 4679.00 *** | 203.77 *** | 8.71 *** | 8.53 *** | 11.20 *** | 13.27 *** | ||
| Year | 2019–2020 | 2020–2021 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Temperature (°C) | Rainfall (mm) | Relative Humidity (%) | Temperature (°C) | Rainfall (mm) | Relative Humidity (%) | ||||
| Month | Max | Min | Max | Min | |||||
| October | 26.3 | 17.2 | 0.98 | 32.6 | 25.3 | 16.2 | 0.94 | 31.6 | |
| December | 25.9 | 15.3 | 0.85 | 34.2 | 24.9 | 14.3 | 0.82 | 33.2 | |
| January | 24.5 | 13.2 | 1.1 | 35.1 | 23.2 | 12.4 | 0.54 | 32.7 | |
| February | 22.3 | 10.3 | 3.1 | 46.2 | 20.3 | 11.1 | 3.32 | 42.4 | |
| March | 21.4 | 9.7 | 6.4 | 44.3 | 20.6 | 10.7 | 6.85 | 43.1 | |
| April | 23.7 | 13.8 | 0.5 | 43.8 | 22.5 | 12.5 | 0.63 | 44.8 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Shamy, M.A.; Alshaal, T.; Mohamed, H.H.; Rady, A.M.S.; Hafez, E.M.; Alsohim, A.S.; Abd El-Moneim, D. Quinoa Response to Application of Phosphogypsum and Plant Growth-Promoting Rhizobacteria under Water Stress Associated with Salt-Affected Soil. Plants 2022, 11, 872. https://doi.org/10.3390/plants11070872
El-Shamy MA, Alshaal T, Mohamed HH, Rady AMS, Hafez EM, Alsohim AS, Abd El-Moneim D. Quinoa Response to Application of Phosphogypsum and Plant Growth-Promoting Rhizobacteria under Water Stress Associated with Salt-Affected Soil. Plants. 2022; 11(7):872. https://doi.org/10.3390/plants11070872
Chicago/Turabian StyleEl-Shamy, Moshira A., Tarek Alshaal, Hossam Hussein Mohamed, Asmaa M. S. Rady, Emad M. Hafez, Abdullah S. Alsohim, and Diaa Abd El-Moneim. 2022. "Quinoa Response to Application of Phosphogypsum and Plant Growth-Promoting Rhizobacteria under Water Stress Associated with Salt-Affected Soil" Plants 11, no. 7: 872. https://doi.org/10.3390/plants11070872
APA StyleEl-Shamy, M. A., Alshaal, T., Mohamed, H. H., Rady, A. M. S., Hafez, E. M., Alsohim, A. S., & Abd El-Moneim, D. (2022). Quinoa Response to Application of Phosphogypsum and Plant Growth-Promoting Rhizobacteria under Water Stress Associated with Salt-Affected Soil. Plants, 11(7), 872. https://doi.org/10.3390/plants11070872