
Exogenous Copper Application for the Elemental Defense of Rice Plants against Rice Leaffolder (Cnaphalocrocis medinalis)

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Cu Impedes the Growth and Development of C. medinalis Larvae
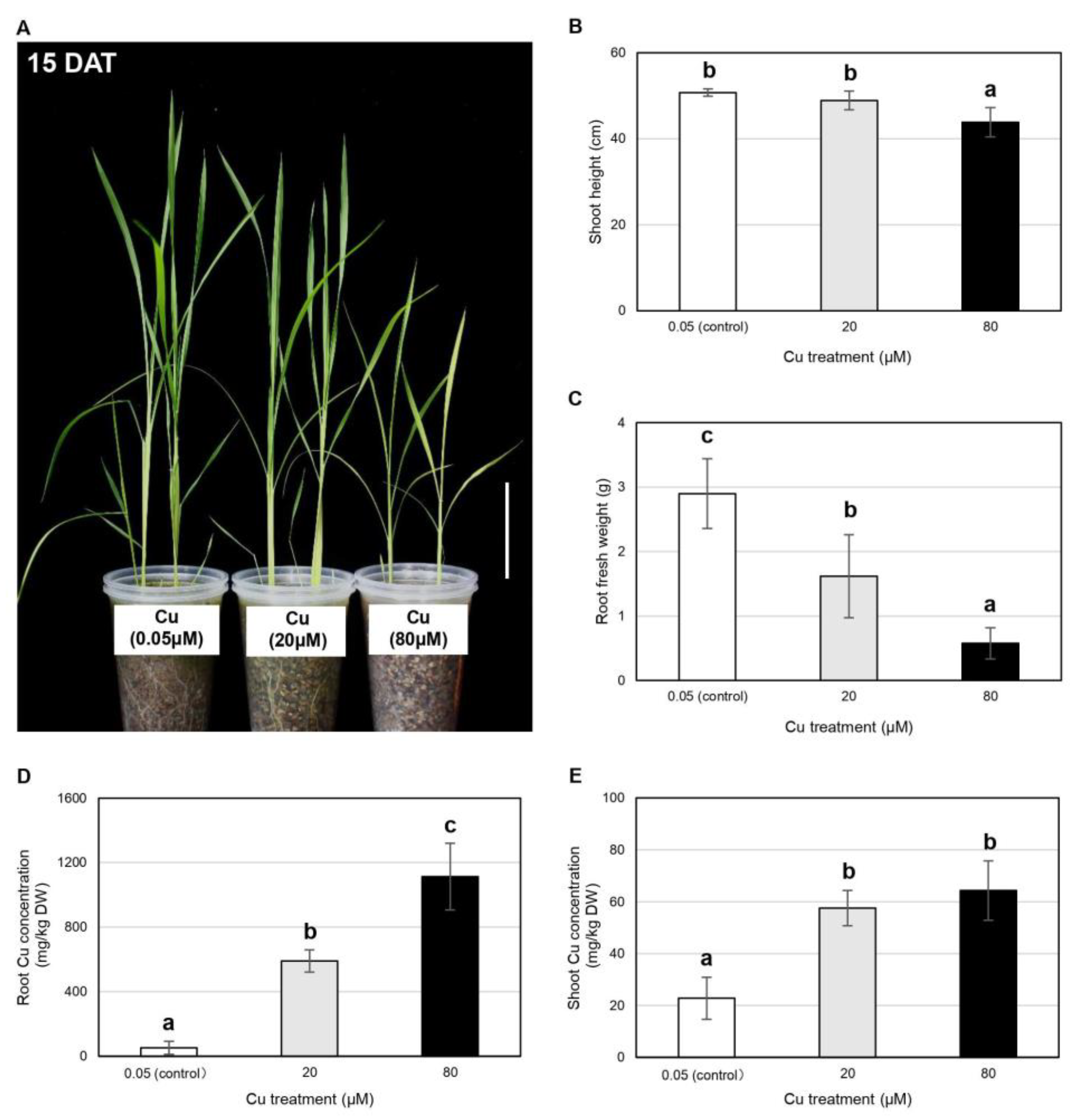
2.2. Vegetative Rice Growth Is Inhibited by Cu in a Dose-Dependent Manner but Is Not Inhibited by Fe
2.3. The Growth and Yield Components of Reproductive Rice Plants Are Unaffected by 10 μM Cu
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Insect Experiments
4.3. Rice Physiological Characteristics
4.4. Elemental Analysis
4.5. Phytohormone Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nathan, S.S.; Kalaivani, K.; Murugan, K.; Chung, P.G. Efficacy of neem limonoids on Cnaphalocrocis medinalis (Guenée) (Lepidoptera: Pyralidae) the rice leaffolder. Crop Prot. 2005, 24, 760–763. [Google Scholar] [CrossRef]
- Guo, T.W.; Liao, C.T.; Chuang, W.P. Defensive responses of rice cultivars resistant to Cnaphalocrocis medinalis (Lepidoptera: Crambidae). Arthropod Plant Interact. 2019, 13, 611–620. [Google Scholar] [CrossRef]
- Cheng, C.H.; Wu, S.C. Population dynamics and forecasting of rice leaffolder, Cnaphalocrocis medinalis (Guenée) in Taiwan. Plant Prot. Bull. Taiwan 1999, 41, 199–213. [Google Scholar]
- De Jong, P.D. Effect of folding and feeding by Cnaphalocrocis medinalis on photosynthesis and transpiration of rice leaves. Entomol. Exp. Appl. 1992, 63, 101–102. [Google Scholar] [CrossRef]
- Mahesh, P.; Srikanth, J.; Chandran, K.; Singaravelu, B.; Salin, K.P.; Jayabose, C.; Balan, S. Occurrence, damage pattern and status of the rice leaf folder Cnaphalocrocis ruralis Walker (Lepidoptera: Crambidae) in Erianthus spp. in India. Exp. Agric. 2019, 55, 471–483. [Google Scholar] [CrossRef]
- Padmavathi, C.; Katti, G.; Padmakumari, A.P.; Voleti, S.R.; Subba Rao, L.V. The effect of leaffolder Cnaphalocrocis medinalis (Guenee) [Lepidoptera: Pyralidae] injury on the plant physiology and yield loss in rice. J. Appl. Entomol. 2013, 137, 249–256. [Google Scholar] [CrossRef]
- Cheah, B.H.; Lin, H.H.; Chien, H.J.; Liao, C.T.; Liu, L.Y.D.; Lai, C.C.; Lin, Y.F.; Chuang, W.P. SWATH-MS-based quantitative proteomics reveals a uniquely intricate defense response in Cnaphalocrocis medinalis-resistant rice. Sci. Rep. 2020, 10, 6597. [Google Scholar] [CrossRef]
- Yang, Y.; Xu, H.; Lu, Y.; Wang, C.; Lu, Z. Midgut transcriptomal response of the rice leaffolder, Cnaphalocrocis medinalis (Guenée) to Cry1C toxin. PLoS ONE 2018, 13, e0191686. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Lei, W.; Wen, L.; Hou, M. Silicon-mediated resistance in a susceptible rice variety to the rice leaf folder, Cnaphalocrocis medinalis Guenée (Lepidoptera: Pyralidae). PLoS ONE 2015, 10, e0120557. [Google Scholar] [CrossRef]
- Kogan, M. Integrated pest management: Historical perspectives and contemporary developments. Annu. Rev. Entomol. 1998, 43, 243–270. [Google Scholar] [CrossRef]
- Bindraban, P.S.; Dimkpa, C.; Nagarajan, L.; Roy, A.; Rabbinge, R. Revisiting fertilisers and fertilisation strategies for improved nutrient uptake by plants. Biol. Fertil. Soils 2015, 51, 897–911. [Google Scholar] [CrossRef] [Green Version]
- Datnoff, L.E. Mineral Nutrition and Plant Disease; American Phytopathological Society: St. Paul, MN, USA, 2007; pp. 1–278. [Google Scholar]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: Cambridge, MA, USA, 2012; pp. 1–672. [Google Scholar]
- Behmer, S.T.; Lloyd, C.M.; Raubenheimer, D.; Stewart-Clark, J.; Knight, J.; Leighton, R.S.; Harper, F.A.; Smith, J.A.C. Metal hyperaccumulation in plants: Mechanisms of defence against insect herbivores. Funct. Ecol. 2005, 19, 55–66. [Google Scholar] [CrossRef]
- Boyd, R.S.; Martens, S.N. The significance of metal hyperaccumulation for biotic interactions. Chemoecology 1998, 8, 1–7. [Google Scholar] [CrossRef]
- Hörger, A.C.; Fones, H.N.; Preston, G.M. The current status of the elemental defense hypothesis in relation to pathogens. Front. Plant Sci. 2013, 4, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyd, R.S. Plant defense using toxic inorganic ions: Conceptual models of the defensive enhancement and joint effects hypotheses. Plant Sci. 2012, 195, 88–95. [Google Scholar] [CrossRef]
- Cheruiyot, D.J.; Boyd, R.S.; Moar, W.J. Exploring lower limits of plant elemental defense by cobalt, copper, nickel, and zinc. J. Chem. Ecol. 2013, 39, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Cheruiyot, D.J.; Boyd, R.S.; Moar, W. Testing the joint effects hypothesis of elemental defense using Spodoptera exigua. J. Chem. Ecol. 2015, 41, 168–177. [Google Scholar] [CrossRef]
- Liu, Z.; Sun, Z.; Zeng, C.; Dong, X.; Li, M.; Liu, Z.; Yan, M. The elemental defense effect of cadmium on Alternaria brassicicola in Brassica juncea. BMC Plant Biol. 2022, 22, 17. [Google Scholar] [CrossRef]
- Cabot, C.; Martos, S.; Llugany, M.; Gallego, B.; Tolrà, R.; Poschenrieder, C. A role for zinc in plant defense against pathogens and herbivores. Front. Plant Sci. 2019, 10, 1171. [Google Scholar] [CrossRef]
- Boyd, R. The defense hypothesis of elemental hyperaccumulation: Status, challenges and new directions. Plant Soil 2007, 293, 153–176. [Google Scholar] [CrossRef]
- Han, Y.; Li, P.; Gong, S.; Yang, L.; Wen, L.; Hou, M. Defense responses in rice induced by silicon amendment against infestation by the leaf folder Cnaphalocrocis medinalis. PLoS ONE 2016, 11, e0153918. [Google Scholar] [CrossRef] [PubMed]
- Martos, S.; Gallego, B.; Cabot, C.; Llugany, M.; Barceló, J.; Poschenrieder, C. Zinc triggers signaling mechanisms and defense responses promoting resistance to Alternaria brassicicola in Arabidopsis thaliana. Plant Sci. 2016, 249, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Winter, T.R.; Borkowski, L.; Zeier, J.; Rostás, M. Heavy metal stress can prime for herbivore-induced plant volatile emission. Plant Cell Environ. 2012, 35, 1287–1298. [Google Scholar] [CrossRef] [PubMed]
- Chmielowska, J.; Veloso, J.; Gutiérrez, J.; Silvar, C.; Díaz, J. Cross-protection of pepper plants stressed by copper against a vascular pathogen is accompanied by the induction of a defence response. Plant Sci. 2010, 178, 176–182. [Google Scholar] [CrossRef]
- Molina, A.; Hunt, M.D.; Ryals, J.A. Impaired fungicide activity in plants blocked in disease resistance signal transduction. Plant Cell 1998, 10, 1903–1914. [Google Scholar] [CrossRef]
- Cui, J.L.; Zhao, Y.P.; Lu, Y.J.; Chan, T.S.; Zhang, L.L.; Tsang, D.C.W.; Li, X.D. Distribution and speciation of copper in rice (Oryza sativa L.) from mining-impacted paddy soil: Implications for copper uptake mechanisms. Environ. Int. 2019, 126, 717–726. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Fujita, M. Salicylic acid alleviates copper toxicity in rice (Oryza sativa L.) seedlings by up-regulating antioxidative and glyoxalase systems. Ecotoxicology 2013, 22, 959–973. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Seraj, Z.I.; Fujita, M. Exogenous sodium nitroprusside and glutathione alleviate copper toxicity by reducing copper uptake and oxidative damage in rice (Oryza sativa L.) seedlings. Protoplasma 2014, 251, 1373–1386. [Google Scholar] [CrossRef]
- Xu, J.; Yang, L.; Wang, Z.; Dong, G.; Huang, J.; Wang, Y. Toxicity of copper on rice growth and accumulation of copper in rice grain in copper contaminated soil. Chemosphere 2006, 62, 602–607. [Google Scholar] [CrossRef]
- Giannousi, K.; Avramidis, I.; Dendrinou-Samara, C. Synthesis, characterization and evaluation of copper based nanoparticles as agrochemicals against Phytophthora infestans. RSC Adv. 2013, 3, 21743–21752. [Google Scholar] [CrossRef]
- Mondal, K.; Mani, C. Investigation of the antibacterial properties of nano-copper against Xanthomonas axonopodis pv. punicae, the incident of pomegranate bacterial blight. Ann. Microbiol. 2012, 62, 889–893. [Google Scholar] [CrossRef]
- Gao, Y.; Li, D.; Li, D.; Xu, P.; Mao, K.; Zhang, Y.; Qin, X.; Tang, T.; Wan, H.; Li, J.; et al. Efficacy of an adhesive nanopesticide on insect pests of rice in field trials. J. Asia-Pac. Entomol. 2020, 23, 1222–1227. [Google Scholar] [CrossRef]
- Mehraban, P.; Zadeh, A.A.; Sadeghipour, H.R. Iron toxicity in rice (Oryza sativa L.), under different potassium nutrition. Asian J. Plant Sci. 2008, 7, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Nichol, H.; Law, J.H.; Winzerling, J.J. Iron metabolism in insects. Annu. Rev. Entomol. 2002, 47, 535–559. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Zhou, B. Iron homeostasis in insects: Insights from Drosophila studies. IUBMB Life 2013, 65, 863–872. [Google Scholar] [CrossRef] [PubMed]
- Onyango, D.A.; Entila, F.; Dida, M.M.; Ismail, A.M.; Drame, K.N. Mechanistic understanding of iron toxicity tolerance in contrasting rice varieties from Africa: 1. Morpho-physiological and biochemical responses. Funct. Plant Biol. 2018, 46, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.B.; Shhadi, M.Y.; Gregorio, G.; Matthus, E.; Becker, M.; Frei, M. Genetic and physiological analysis of tolerance to acute iron toxicity in rice. Rice 2014, 7, 8. [Google Scholar] [CrossRef] [Green Version]
- Becker, M.; Asch, F. Iron toxicity in rice—Conditions and management concepts. J. Plant Nutr. Soil Sci. 2005, 168, 558–573. [Google Scholar] [CrossRef]
- Chen, L.C.; Huang, S.H.; Cheng, C.H. Review of the screening tests for rice varietal resistance to major diseases and insect pests in Taiwan. In Proceedings of the Symposium on Achievements and Perspectives of Rice Protection in Taiwan; Chia-yi Agricultural Experiment Station: Chiayi City, Taiwan, 2009; pp. 83–103. [Google Scholar]
- Yoshida, S. Routine procedure for growing rice plants in culture solution. In Laboratory Manual for Physiological Studies of Rice; Yoshida, S., Forno, D.A., Cook, J.H., Gomez, K.A., Eds.; International Rice Research Institute: Los Baños, Philippines, 1976; pp. 61–66. [Google Scholar]
- Shono, Y.; Hirano, M. Improved mass-rearing of the rice leaffolder, Cnaphalocrocis medinalis (GUENEE) (Lepidoptera: Pyralidae) using corn seedlings. Appl. Entomol. Zool. 1989, 24, 258–263. [Google Scholar] [CrossRef] [Green Version]
- Farrar, R.R.; Barbour, J.D.; Kennedy, G.G. Quantifying food consumption and growth in insects. Ann. Entomol. Soc. Am. 1989, 82, 593–598. [Google Scholar] [CrossRef]
- Waldbauer, G.P. The consumption and utilization of food by insects. In Advances in Insect Physiology; Beament, J.W.L., Treherne, J.E., Wigglesworth, V.B., Eds.; Academic Press: Cambridge, MA, USA, 1968; Volume 5, pp. 229–288. [Google Scholar]
- Matsuzaki, A.; Rutger, J.N. Characteristics relating young panicle development and heading of closely related calrose rice genotypes. Jpn. J. Crop Sci. 1991, 60, 298–305. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-F.; Liang, H.-M.; Yang, S.-Y.; Boch, A.; Clemens, S.; Chen, C.-C.; Wu, J.-F.; Huang, J.-L.; Yeh, K.-C. Arabidopsis IRT3 is a zinc-regulated and plasma membrane localized zinc/iron transporter. New Phytol. 2009, 182, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenth, R. Least-squares means: The R package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Concentration | Panicle Number per Plant | Grain Number per Panicle | Thousand Grain Weight (g) | Grain Yield (g/Plant) |
|---|---|---|---|---|---|
| Cu | 0.05 μM (control) | 2.75 ± 0.46 a | 75.14 ± 23.61 a | 28.48 ± 0.34 c | 5.89 ± 1.53 a |
| 10 μM | 2.63 ± 0.52 a | 65.90 ± 24.94 a | 29.49 ± 0.44 d | 5.10 ± 1.40 a | |
| 20 μM | 2.13 ± 0.35 a | 81.18 ± 25.33 a | 27.44 ± 0.12 b | 4.73 ± 0.77 a | |
| 30 μM | 2.25 ± 0.71 a | 76.20 ± 21.70 a | 26.39 ± 0.21 a | 4.76 ± 0.82 a | |
| Fe | 15 mg/L (control) | 3.00 ± 0.53 b | 78.88 ± 26.71 a | 27.62 ± 0.15 b | 6.49 ± 0.40 b |
| 65 mg/L | 2.50 ± 0.53 ab | 81.65 ± 30.52 a | 26.72 ± 0.26 a | 5.45 ± 0.93 ab | |
| 265 mg/L | 2.25 ± 0.46 a | 75.33 ± 29.09 a | 27.01 ± 0.24 a | 4.58 ± 1.23 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheah, B.H.; Chuang, W.-P.; Lo, J.-C.; Li, Y.; Cheng, C.-Y.; Yang, Z.-W.; Liao, C.-T.; Lin, Y.-F. Exogenous Copper Application for the Elemental Defense of Rice Plants against Rice Leaffolder (Cnaphalocrocis medinalis). Plants 2022, 11, 1104. https://doi.org/10.3390/plants11091104
Cheah BH, Chuang W-P, Lo J-C, Li Y, Cheng C-Y, Yang Z-W, Liao C-T, Lin Y-F. Exogenous Copper Application for the Elemental Defense of Rice Plants against Rice Leaffolder (Cnaphalocrocis medinalis). Plants. 2022; 11(9):1104. https://doi.org/10.3390/plants11091104
Chicago/Turabian StyleCheah, Boon Huat, Wen-Po Chuang, Jing-Chi Lo, Yi Li, Chih-Yun Cheng, Zhi-Wei Yang, Chung-Ta Liao, and Ya-Fen Lin. 2022. "Exogenous Copper Application for the Elemental Defense of Rice Plants against Rice Leaffolder (Cnaphalocrocis medinalis)" Plants 11, no. 9: 1104. https://doi.org/10.3390/plants11091104
APA StyleCheah, B. H., Chuang, W. -P., Lo, J. -C., Li, Y., Cheng, C. -Y., Yang, Z. -W., Liao, C. -T., & Lin, Y. -F. (2022). Exogenous Copper Application for the Elemental Defense of Rice Plants against Rice Leaffolder (Cnaphalocrocis medinalis). Plants, 11(9), 1104. https://doi.org/10.3390/plants11091104