
Biopesticide Activity of Guayule Resin

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Guayule Efficacy against T. urticae
2.2. Guayule Efficacy against B. tabaci
2.3. Guayule Efficacy against M. persicae
2.4. Guayule Efficacy against F. occidentalis
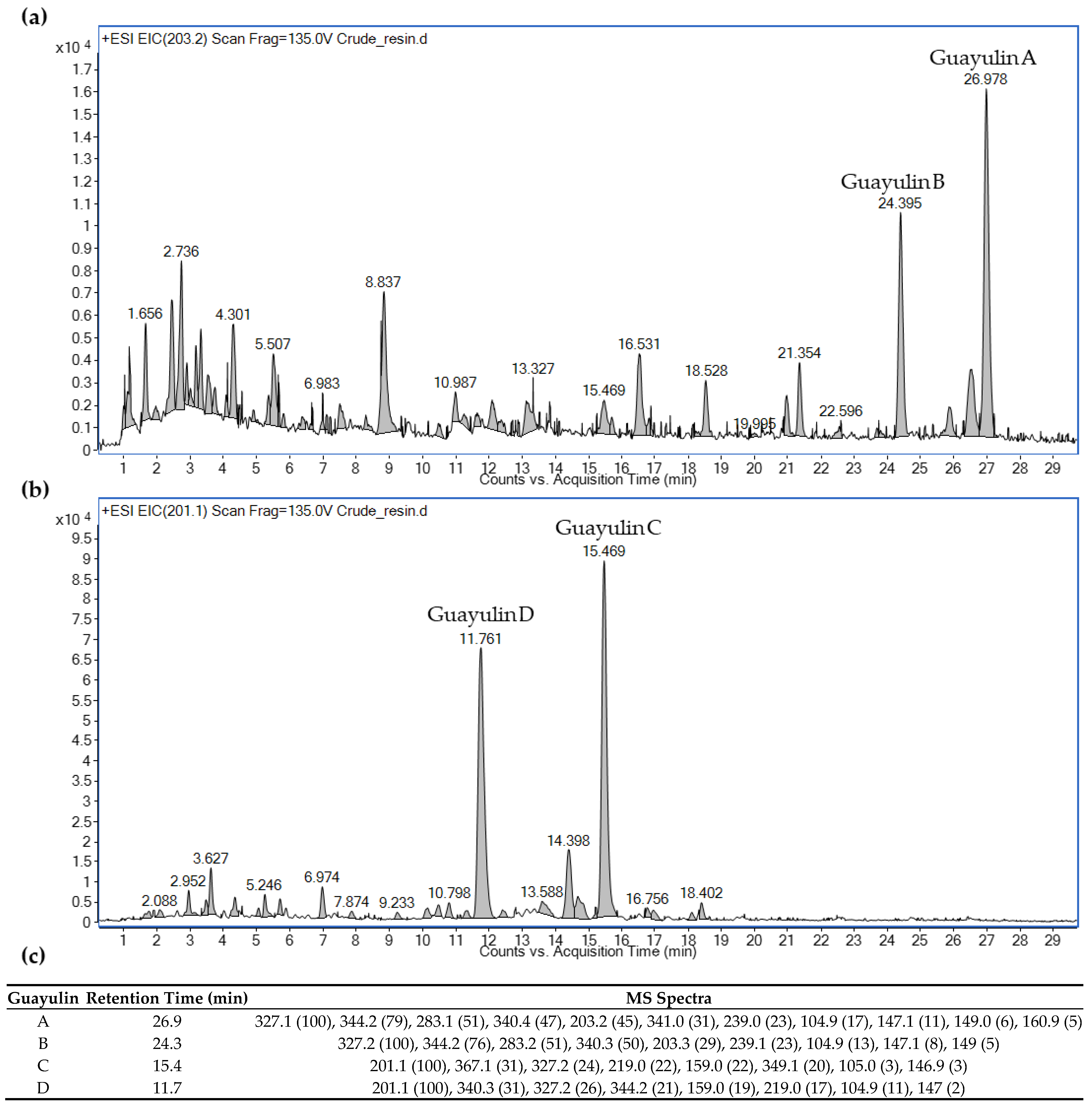
2.5. Characterization of Guayule Extracts
3. Discussion
4. Materials and Methods
4.1. Crude Resin and Fractionation
4.2. Insect Populations and Rearing Conditions
4.3. Test Items and Treatments
4.4. Bioassays
4.4.1. Contact Mortality (Leaf Disc Method)
4.4.2. Antifeedant Effects (Feeding Deterrent)
4.5. Chemical Profile Using Hplc-MS/MS
4.6. Correlation of Activity and Compounds
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, D. Advances in plant biopesticides. In Advances in Plant Biopesticides; Springer: Berlin/Heidelberg, Germany, 2014; pp. 323–345. ISBN 9788132220060. [Google Scholar]
- Campos, E.V.R.; Proença, P.L.F.; Oliveira, J.L.; Bakshi, M.; Abhilash, P.C.; Fraceto, L.F. Use of botanical insecticides for sustainable agriculture: Future perspectives. Ecol. Indic. 2019, 105, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Damalas, C.A.; Koutroubas, S.D. Current status and recent developments in biopesticide use. Agriculture 2018, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Kumar, J.; Ramlal, A.; Mallick, D.; Mishra, V. An overview of some biopesticides and their importance in plant protection for commercial acceptance. Plants 2021, 10, 1185. [Google Scholar] [CrossRef] [PubMed]
- FAO. New Standards to Curb the Global Spread of Plant Pests and Diseases. Available online: https://www.fao.org/news/story/en/item/1187738/icode/ (accessed on 29 April 2021).
- EU Pesticides Database. Available online: https://ec.europa.eu/food/plant/pesticides/eu-pesticides-database/active-substances/?event=search.as (accessed on 30 April 2021).
- Cheng, F.; Dehghanizadeh, M.; Audu, M.A.; Jarvis, J.M.; Holguin, F.O.; Brewer, C.E. Characterization and evaluation of guayule processing residues as potential feedstock for biofuel and chemical production. Ind. Crops Prod. 2020, 150, 112311. [Google Scholar] [CrossRef]
- Gutiérrez, C.; Gonzalez-Coloma, A.; Hoffmann, J.J. Antifeedant properties of natural products from Parthenium argentatum, P. argentatum × P. tomentosum (Asteraceae) and Castela emoryi (Simaroubeaceae) against Reticulitermes flavipes. Ind. Crops Prod. 1999, 10, 35–40. [Google Scholar] [CrossRef]
- Holt, G.A.; Chow, P.; Wanjura, J.D.; Pelletier, M.G.; Coffelt, T.A.; Nakayama, F.S. Termite resistance of biobased composition boards made from cotton byproducts and guayule bagasse. Ind. Crops Prod. 2012, 36, 508–512. [Google Scholar] [CrossRef]
- Nakayama, F.S.; Vinyard, S.H.; Chow, P.; Bajwa, D.S.; Youngquist, J.A.; Muehl, J.H.; Krzysik, A.M. Guayule as a wood preservative. Ind. Crops Prod. 2001, 14, 105–111. [Google Scholar] [CrossRef]
- Bultman, J.D.; Gilbertson, R.L.; Adaskaveg, J.; Amburgey, T.L.; Parikh, S.V.; Bailey, C.A. The efficacy of guayule resin as a pesticide. Bioresour. Technol. 1991, 35, 197–201. [Google Scholar] [CrossRef]
- Isman, M.B.; Rodriguez, E. Larval growth inhibitors from species of Parthenium (Asteraceae). Phytochemistry 1983, 22, 2709–2713. [Google Scholar] [CrossRef]
- Zhang, Z. Mites of Greenhouses Identification, Biology and Control; CABI Publishing: Wallingford/Oxfordshire, UK, 2003; ISBN 085199590X. [Google Scholar]
- Roh, H.S.; Lim, E.G.; Kim, J.; Park, C.G. Acaricidal and oviposition deterring effects of santalol identified in sandalwood oil against two-spotted spider mite Tetranychus urticae Koch (Acari: Tetranychidae). J. Pest Sci. 2011, 84, 495–501. [Google Scholar] [CrossRef] [Green Version]
- Centre for Agricultural Bioscience International Invasive Species Compendium: Tetranychus urticae (Two-Spotted Spider Mite). Available online: https://www.cabi.org/isc/datasheet/53366 (accessed on 2 September 2021).
- Insecticide Resistance Action Committee Arthropod Pesticide Resistance Database. Available online: https://www.pesticideresistance.org/search.php (accessed on 2 September 2021).
- Oliveira, M.R.V.; Henneberry, T.J.; Anderson, P. History, current status, and collaborative research projects for Bemisia tabaci. Crop Prot. 2001, 20, 709–723. [Google Scholar] [CrossRef] [Green Version]
- Centre for Agricultural Bioscience International Invasive Species Compendium: Bemisia tabaci (Tobacco Whitefly). Available online: https://www.cabi.org/isc/datasheet/8927 (accessed on 11 January 2022).
- Bielza, P.; Moreno, I.; Belando, A.; Grávalos, C.; Izquierdo, J.; Nauen, R. Spiromesifen and spirotetramat resistance in field populations of Bemisia tabaci Gennadius in Spain. Pest Manag. Sci. 2019, 75, 45–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centre for Agricultural Bioscience International Invasive Species Compendium: Myzus persicae (Green Peach Aphid). Available online: https://www.cabi.org/isc/datasheet/35642 (accessed on 2 September 2021).
- Bass, C.; Puinean, A.M.; Zimmer, C.T.; Denholm, I.; Field, L.M.; Foster, S.P.; Gutbrod, O.; Nauen, R.; Slater, R.; Williamson, M.S. The evolution of insecticide resistance in the peach potato aphid, Myzus persicae. Insect Biochem. Mol. Biol. 2014, 51, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centre for Agricultural Bioscience International Invasive Species Compendium: Frankliniella occidentalis (Western Flower Thrips). Available online: https://www.cabi.org/isc/datasheet/24426 (accessed on 2 September 2021).
- EPPO. EPPO Standards—EPPO A1 and A2 Lists of Pests Recommended for Regulation as Quarantine Pests; European and Mediterranean Plant Protection Organization: Paris, France, 2021; Volume 2. [Google Scholar]
- Demirozer, O.; Tyler-Julian, K.; Funderburk, J.; Leppla, N.; Reitz, S. Frankliniella occidentalis (Pergande) integrated pest management programs for fruiting vegetables in Florida. Pest Manag. Sci. 2012, 68, 1537–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozalén, J.; García-Martínez, M.M.; Zalacain, A.; López-Córcoles, H.; Hurtado de Mendoza, J.; Cornish, K.; Carmona, M. Future trends for the analysis of guayulins in guayule (Parthenium argentatum Gray) resins. Ind. Crops Prod. 2021, 159, 113027. [Google Scholar] [CrossRef]
- Dehghanizadeh, M.; Mendoza Moreno, P.; Sproul, E.; Bayat, H.; Quinn, J.C.; Brewer, C.E. Guayule (Parthenium argentatum) resin: A review of chemistry, extraction techniques, and applications. Ind. Crops Prod. 2021, 165, 113410. [Google Scholar] [CrossRef]
- Xu, Y.M.; Madasu, C.; Liu, M.X.; Wijeratne, E.M.K.; Dierig, D.; White, B.; Molnár, I.; Gunatilaka, A.A.L. Cycloartane- and Lanostane-Type Triterpenoids from the Resin of Parthenium argentatum AZ-2, a Byproduct of Guayule Rubber Production. ACS Omega 2021, 6, 15486–15498. [Google Scholar] [CrossRef]
- Romo de Vivar, A.; Martínez-Vázquez, M.; Matsubara, C.; Pérez-Sánchez, G.; Joseph-Nathan, P. Triterpenes in Parthenium argentatum, structures of argentatins C and D. Phytochemistry 1990, 29, 915–918. [Google Scholar] [CrossRef]
- Maia, M.F.; Moore, S.J. Plant-based insect repellents: A review of their efficacy, development and testing. Malar. J. 2011, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Miresmailli, S.; Isman, M.B. Botanical insecticides inspired by plant-herbivore chemical interactions. Trends Plant Sci. 2014, 19, 29–35. [Google Scholar] [CrossRef]
- Jara, F.M.; Cornish, K.; Carmona, M. Potential applications of guayulins to improve feasibility of guayule cultivation. Agronomy 2019, 9, 804. [Google Scholar] [CrossRef] [Green Version]
- Bailen, M.; Martínez-Díaz, R.A.; Hoffmann, J.J.; Gonzalez-Coloma, A. Molecular Diversity from Arid-Land Plants: Valorization of Terpenes and Biotransformation Products. Chem. Biodivers. 2020, 17, e1900663. [Google Scholar] [CrossRef] [PubMed]
- Isman, M. Insect antifeedants. Pestic. Outlook 2002, 13, 152–157. [Google Scholar] [CrossRef]
- Ruiz-Vásquez, L.; Reina, M.; Fajardo, V.; López, M.; González-Coloma, A. Insect Antifeedant Components of Senecio fistulosus var. fistulosus—Hualtata. Plants 2019, 8, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rimando, A.M.; Duke, S.O. Natural products for pest management. ACS Symp. Ser. 2006, 927, 2–21. [Google Scholar] [CrossRef] [Green Version]
- El-Wakeil, N.E. Botanical Pesticides and Their Mode of Action. Gesunde Pflanz. 2013, 65, 125–149. [Google Scholar] [CrossRef]
- Abbot, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Dose (cc/L) | Contact Mortality (%M) 1 | Settling Inhibition (%SI) | Egg Laying Inhibition (%IO) |
|---|---|---|---|---|
| Crude guayule resin | 1% (w/v) | 26.8 ± 10.4 d | 97.1 ± 2.1 * | 96.3 ± 4.3 * |
| Hexane fraction | 1% (w/v) | 83.1 ± 5.9 ab | 85.2 ± 2.2 * | 81.8 ± 5.3 * |
| Ethyl acetate fraction | 1% (w/v) | 8.8 ± 3.5 e | 70.4 ± 7.2 * | 68.6 ± 10.1 * |
| Methanol fraction | 1% (w/v) | 53.1 ± 19.4 c | 28.5 ± 10.2 | 51.4 ± 8.5 |
| SPINTOR® 480 SC (Spinosad) | 0.7 | 21.5 ± 2.0 d | 0.0 ± 0.0 | 45.1 ± 3.3 |
| K_Prototype-1 | 4 | 100.0 ± 0.0 a | 77.3 ± 6.5 * | 78.4 ± 5.3 * |
| K_Prototype-2 | 4 | 100.0 ± 0.0 a | 91.4 ± 2.3 * | 93.6 ± 2.1 * |
| K_Prototype-3 | 4 | 76.3 ± 8.4 b | 92.1 ± 3.3 * | 91.9 ± 4.1 * |
| K_Prototype-4 | 4 | 92.6 ± 4.0 ab | 98.2 ± 1.7 * | 99.0 ± 0.8 * |
| Treatment | Dose (cc/L) | Contact Mortality (%M) 1 | Settling Inhibition (%SI) | Egg Laying Inhibition (%IO) |
|---|---|---|---|---|
| Crude guayule resin | 1% (w/v) | 64.8 ± 11.5 c | 77.5 ± 6.4 * | 86.3 ± 2.7 * |
| Hexane fraction | 1% (w/v) | 70.7 ± 7.5 b | 47.7 ± 9.1 | 45.4 ± 8.3 |
| Ethyl acetate fraction | 1% (w/v) | 43.5 ± 8.7 d | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Methanol fraction | 1% (w/v) | 38.7 ± 7.6 d | 36.8 ± 9.5 | 45.9 ± 7.7 |
| SPINTOR® 480 SC (Spinosad) | 0.7 | 30.8 ± 3.3 e | 0.0 ± 0.0 | 60.5 ± 7.2 * |
| K_Prototype-1 | 4 | 100.0 ± 0.0 a | 77.4 ± 4.2 * | 85.2 ± 3.3 * |
| K_Prototype-2 | 4 | 100.0 ± 0.0 a | 89.3 ± 4.6 * | 98.6 ± 1.1 * |
| K_Prototype-3 | 4 | 98.7 ± 0.9 a | 93.9 ± 2.1 * | 94.5 ± 3.1 * |
| K_Prototype-4 | 4 | 98.3 ± 1.7 a | 30.4 ± 6.2 | 68.3 ± 5.2 * |
| Treatment | Dose (cc/L) | Contact Mortality (%M) | Settling Inhibition (%SI) |
|---|---|---|---|
| Crude guayule resin | 1% (w/v) | 14.8 ± 7.5 c | 15.8 ± 7.9 |
| Hexane fraction | 1% (w/v) | 49.1 ± 10.5 b | 72.7 ± 6.6 * |
| Ethyl acetate fraction | 1% (w/v) | 8.8. ± 3.5 c | 70.4 ± 5.6 * |
| Methanol fraction | 1% (w/v) | 53.1 ± 19.5 b | 51.4 ± 6.6 |
| SPINTOR® 480 SC (Spinosad) | 0.7 | 24.5 ± 6.4 c | 45.0 ± 8.1 |
| K_Prototype-1 | 4 | 100.0 ± 0.0 a | 73.2 ± 10.2 * |
| K_Prototype-2 | 4 | 100.0 ± 0.0 a | 67.7 ± 11.1 * |
| K_Prototype-3 | 4 | 88.1 ± 4.8 a | 62.1 ± 9.8 * |
| K_Prototype-4 | 4 | 97.9 ± 2.0 a | 61.4 ± 8.9 * |
| Treatment | Dose (cc/L) | Contact Mortality (%M) 1 | Settling Inhibition (%SI) | Feeding Reduction (%FR) |
|---|---|---|---|---|
| Crude guayule resin | 1% (w/v) | 31.8 ± 10.9 c | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Hexane fraction | 1% (w/v) | 37.1 ± 7.7 c | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Ethyl acetate fraction | 1% (w/v) | 20.6 ± 9.1 d | 0.0 ± 0.0 | 20.5 ± 11.2 |
| Methanol fraction | 1% (w/v) | 7.3 ± 7.2 d | 0.0 ± 0.0 | 0.0 ± 0.0 |
| SPINTOR® 480 SC (Spinosad) | 0.7 | 59.8 ± 7.1 b | 51.4 ± 7.3 * | 49.3 ± 4.2 |
| K_Prototype-1 | 4 | 97.5 ± 2.5 a | 62.1 ± 9.3 * | 82.1 ± 5.2 * |
| K_Prototype-2 | 4 | 97.8 ± 2.1 a | 0.0 ± 0.0 | 86.3 ± 7.3 * |
| K_Prototype-3 | 4 | 60.8 ± 3.7 b | 44.4 ± 10.2 | 42.67 ± 9.1 |
| K_Prototype-4 | 4 | 92.1 ± 4.8 a | 0.0 ± 0.0 | 60.91 ± 6.3 * |
| %M 1 | %SI | %IO | %FR | C1 | C2 | aC | C4 | gD | gC | isoaA | aA | C6 | aD | gB | isoaB | aB | gA | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T. urticae | %M | PC | 1 | 0.293 | 0.206 | −0.899 | −1.000 ** | −0.994 | −0.891 | 0.973 | 0.970 | 0.995 | 0.998 * | 0.985 | 0.994 | −0.389 | 0.976 | 0.972 | −0.353 | |
| Sig. | 0.811 | 0.868 | 0.288 | 0.008 | 0.068 | 0.300 | 0.149 | 0.155 | 0.062 | 0.039 | 0.110 | 0.067 | 0.745 | 0.140 | 0.151 | 0.770 | ||||
| %SI | PC | 1 | 0.996 | 0.155 | −0.281 | −0.189 | 0.173 | 0.062 | 0.053 | 0.198 | 0.350 | 0.124 | 0.190 | 0.767 | 0.077 | 0.060 | 0.791 | |||
| Sig. | 0.057 | 0.901 | 0.819 | 0.879 | 0.889 | 0.960 | 0.966 | 0.873 | 0.772 | 0.921 | 0.878 | 0.444 | 0.951 | 0.962 | 0.419 | |||||
| %IO | PC | 1 | 0.243 | −0.194 | −0.100 | 0.260 | −0.027 | −0.036 | 0.109 | 0.265 | 0.035 | 0.101 | 0.821 | −0.012 | −0.030 | 0.843 | ||||
| Sig. | 0.844 | 0.876 | 0.936 | 0.832 | 0.983 | 0.977 | 0.930 | 0.829 | 0.978 | 0.935 | 0.387 | 0.992 | 0.981 | 0.362 | ||||||
| B. tabaci | %M | PC | 1 | 0.826 | 0.764 | −0.491 | −0.816 | −0.757 | −0.475 | 0.668 | 0.661 | 0.763 | 0.855 | 0.713 | 0.758 | 0.204 | 0.679 | 0.666 | 0.242 | |
| Sig. | 0.381 | 0.446 | 0.673 | 0.393 | 0.454 | 0.685 | 0.535 | 0.540 | 0.448 | 0.347 | 0.495 | 0.452 | 0.869 | 0.525 | 0.536 | 0.845 | ||||
| %SI | PC | 1 | 0.995 | 0.086 | −0.348 | −0.257 | 0.104 | 0.132 | 0.123 | 0.266 | 0.415 | 0.193 | 0.258 | 0.720 | 0.147 | 0.129 | 0.746 | |||
| Sig. | 0.065 | 0.945 | 0.774 | 0.835 | 0.934 | 0.916 | 0.922 | 0.829 | 0.728 | 0.876 | 0.834 | 0.488 | 0.906 | 0.917 | 0.463 | |||||
| %IO | PC | 1 | 0.187 | −0.250 | −0.156 | 0.205 | 0.030 | 0.021 | 0.166 | 0.320 | 0.092 | 0.158 | 0.787 | 0.045 | 0.027 | 0.811 | ||||
| Sig. | 0.880 | 0.839 | 0.900 | 0.869 | 0.981 | 0.987 | 0.894 | 0.793 | 0.941 | 0.899 | 0.423 | 0.971 | 0.983 | 0.398 | ||||||
| M. persicae | %M | PC | 1 | 0.409 | −0.937 | −0.996 | −1.000 ** | −0.931 | 0.990 | 0.989 | 1.000 ** | 0.988 | 0.997 * | 1.000 ** | −0.476 | 0.992 | 0.990 | −0.441 | ||
| Sig. | 0.732 | 0.227 | 0.053 | 0.007 | 0.239 | 0.088 | 0.094 | 0.001 | 0.100 | 0.049 | 0.006 | 0.684 | 0.079 | 0.090 | 0.709 | |||||
| %SI | PC | 1 | −0.702 | −0.331 | −0.419 | −0.714 | 0.531 | 0.538 | 0.410 | 0.261 | 0.477 | 0.417 | −0.997 * | 0.518 | 0.533 | −0.999 * | ||||
| Sig. | 0.505 | 0.786 | 0.725 | 0.493 | 0.644 | 0.638 | 0.731 | 0.832 | 0.683 | 0.726 | 0.048 | 0.653 | 0.642 | 0.023 | ||||||
| F. occiden. | %M | PC | 1 | −0.949 | −0.585 | −0.875 | −0.825 | −0.571 | 0.747 | 0.741 | 0.831 | 0.908 | 0.787 | 0.826 | 0.093 | 0.757 | 0.745 | 0.131 | ||
| Sig. | 0.204 | 0.602 | 0.321 | 0.382 | 0.613 | 0.463 | 0.469 | 0.376 | 0.275 | 0.423 | 0.381 | 0.941 | 0.453 | 0.465 | 0.916 | |||||
| %FR | PC | 1 | 0.301 | 0.679 | 0.606 | 0.283 | −0.500 | −0.492 | −0.613 | −0.730 | −0.553 | −0.607 | −0.401 | −0.513 | −0.497 | −0.437 | ||||
| Sig. | 0.806 | 0.525 | 0.586 | 0.817 | 0.667 | 0.673 | 0.580 | 0.479 | 0.627 | 0.585 | 0.737 | 0.657 | 0.669 | 0.712 |
| Name | Dose | Target | Batch code | Type |
|---|---|---|---|---|
| Crude guayule resin | 1% (w/v) | Unknown | GR-0720-CR | Candidate |
| Hexane fraction | 1% (w/v) | Unknown | GR-0720-H | Candidate |
| Ethyl acetate fraction | 1% (w/v) | Unknown | GR-0720-EA | Candidate |
| Methanol fraction | 1% (w/v) | Unknown | GR-0720-M | Candidate |
| SPINTOR® 480 SC | 0.7 cc/L | Broad-spectrum | N/A | End-use product (EP) |
| K_Prototype-1 | 4 cc/L | Broad-spectrum | K1-2020-1 | Prototype |
| K_Prototype-2 | 4 cc/L | Broad-spectrum | K2-2020-1 | Prototype |
| K_Prototype-3 | 4 cc/L | B. tabaci (main), side pests | K3-2020-0021 | End-use product (EP) |
| K_Prototype-4 | 4 cc/L | T. urticae (main), side pests | K4-2020-0003 | End-use product (EP) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Latorre, G.; García-Martínez, M.M.; Martín-Bejerano, M.; Julio, L.F.; Zalacain, A.; Carrión, M.E.; Carmona, M. Biopesticide Activity of Guayule Resin. Plants 2022, 11, 1169. https://doi.org/10.3390/plants11091169
Latorre G, García-Martínez MM, Martín-Bejerano M, Julio LF, Zalacain A, Carrión ME, Carmona M. Biopesticide Activity of Guayule Resin. Plants. 2022; 11(9):1169. https://doi.org/10.3390/plants11091169
Chicago/Turabian StyleLatorre, Guayente, María Mercedes García-Martínez, María Martín-Bejerano, Luis F. Julio, Amaya Zalacain, María Engracia Carrión, and Manuel Carmona. 2022. "Biopesticide Activity of Guayule Resin" Plants 11, no. 9: 1169. https://doi.org/10.3390/plants11091169
APA StyleLatorre, G., García-Martínez, M. M., Martín-Bejerano, M., Julio, L. F., Zalacain, A., Carrión, M. E., & Carmona, M. (2022). Biopesticide Activity of Guayule Resin. Plants, 11(9), 1169. https://doi.org/10.3390/plants11091169