
GC/MS Profiling, Anti-Collagenase, Anti-Elastase, Anti-Tyrosinase and Anti-Hyaluronidase Activities of a Stenocarpus sinuatus Leaves Extract

 ,
,  , ,
, ,  ,
, 
Abstract
:1. Introduction
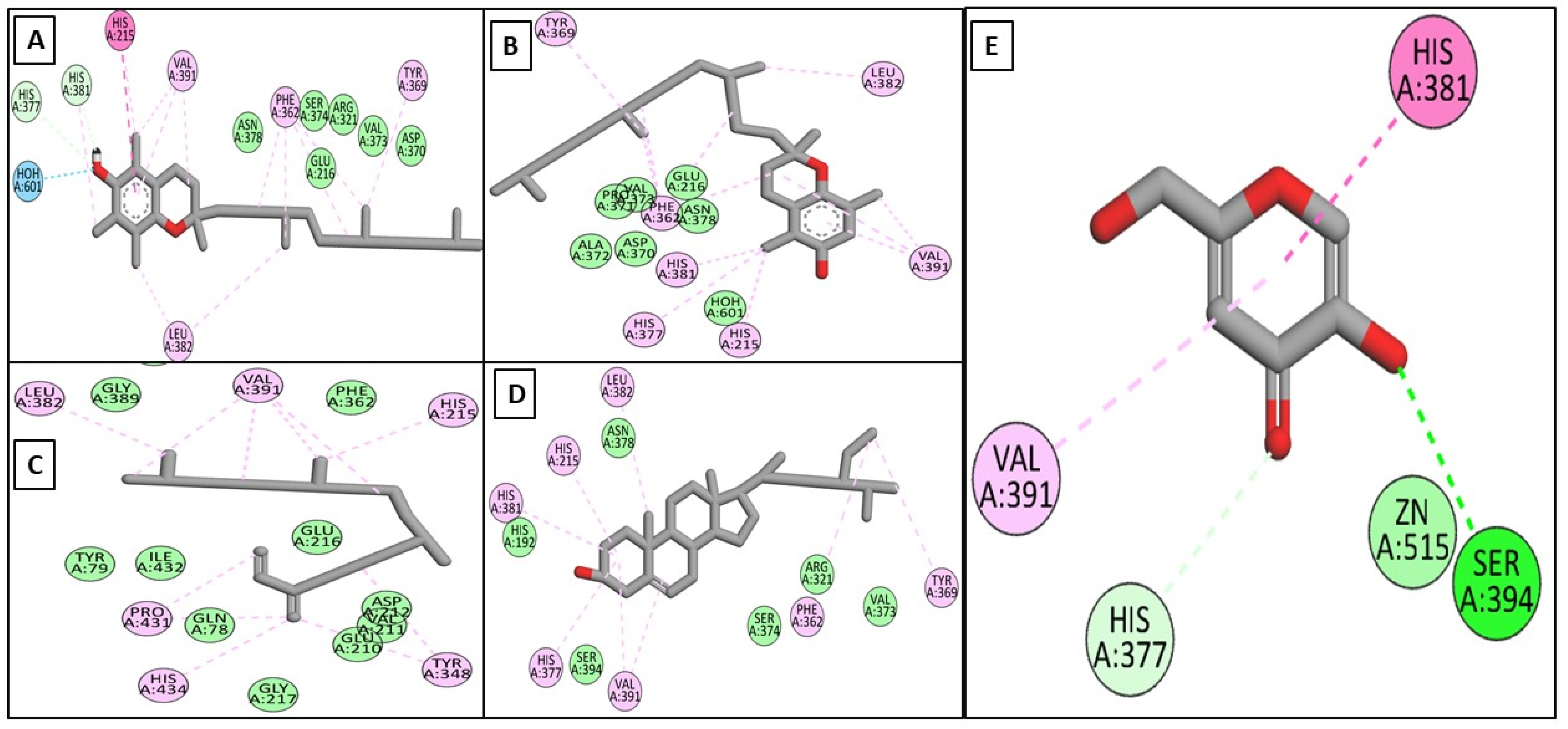
2. Results and Discussion
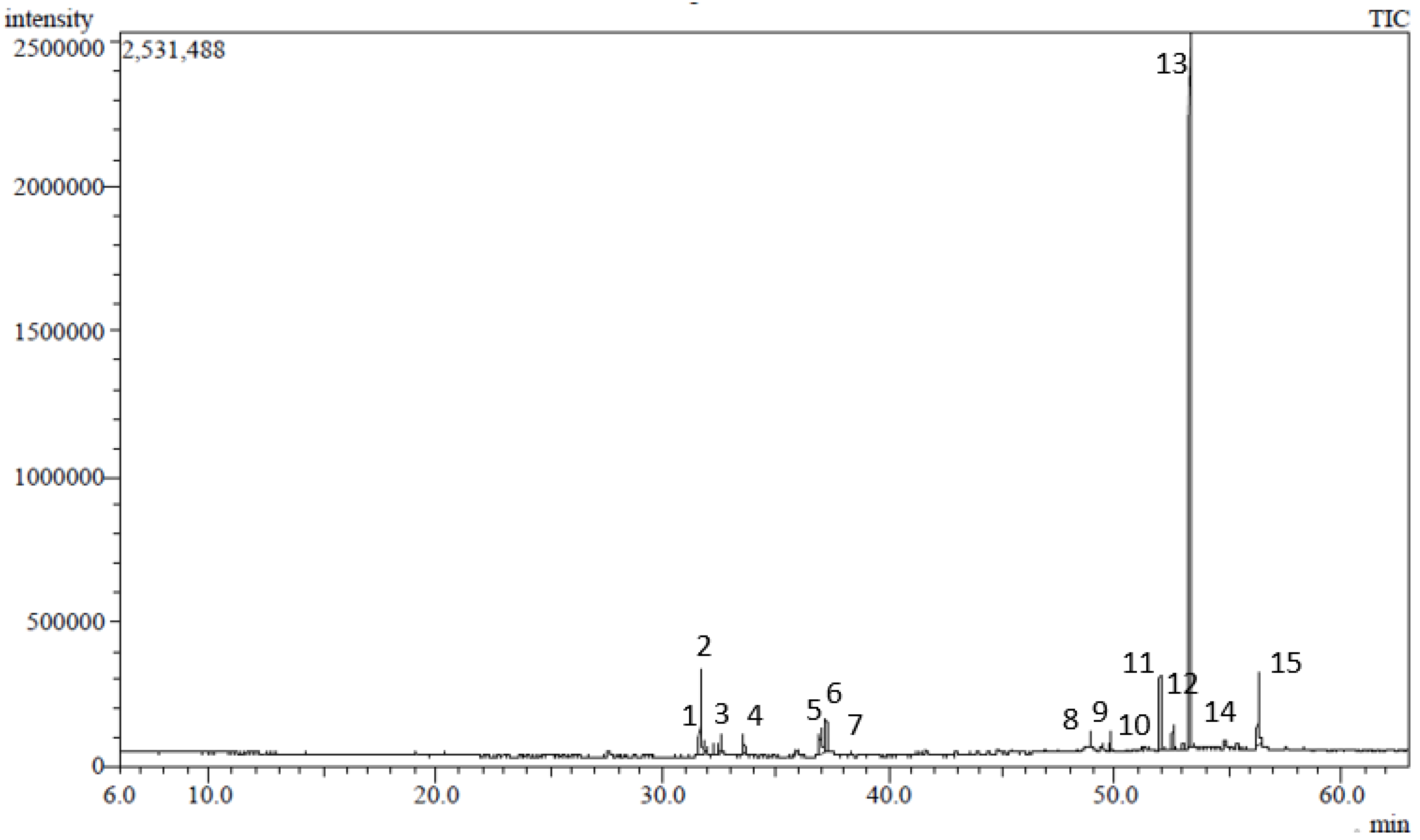
2.1. GC/MS Analysis of the Hexane-Soluble Fraction of a Stenocarpus sinuatus Extract
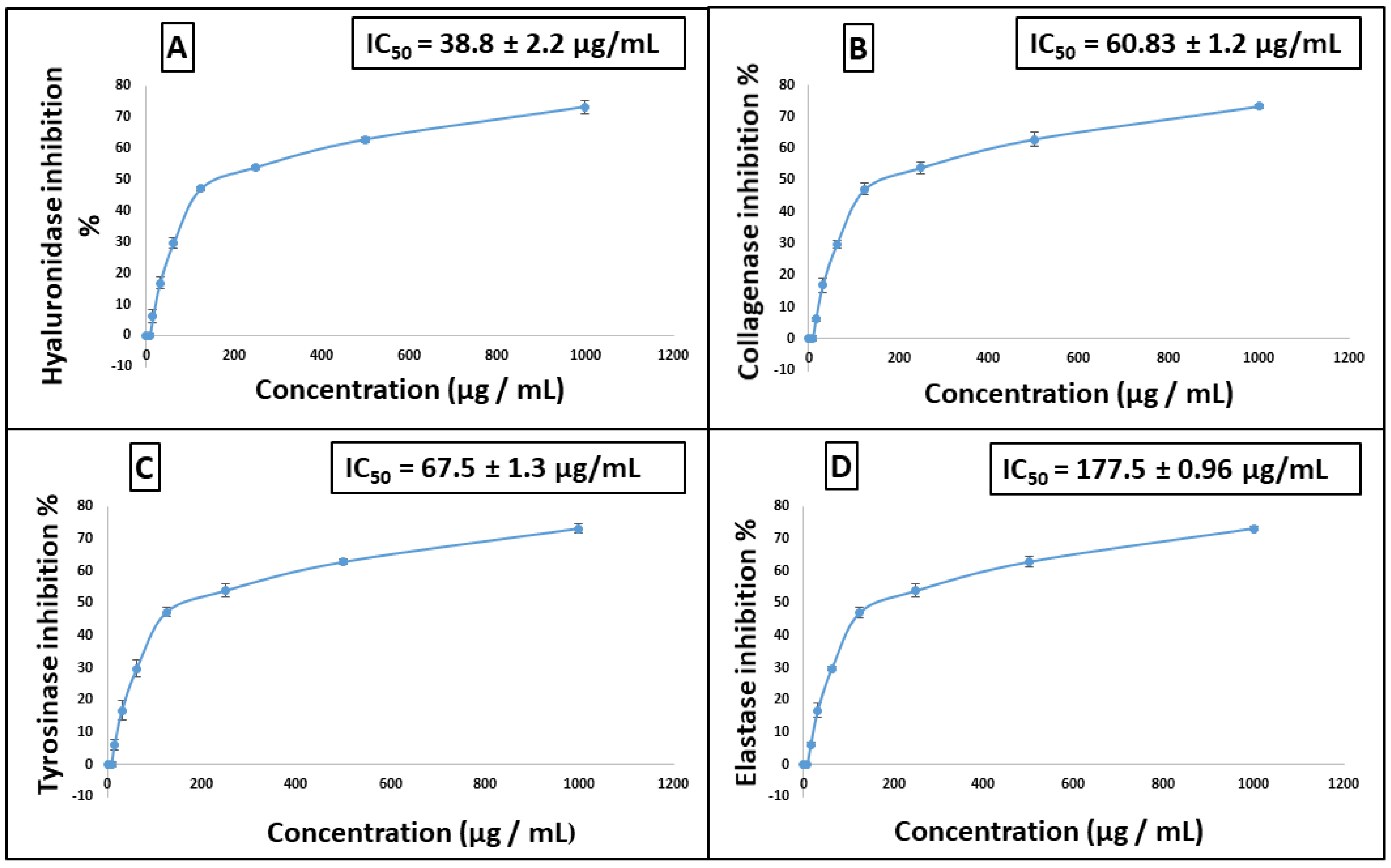
2.2. In Vitro Assays to Evaluate the Antiaging Potential of Stenocarpus sinuatus Leaf Hexane-Soluble Fraction
2.3. In Silico Studies
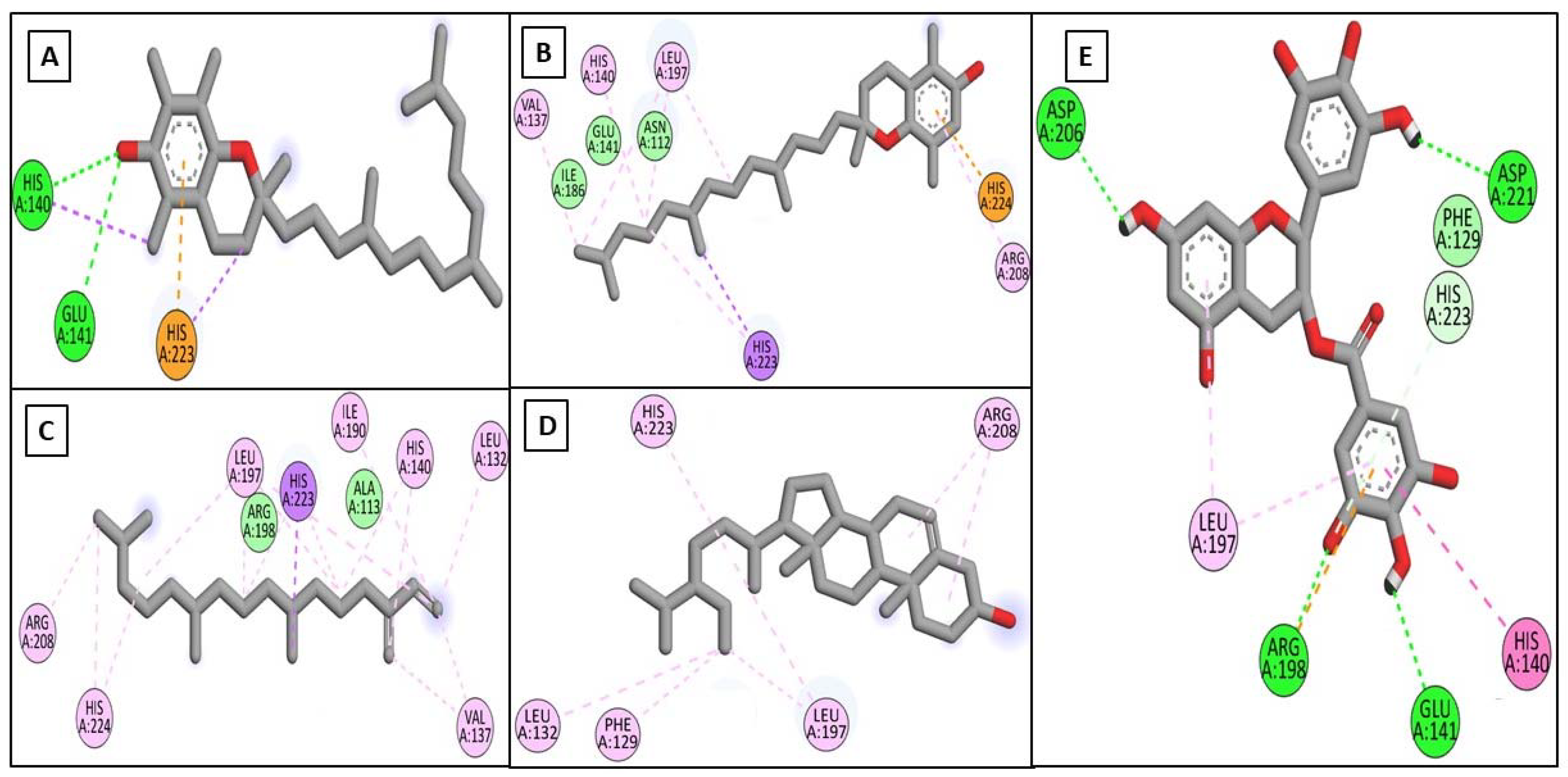
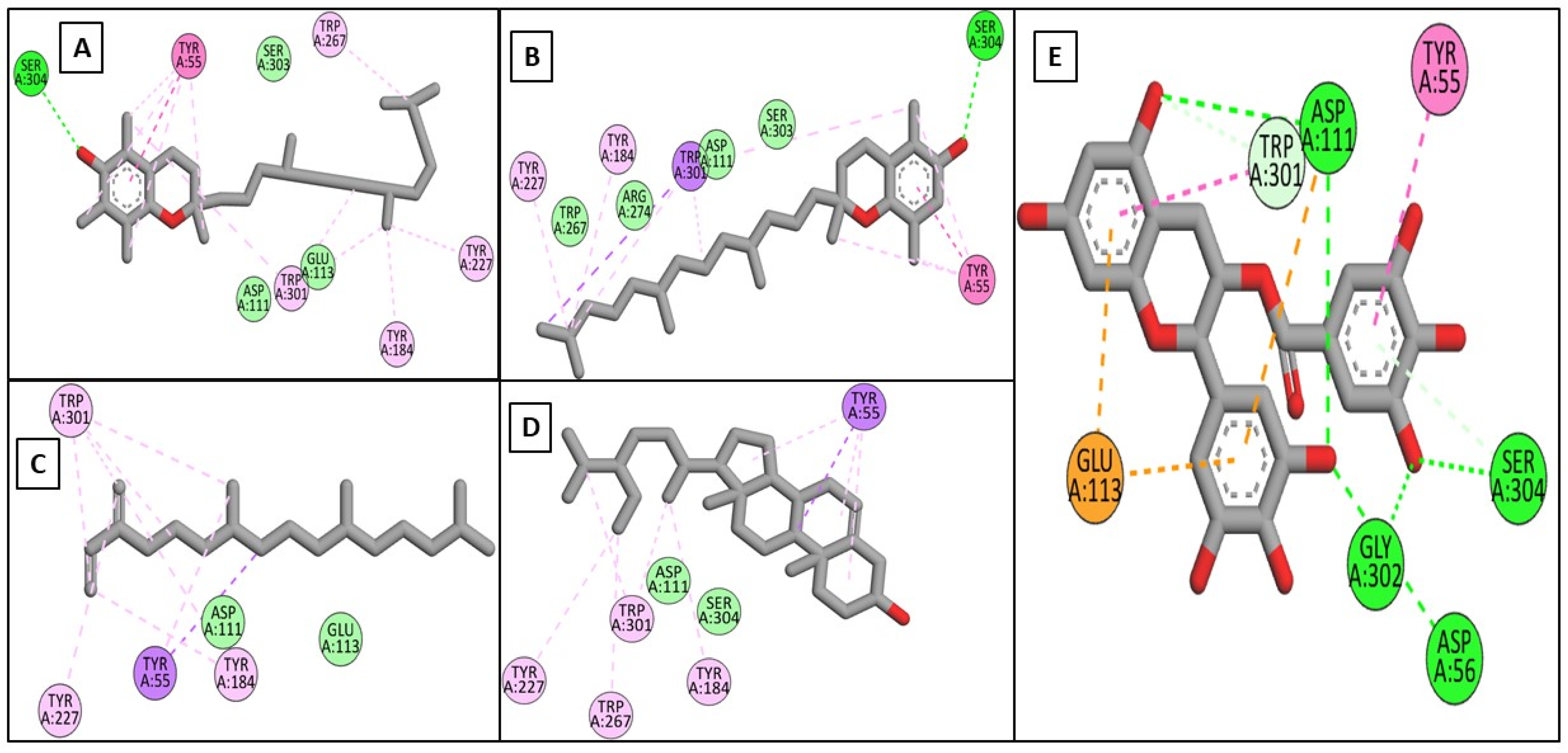
2.3.1. In Silico Docking Studies on the Target Enzymes
2.3.2. In Silico Toxicity Study
3. Materials and Methods
3.1. Plant Material
3.2. Chemicals and Reagents
3.3. Preparation of the Extracts
3.4. GC/MS Analysis of the Hexane-Soluble Fraction
3.5. In Vitro Antiaging Assays
3.5.1. Anti-Collagenase Assay
3.5.2. Anti-Elastase Assay
3.5.3. Anti-Tyrosinase Assay
3.5.4. Anti-Hyaluronidase Assay
3.6. In Silico Studies
3.6.1. In Silico Docking Studies on the Target Enzymes
3.6.2. In Silico Toxicity Study
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yagi, M.; Takabe, W.; Ishizaki, K.; Yonei, Y. The glycative stress and skin aging, and its countermeasure. Fragr. J. 2017, 45, 14–21. [Google Scholar]
- Ganceviciene, R.; Liakou, A.I.; Theodoridis, A.; Makrantonaki, E.; Zouboulis, C.C. Skin anti-aging strategies. Derm.-Endocrinol. 2012, 4, 308–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavinato, M.; Waltenberger, B.; Baraldo, G.; Grade, C.V.C.; Stuppner, H.; Jansen-Duerr, P. Plant extracts and natural compounds used against UVB-induced photoaging. Biogerontology 2017, 18, 499–516. [Google Scholar] [CrossRef] [PubMed]
- Banglao, W.; Thongmee, A.; Sukplang, P.; Wanakhachornkrai, O. Determination of antioxidant, anti-aging and cytotoxicity activity of the essential oils from Cinnamomum zeylanicum. J. Microbiol. Biotechnol. Food Sci. 2020, 10, 436–440. [Google Scholar] [CrossRef]
- Fayad, S.; Morin, P.; Nehmé, R. Use of chromatographic and electrophoretic tools for assaying elastase, collagenase, hyaluronidase, and tyrosinase activity. J. Chromatogr. A 2017, 1529, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Abd Razak, D.L.; Jamaluddin, A.; Abd Rashid, N.Y.; Sani, N.A.; Abdul Manan, M. Assessment of cosmeceutical potentials of selected mushroom fruitbody extracts through evaluation of antioxidant, anti-hyaluronidase and anti-tyrosinase activity. J 2020, 3, 329–342. [Google Scholar] [CrossRef]
- Mostafa, N.M.; Mostafa, A.M.; Ashour, M.L.; Elhady, S.S. Neuroprotective effects of black pepper cold-pressed oil on scopolamine-induced oxidative stress and memory impairment in rats. Antioxidants 2021, 10, 1993. [Google Scholar] [CrossRef]
- El-Nashar, H.A.S.; Mostafa, N.M.; Eldahshan, O.A.; Singab, A.N.B. A new antidiabetic and anti-inflammatory biflavonoid from Schinus polygama (Cav.) Cabrera leaves. Nat. Prod. Res. 2020, 36, 1182–1190. [Google Scholar] [CrossRef]
- Abdallah, S.H.; Mostafa, N.M.; Mohamed, M.A.; Nada, A.S.; Singab, A.N.B. UPLC-ESI-MS/MS profiling and hepatoprotective activities of Stevia leaves extract, butanol fraction and stevioside against radiation-induced toxicity in rats. Nat. Prod. Res. 2021, 1–7. [Google Scholar] [CrossRef]
- Al-Madhagy, S.A.; Mostafa, N.M.; Youssef, F.S.; Awad, G.E.A.; Eldahshan, O.A.; Singab, A.N.B. Metabolic profiling of a polyphenolic-rich fraction of Coccinia grandis leaves using LC-ESI-MS/MS and in vivo validation of its antimicrobial and wound healing activities. Food Funct. 2019, 10, 6267–6275. [Google Scholar] [CrossRef]
- Mostafa, N.M.; Abd El-Ghffar, E.A.; Hegazy, H.G.; Eldahshan, O.A. New methoxyflavone from Casimiroa sapota and the biological activities of its leaves extract against lead acetate induced hepatotoxicity in rats. Chem. Biodivers. 2018, 15, e1700528. [Google Scholar] [CrossRef] [PubMed]
- Shahat, E.A.; Bakr, R.O.; Eldahshan, O.A.; Ayoub, N.A. Chemical composition and biological activities of the essential oil from leaves and flowers of Pulicaria incisa sub. candolleana (Family Asteraceae). Chem. Biodivers. 2017, 14, e1600156. [Google Scholar] [CrossRef] [PubMed]
- Singab, A.N.B.; Mostafa, N.M.; Eldahshan, O.A.; Ashour, M.L.; Wink, M. Profile of volatile components of hydrodistilled and extracted leaves of Jacaranda acutifolia and their antimicrobial activity against foodborne pathogens. Nat. Prod. Commun. 2014, 9, 1934578X1400900731. [Google Scholar] [CrossRef] [Green Version]
- Todirascu-Ciornea, E.; El-Nashar, H.A.S.; Mostafa, N.M.; Eldahshan, O.A.; Boiangiu, R.S.; Dumitru, G.; Hritcu, L.; Singab, A.N.B. Schinus terebinthifolius essential oil attenuates scopolamine-induced memory deficits via cholinergic modulation and antioxidant properties in a zebrafish model. Evid.-Based Complement. Altern. Med. 2019, 2019, 5256781. [Google Scholar] [CrossRef] [Green Version]
- Faheem, S.A.; Saeed, N.M.; El-Naga, R.N.; Ayoub, I.M.; Azab, S.S. Hepatoprotective effect of cranberry nutraceutical extract in non-alcoholic fatty liver model in rats: Impact on insulin resistance and Nrf-2 expression. Front. Pharmacol. 2020, 11, 218. [Google Scholar] [CrossRef] [Green Version]
- Divya, G.; Albert, A.; Singab, A.N.B.; Ayoub, I.M.; Al-Sayed, E.; Paul, E.; Manoharan, K.; Saso, L.; Selvam, G.S. Renoprotective effect of tectorigenin glycosides isolated from Iris spuria L. (Zeal) against hyperoxaluria and hyperglycemia in NRK-49Fcells. Nat. Prod. Res. 2021, 35, 1029–1034. [Google Scholar] [CrossRef]
- Ayoub, I.M.; George, M.Y.; Menze, E.T.; Mahmoud, M.; Botros, M.; Essam, M.; Ashmawy, I.; Shendi, P.; Hany, A.; Galal, M.; et al. Insights into the neuroprotective effects of Salvia officinalis L. and Salvia microphylla Kunth in the memory impairment rat model. Food Funct. 2022. [Google Scholar] [CrossRef]
- Elkady, W.M.; Ayoub, I.M.; Abdel-Mottaleb, Y.; ElShafie, M.F.; Wink, M. Euryops pectinatus L. Flower extract inhibits P-glycoprotein and reverses multi-drug resistance in cancer cells: A mechanistic study. Molecules 2020, 25, 647. [Google Scholar] [CrossRef] [Green Version]
- Brinza, I.; Ayoub, I.M.; Eldahshan, O.A.; Hritcu, L. Baicalein 5,6-dimethyl ether prevents memory deficits in the scopolamine zebrafish model by regulating cholinergic and antioxidant systems. Plants 2021, 10, 1245. [Google Scholar] [CrossRef]
- Younis, I.Y.; Eldahshan, O.A.; Abdel-Aziz, M.A.; Ali, Z.Y. Green synthesis of magnesium nanoparticles mediated from Rosa floribunda charisma extract and its antioxidant, antiaging and antibiofilm activities. Sci. Rep. 2021, 11, 16868. [Google Scholar]
- Amuka, O.; Machocho, A.K.; Okemo, P.O.; Mbugua, P.K. Antifungal and antibacterial activity of crude stem bark extracts’ of Bersama abysinicca Verdc. and Faurea saligna Harr. Res. J. Med. Plant 2015, 9, 160–169. [Google Scholar] [CrossRef] [Green Version]
- Deans, B.J.; Kilah, N.L.; Bissember, A.C.; Smith, J.A.; Jordan, G.J. Arbutin derivatives isolated from ancient Proteaceae: Potential phytochemical markers present in Bellendena, Cenarrhenes, and Persoonia Genera. J. Nat. Prod. 2018, 81, 1241–1251. [Google Scholar] [CrossRef] [PubMed]
- Fiorito, S.; Genovese, S.; Epifano, F.; Mathieu, V.; Kiss, R.; Taddeo, V.A. Cytotoxic activity of lomatiol and 7-(3′-Hydroxymethyl-3′-methylallyloxy)coumarin. Nat. Prod. Commun. 2016, 11, 1934578X1601100322. [Google Scholar] [CrossRef] [Green Version]
- Giang, P.M.; Thao, D.T.; Nga, N.T.; Van Trung, B.; Anh, D.H.; Viet, P.H. Evaluation of the antioxidant, hepatoprotective, and anti-inflammatory activities of bisresorcinol isolated from the trunk of Heliciopsis Terminalis. Pharm. Chem. J. 2019, 53, 628–634. [Google Scholar] [CrossRef]
- Vinueza, D.; Yanza, K.; Tacchini, M.; Grandini, A.; Sacchetti, G.; Chiurato, M.A.; Guerrini, A. Flavonoids in Ecuadorian Oreocallis grandiflora (Lam.) R. Br.: Perspectives of use of this species as a food supplement. Evid.-Based Complement. Altern. Med. 2018, 2018, 1353129. [Google Scholar] [CrossRef]
- Wang, H.; Leach, D.N.; Thomas, M.C.; Blanksby, S.J.; Forster, P.I.; Waterman, P.G. Bisresorcinol derivatives from Grevillea glauca. Helv. Chim. Acta 2011, 94, 1812–1819. [Google Scholar] [CrossRef]
- Yang, F.; Zhao, H.; Carroll, A.R. Tropane alkaloids from the Australian plant Triunia montana (Proteaceae). Tetrahedron Lett. 2017, 58, 736–739. [Google Scholar] [CrossRef]
- Monzote, L.; Piñón, A.; Setzer, W.N. Antileishmanial potential of tropical rainforest plant extracts. Medicines 2014, 1, 32–55. [Google Scholar] [CrossRef] [Green Version]
- Tong, J.; Zhou, Z.; Qi, W.; Jiang, S.; Yang, B.; Zhong, Z.; Jia, Y.; Li, X.; Xiong, L.; Nie, L. Antidepressant effect of helicid in chronic unpredictable mild stress model in rats. Int. Immunopharmacol. 2019, 67, 13–21. [Google Scholar] [CrossRef]
- El Hawary, S.S.; Abubaker, M.; Abd El-Kader, E.M.; Mahrous, E.A. Phytochemical constituents and anti-tyrosinase activity of Macadamia integrifolia leaves extract. Nat. Prod. Res. 2020, 36, 1089–1094. [Google Scholar] [CrossRef]
- Abubaker, M.; El Hawary, S.S.; Mahrous, E.A.; Abd El-Kader, E.M. Study of Nutritional contents of Macadamia integrifolia Maiden & Betche leaves, kernel and pericarp cultivated in Egypt. Int. J. Pharmacogn. Phytochem. Res. 2017, 9, 1442–1445. [Google Scholar]
- Wall, M.M. Functional lipid characteristics, oxidative stability, and antioxidant activity of macadamia nut (Macadamia integrifolia) cultivars. Food Chem. 2010, 121, 1103–1108. [Google Scholar] [CrossRef]
- Kamagaju, L.; Morandini, R.; Bizuru, E.; Nyetera, P.; Nduwayezu, J.B.; Stevigny, C.; Ghanem, G.; Duez, P. Tyrosinase modulation by five Rwandese herbal medicines traditionally used for skin treatment. J. Ethnopharmacol. 2013, 146, 824–834. [Google Scholar] [CrossRef]
- Chuang, T.-H.; Chan, H.-H.; Wu, T.-S.; Li, C.-F. Chemical constituents and biological studies of the leaves of Grevillea robusta. Molecules 2011, 16, 9331–9339. [Google Scholar] [CrossRef] [Green Version]
- Vinueza, D.; Cajamarca, D.; Acosta, K.; Pilco, G. Oreocallis grandiflora photoprotective effect against ultraviolet B radiation-induced cell death. Asian J. Pharm. Clin. Res. 2018, 11, 276–280. [Google Scholar] [CrossRef] [Green Version]
- Mock, J.; Murphy, S.T.; Ritchie, E.; Taylor, W.C. Chemical studies of the Proteaceae. VI. Two naphthoquinones from Stenocarpus salignus. Aust. J. Chem. 1973, 26, 1121–1130. [Google Scholar] [CrossRef]
- Dorey, J.B. Missing for almost 100 years: The rare and potentially threatened bee, Pharohylaeus lactiferus (Hymenoptera, Colletidae). J. Hymenopt. Res. 2021, 81, 165–180. [Google Scholar] [CrossRef]
- Kerdpol, K.; Nutho, B.; Krusong, K.; Poo-arporn, R.P.; Rungrotmongkol, T.; Hannongbua, S. Encapsulation of α-tocopherol in large-ring cyclodextrin containing 26 α-D-glucopyranose units: A molecular dynamics study. J. Mol. Liq. 2021, 339, 116802. [Google Scholar] [CrossRef]
- Bruno, R.S.; Mah, E.; Vitamin, E. Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Ching, L.S.; Mohamed, S. Alpha-tocopherol content in 62 edible tropical plants. J. Agric. Food Chem. 2001, 49, 3101–3105. [Google Scholar] [CrossRef]
- García, M.F.; Vergara, C.E.; Forero-Doria, O.; Guzman, L.; del Carmen Perez-Camino, M. Chemical evaluation and thermal behavior of Chilean hazelnut oil (Gevuina avellana Mol) a comparative study with extra virgin olive oil. Eur. Food Res. Technol. 2019, 245, 1021–1029. [Google Scholar] [CrossRef] [Green Version]
- Medel, F.; Nunez, R.; Medel, G.; Palma, H.; Manquian, N.; Fuentes, R. Fractions of vitamin E (tocotrienols and tocopherols) in nut oil of Gevuina avellana Mol. Acta Hortic. 2009, 845, 687–691. [Google Scholar] [CrossRef]
- Rattanawiwatpong, P.; Wanitphakdeedecha, R.; Bumrungpert, A.; Maiprasert, M. Anti-aging and brightening effects of a topical treatment containing vitamin C, vitamin E, and raspberry leaf cell culture extract: A split-face, randomized controlled trial. J. Cosmet. Dermatol. 2020, 19, 671–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villareal, M.O.; Kume, S.; Bourhim, T.; Bakhtaoui, F.Z.; Kashiwagi, K.; Han, J.; Gadhi, C.; Isoda, H. Activation of MITF by argan oil leads to the inhibition of the tyrosinase and dopachrome tautomerase expressions in B16 murine melanoma cells. Evid.-Based Complement. Altern. Med. 2013, 2013, 340107. [Google Scholar] [CrossRef] [Green Version]
- Ricciarelli, R.; Maroni, P.; Özer, N.; Zingg, J.-M.; Azzi, A. Age-dependent increase of collagenase expression can be reduced by α-tocopherol via protein kinase C inhibition. Free. Radic. Biol. Med. 1999, 27, 729–737. [Google Scholar] [CrossRef]
- Heo, S.-I.; Jung, M.-J.; Kim, M.-K.; Wang, M.-H. Antioxidative activities and tyrosinase inhibitory effects of Korean medicinal plants. J. Appl. Biol. Chem. 2007, 50, 115–119. [Google Scholar]
- Yenilmez, E.; Başaran, E.; Yazan, Y. Release characteristics of vitamin E incorporated chitosan microspheres and in vitro–in vivo evaluation for topical application. Carbohydr. Polym. 2011, 84, 807–811. [Google Scholar] [CrossRef]
- Poljšak, N.; Kočevar Glavač, N. Tilia sp. Seed Oil—Composition, antioxidant activity and potential use. Appl. Sci. 2021, 11, 4932. [Google Scholar] [CrossRef]
- Doering, T.; Holtkötter, O.; Schlotmann, K.; Jassoy, C.; Petersohn, D.; Wadle, A.; Waldmann-Laue, M. Cutaneous restructuration by apple seed phytosterols: From DNA chip analysis to morphological alterations. Int. J. Cosmet. Sci. 2005, 27, 142. [Google Scholar] [CrossRef]
- Edmond, M.P.; Mostafa, N.M.; El-Shazly, M.; Singab, A.N.B. Two clerodane diterpenes isolated from Polyalthia longifolia leaves: Comparative structural features, anti-histaminic and anti-Helicobacter pylori activities. Nat. Prod. Res. 2021, 35, 5282–5286. [Google Scholar] [CrossRef]
- Mostafa, N.M.; Edmond, M.P.; El-Shazly, M.; Fahmy, H.A.; Sherif, N.H.; Singab, A.N.B. Phytoconstituents and renoprotective effect of Polyalthia longifolia leaves extract on radiation-induced nephritis in rats via TGF-β/smad pathway. Nat. Prod. Res. 2021, 1–6. [Google Scholar] [CrossRef]
- Jeong, S.H. Inhibitory effect of phytol on cellular senescence. Biomed. Dermatol. 2018, 2, 13. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.T.; Ali, E.S.; Uddin, S.J.; Shaw, S.; Islam, M.A.; Ahmed, M.I.; Shill, M.C.; Karmakar, U.K.; Yarla, N.S.; Khan, I.N. Phytol: A review of biomedical activities. Food Chem. Toxicol. 2018, 121, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Yasmeen, S.; Gupta, P. Interaction of selected terpenoids from Dalbergia sissoo with catalytic domain of matrix metalloproteinase-1: An in silico assessment of their anti-wrinkling potential. Bioinform. Biol. Insights 2019, 13, 1177932219896538. [Google Scholar] [CrossRef] [PubMed]
- Temin, P. GC- MS Analysis of Mussaenda roxburghii Hk.f.: A folk food plant used among tribes of Arunachal Pradesh, India. Pharmacogn. J. 2016, 8, 395–398. [Google Scholar]
- Ak, G.; Zengin, G.; Ceylan, R.; Fawzi Mahomoodally, M.; Jugreet, S.; Mollica, A.; Stefanucci, A. Chemical composition and biological activities of essential oils from Calendula officinalis L. flowers and leaves. Flavour Fragr. J. 2021, 36, 554–563. [Google Scholar] [CrossRef]
- Mahalakashmi, R.; Thangapandian, V. Gas chromatography and mass spectrometry analysis of bioactive constituents of Maytenus heyneana (Roth) Roju & Babu (Celestraceae). J. Pharmacogn. Phytochem. 2019, 8, 2748–2752. [Google Scholar]
- Mostafa, N.M.; Ashour, M.L.; Eldahshan, O.A.; Singab, A.N.B. Cytotoxic activity and molecular docking of a novel biflavonoid isolated from Jacaranda acutifolia (Bignoniaceae). Nat. Prod. Res. 2016, 30, 2093–2100. [Google Scholar] [CrossRef]
- Moussa, A.Y.; Mostafa, N.M.; Singab, A.N.B. Pulchranin A: First report of isolation from an endophytic fungus and its inhibitory activity on cyclin dependent kinases. Nat. Prod. Res. 2020, 34, 2715–2722. [Google Scholar] [CrossRef]
- Ashmawy, A.; Mostafa, N.; Eldahshan, O. GC/MS analysis and molecular profiling of lemon volatile oil against breast cancer. J. Essent. Oil Bear. Plants 2019, 22, 903–916. [Google Scholar] [CrossRef]
- Elhawary, E.A.; Mostafa, N.M.; Labib, R.M.; Singab, A.N. Metabolomic profiles of essential oils from selected rosa varieties and their antimicrobial activities. Plants 2021, 10, 1721. [Google Scholar] [CrossRef]
- El-Nashar, H.A.S.; Mostafa, N.M.; El-Badry, M.A.; Eldahshan, O.A.; Singab, A.N.B. Chemical composition, antimicrobial and cytotoxic activities of essential oils from Schinus polygamus (Cav.) cabrera leaf and bark grown in Egypt. Nat. Prod. Res. 2020, 35, 5369–5372. [Google Scholar] [CrossRef] [PubMed]
- Ashmawy, A.M.; Ayoub, I.M.; Eldahshan, O.A. Chemical composition, cytotoxicity and molecular profiling of Cordia africana Lam. on human breast cancer cell line. Nat. Prod. Res. 2020, 35, 4133–4138. [Google Scholar] [CrossRef] [PubMed]
- Labib, R.M.; Ayoub, I.M.; Michel, H.E.; Mehanny, M.; Kamil, V.; Hany, M.; Magdy, M.; Moataz, A.; Maged, B.; Mohamed, A. Appraisal on the wound healing potential of Melaleuca alternifolia and Rosmarinus officinalis L. essential oil-loaded chitosan topical preparations. PLoS ONE 2019, 14, e0219561. [Google Scholar] [CrossRef] [PubMed]
- Gad, H.; Al-Sayed, E.; Ayoub, I. Phytochemical discrimination of Pinus species based on GC–MS and ATR-IR analyses and their impact on Helicobacter pylori. Phytochem. Anal. 2021, 32, 820–835. [Google Scholar] [CrossRef]
- Gad, H.A.; Ayoub, I.M.; Wink, M. Phytochemical profiling and seasonal variation of essential oils of three Callistemon species cultivated in Egypt. PLoS ONE 2019, 14, e0219571. [Google Scholar] [CrossRef] [Green Version]
- Ayoub, N.; Singab, A.N.; Mostafa, N.; Schultze, W. Volatile constituents of leaves of Ficus carica Linn. grown in Egypt. J. Essent. Oil Bear. Plants 2010, 13, 316–321. [Google Scholar] [CrossRef]
- Ayoub, I.M.; Korinek, M.; El-Shazly, M.; Wetterauer, B.; El-Beshbishy, H.A.; Hwang, T.-L.; Chen, B.-H.; Chang, F.-R.; Wink, M.; Singab, A.N.B.; et al. Anti-allergic, anti-inflammatory, and anti-hyperglycemic activity of Chasmanthe aethiopica leaf extract and its profiling using LC/MS and GLC/MS. Plants 2021, 10, 1118. [Google Scholar] [CrossRef]
- Korany, D.A.; Ayoub, I.M.; Labib, R.M.; El-Ahmady, S.H.; Singab, A.N.B. The impact of seasonal variation on the volatile profile of leaves and stems of Brownea grandiceps (Jacq.) with evaluation of their anti-mycobacterial and anti-inflammatory activities. S. Afr. J. Bot. 2021, 142, 88–95. [Google Scholar] [CrossRef]
- Thabet, A.A.; Ayoub, I.M.; Youssef, F.S.; Al Sayed, E.; Singab, A.N.B. Essential oils from the leaves and flowers of Leucophyllum frutescens (Scrophulariaceae): Phytochemical analysis and inhibitory effects against elastase and collagenase in vitro. Nat. Prod. Res. 2021, 1–5. [Google Scholar] [CrossRef]
- Azab, S.S.; Abdel Jaleel, G.A.; Eldahshan, O.A. Anti-inflammatory and gastroprotective potential of leaf essential oil of Cinnamomum glanduliferum in ethanol-induced rat experimental gastritis. Pharm. Biol. 2017, 55, 1654–1661. [Google Scholar] [CrossRef] [Green Version]
- Brinza, I.; Abd-Alkhalek, A.M.; El-Raey, M.A.; Boiangiu, R.S.; Eldahshan, O.A.; Hritcu, L. Ameliorative effects of rhoifolin in scopolamine-induced amnesic zebrafish (Danio rerio) model. Antioxidants 2020, 9, 580. [Google Scholar] [CrossRef] [PubMed]
- Thring, T.S.A.; Hili, P.; Naughton, D.P. Anti-collagenase, anti-elastase and anti-oxidant activities of extracts from 21 plants. BMC Complementary Altern. Med. 2009, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.-J.; Uyama, H.; Kobayashi, S. Inhibition effects of (+)-catechin-aldehyde polycondensates on proteinases causing proteolytic degradation of extracellular matrix. Biochem. Biophys. Res. Commun. 2004, 320, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Batubara, I.; Darusman, L.K.; Mitsunaga, T.; Rahminiwati, M.; Djauhari, E. Potency of Indonesian medicinal plants as tyrosinase inhibitor and antioxidant agent. J. Biol. Sci. 2010, 10, 138–144. [Google Scholar] [CrossRef] [Green Version]
- Reissig, J.L.; Strominger, J.L.; Leloir, L.F. A modified colorimetric method for the estimation of N-acetylamino sugars. J. Biol. Chem. 1955, 217, 959–966. [Google Scholar] [CrossRef]
- Sanner, M.F. Python: A programming language for software integration and development. J. Mol. Graph. Model. 1999, 17, 57–61. [Google Scholar] [PubMed]
- Vina, A. Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading Trott, Oleg; Olson, Arthur J. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar]
- Case, D.A.; Cheatham Iii, T.E.; Darden, T.; Gohlke, H.; Luo, R.; Merz, K.M., Jr.; Onufriev, A.; Simmerling, C.; Wang, B.; Woods, R.J. The Amber biomolecular simulation programs. J. Comput. Chem. 2005, 26, 1668–1688. [Google Scholar] [CrossRef] [Green Version]
- Vilar, S.; Cozza, G.; Moro, S. Medicinal chemistry and the molecular operating environment (MOE): Application of QSAR and molecular docking to drug discovery. Curr. Top. Med. Chem. 2008, 8, 1555–1572. [Google Scholar] [CrossRef]
- Elsayed, Z.M.; Eldehna, W.M.; Abdel-Aziz, M.M.; El Hassab, M.A.; Elkaeed, E.B.; Al-Warhi, T.; Abdel-Aziz, H.A.; Abou-Seri, S.M.; Mohammed, E.R. Development of novel isatin–nicotinohydrazide hybrids with potent activity against susceptible/resistant Mycobacterium tuberculosis and bronchitis causing–bacteria. J. Enzym. Inhib. Med. Chem. 2021, 36, 384–393. [Google Scholar] [CrossRef]
- Banerjee, P.; Dehnbostel, F.O.; Preissner, R. Prediction is a balancing act: Importance of sampling methods to balance sensitivity and specificity of predictive models based on imbalanced chemical data sets. Front. Chem. 2018, 362. [Google Scholar] [CrossRef] [PubMed]
- Braga, R.C.; Alves, V.M.; Muratov, E.N.; Strickland, J.; Kleinstreuer, N.; Trospsha, A.; Andrade, C.H. Pred-skin: A fast and reliable web application to assess skin sensitization effect of chemicals. J. Chem. Inf. Model. 2017, 57, 1013–1017. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Rt | Compound | Molecular Formula | RI-exp | RI-lit | Content (%) | Identification |
|---|---|---|---|---|---|---|---|
| 1 | 31.54 | 3,7,11,15-Tetramethyl-2-hexadecene | C20H40 | 1811 | 1811 | 0.26% | MS, RI |
| 2 | 31.67 | Neophytadiene | C20H38 | 1817 | 1817 | 8.19% | MS, RI |
| 3 | 32.58 | 7,11,15-Trimethyl-3-methylenehexadeca-1-ene | C20H38 | 1843 | 1844 | 3.07% | MS, RI |
| 4 | 33.55 | Hexadecanoic acid, methyl ester | C17H34O2 | 1907 | 1907 | 2.22% | MS, RI |
| 5 | 36.89 | 9,12-Octadecadienoic acid, methyl ester | C19H34O2 | 2077 | 2076 | 1.79% | MS, RI |
| 6 | 37.03 | 9,12,15-Octadecatrienoic acid, methyl ester, (Z, Z, Z)- (Linolenic acid, methyl ester) | C19H32O2 | 2084 | 2085 | 4.38% | MS, RI |
| 7 | 37.23 | Phytol | C20H40O | 2095 | 2096 | 4.04% | MS, RI |
| 8 | 48.92 | Squalene | C30H50 | 2794 | 2790 | 1.03% | MS, RI |
| 9 | 49.70 | α-Tocospiro A | C29H50O4 | 2844 | 2855 | 0.64% | MS, RI |
| 10 | 49.80 | 2-Methyloctacosane | C29H60 | 2851 | 2857 | 1.54% | MS, RI |
| 11 | 51.97 | β-Tocopherol | C28H48O2 | 3048 | 3043 | 6.07% | MS, RI |
| 12 | 52.49 | Hentriacontane | C31H64 | 3088 | 3100 | 1.64% | MS |
| 13 | 53.27 | Vitamin E (α-tocopherol) | C29H50O2 | 3146 | 3149 | 52.59% | MS, RI |
| 14 | 54.84 | Campesterol | C28H48O | 3255 | 3193 | 0.66% | MS, RI |
| 15 | 56.31 | γ-Sitosterol | C29H50O | 3346 | 3351 | 8.65% | MS, RI |
| Tocopherols | 58.66% | ||||||
| Acyclic diterpenes | 11.52% | ||||||
| Oxygenated diterpenes | 4.04% | ||||||
| Triterpenes | 1.03% | ||||||
| Sterols | 9.31% | ||||||
| Fatty acid methyl esters | 8.39% | ||||||
| Aliphatic alkanes | 3.18% | ||||||
| Others | 0.64% | ||||||
| Total identified | 96.77% | ||||||
| Compound Name | Collagenase | Elastase | Hyaluronidase | Tyrosinase | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Score | HB | Hydrophobic | Score | HB | Hydrophobic | Score | HB | Hydrophobic | Score | HB | Hydrophobic | |
| α-Tocopherol | −9.02 | ----- | LEU185, LEU218, VAL219, LEU239, TYR244, PHE252, MET253 | −6.00 | HIS140, GLU141 | HIS140, HIS223 | −6.40 | SER304 | TYR55, TYR184, TYR227, TRP267, TRP301 | −7.70 | HIS377, HIS381 | HIS215, PHE362, TYR369, HIS381, LEU382 |
| β-Tocopherol | −8.68 | PRO190 | TYR176, LEU184, LEU185, HIS187, PRO193, HIS222, HIS226, HIS232 | −6.20 | ----- | VAL137, HIS140, LEU197, ARG208, HIS223, HIS224 | −6.40 | SER304 | TYR55, TYR184, TYR227, TRP301 | −7.46 | ----- | HIS215, PHE362, TYR369, HIS377, HIS381, LEU382, VAL391 |
| Neophytadiene | −7.83 | ----- | LEU184, LEU185, LEU218, VAL219, HIS222, LEU239, PHE252, MET252 | −5.20 | ----- | LEU132, VAL137, HIS140, ILE190, LEU197, ARG208, HIS223, HIS224 | −5.40 | ----- | TYR55, TRP184, TYR227, TRP301 | −7.13 | ----- | HIS215, TYR348, LEU382, VAL391, PRO431, HIS434 |
| γ-Sitosterol | −7.66 | ----- | TYR179, LEU184, TYR185, HIS187, PHE189, PRO193, HIS222, HIS232 | −6.10 | ----- | PHE129, LEU132, LEU197, ARG208, HIS223 | −6.90 | ----- | TYR55, TYR184, TYR227, TRP226, TRP301 | −6.84 | ----- | HIS215, PHE362, TYR369, HIS377, HIS381, LEU382, VAL391 |
| EGCG | −8.19 | ALA186, VAL219, GLU223, HIS226, PHE241, ILE243 | VLA219, HIS222 | −8.80 | GLU141, ARG198, ASP206, ASP221, HIS223 | HIS140, LEU197, ARG198 | −8.9 | ASP56, ASP111, TRP301, SER304 | ASP111, GLU113, TRP301 | |||
| Kojic acid | −4.62 | HIS377, SER394 | HIS377, SER394 | |||||||||
| Compound Name | lD50 as Predicted by Pro-toxII | Any Potential Toxicity as Predicted by Pro-toxII | Potential Skin Toxicity as Computed by Pred-Skin 3 Using Bayesian Model | % Probability |
|---|---|---|---|---|
| 3,7,11,15-Tetramethyl-2-hexadecene | 5000 mg/kg | none | None | 95 |
| Neophytadiene | 5050 mg/kg | none | None | 89 |
| Hexadecanoic acid, methyl ester | 5000 mg/kg | none | None | 92 |
| 9,12-Octadecadienoic acid, methyl ester | 20,000 mg/kg | none | None | 95 |
| 9,12,15-Octadecatrienoic acid, methyl ester (Linolenic acid, methyl ester) | 20,000 mg/kg | none | None | 99 |
| Phytol | 5000 mg/kg | none | None | 92 |
| Squalene | 5000 mg/kg | none | None | 95 |
| α-Tocospiro A | 300 mg/kg | cytotoxic | None | 88 |
| 2-Methyloctacosane | 750 mg/kg | none | None | 96 |
| β-Tocopherol | 500 mg/kg | none | None | 90 |
| Hentriacontane | 750 mg/kg | none | None | 98 |
| Vitamin E (α-tocopherol) | 5000 mg/kg | none | None | 89 |
| Campesterol | 890 mg/kg | none | None | 97 |
| γ-Sitosterol | 890 mg/Kg | none | None | 98 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Younis, M.M.; Ayoub, I.M.; Mostafa, N.M.; El Hassab, M.A.; Eldehna, W.M.; Al-Rashood, S.T.; Eldahshan, O.A. GC/MS Profiling, Anti-Collagenase, Anti-Elastase, Anti-Tyrosinase and Anti-Hyaluronidase Activities of a Stenocarpus sinuatus Leaves Extract. Plants 2022, 11, 918. https://doi.org/10.3390/plants11070918
Younis MM, Ayoub IM, Mostafa NM, El Hassab MA, Eldehna WM, Al-Rashood ST, Eldahshan OA. GC/MS Profiling, Anti-Collagenase, Anti-Elastase, Anti-Tyrosinase and Anti-Hyaluronidase Activities of a Stenocarpus sinuatus Leaves Extract. Plants. 2022; 11(7):918. https://doi.org/10.3390/plants11070918
Chicago/Turabian StyleYounis, Mai M., Iriny M. Ayoub, Nada M. Mostafa, Mahmoud A. El Hassab, Wagdy M. Eldehna, Sara T. Al-Rashood, and Omayma A. Eldahshan. 2022. "GC/MS Profiling, Anti-Collagenase, Anti-Elastase, Anti-Tyrosinase and Anti-Hyaluronidase Activities of a Stenocarpus sinuatus Leaves Extract" Plants 11, no. 7: 918. https://doi.org/10.3390/plants11070918
APA StyleYounis, M. M., Ayoub, I. M., Mostafa, N. M., El Hassab, M. A., Eldehna, W. M., Al-Rashood, S. T., & Eldahshan, O. A. (2022). GC/MS Profiling, Anti-Collagenase, Anti-Elastase, Anti-Tyrosinase and Anti-Hyaluronidase Activities of a Stenocarpus sinuatus Leaves Extract. Plants, 11(7), 918. https://doi.org/10.3390/plants11070918