
Evaluating Seagrass Meadow Dynamics by Integrating Field-Based and Remote Sensing Techniques

 ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. The Field-Based Data Perspective
2.2. Pre-Image Classification Procedures
2.3. Supervised Image Classification and Its Validation
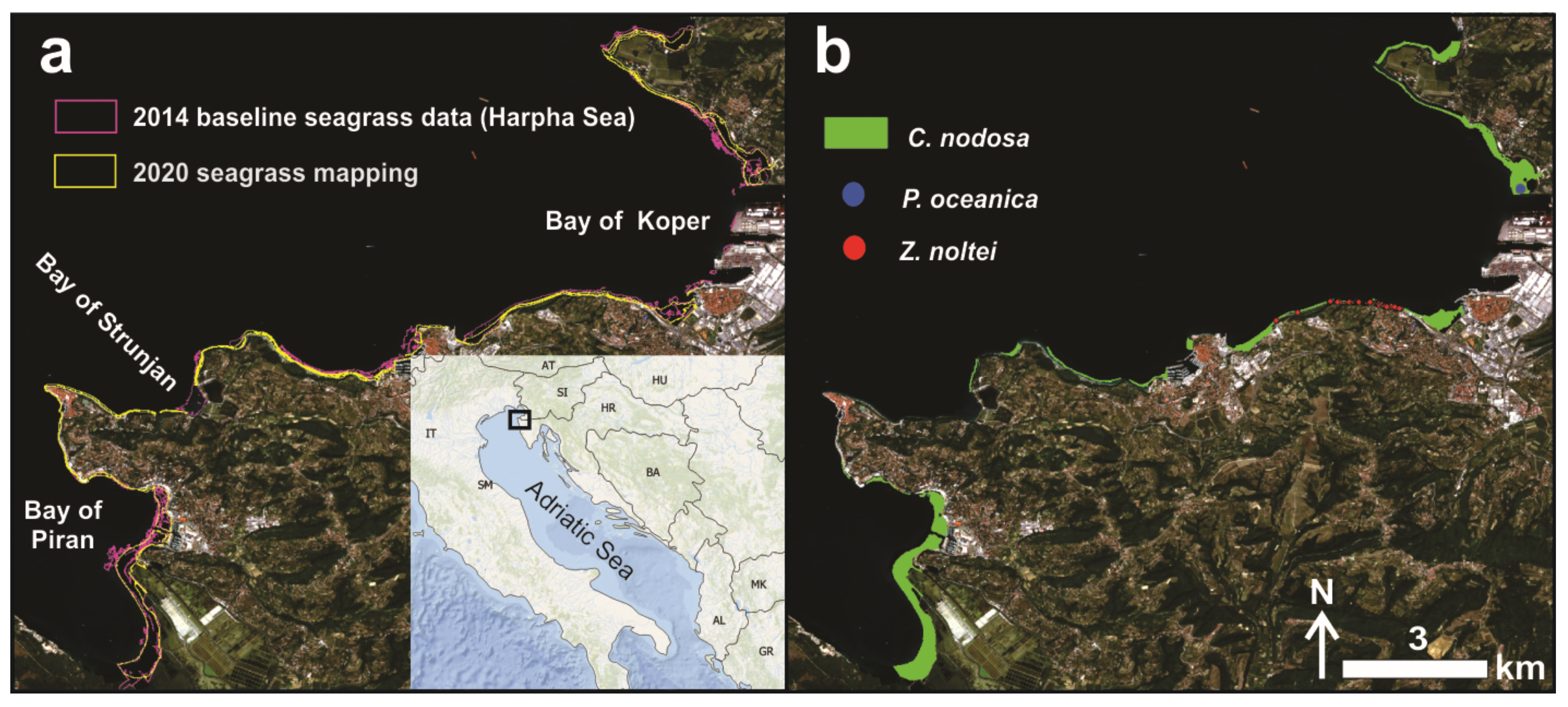
2.4. Seagrass Meadow Dynamics
3. Discussion
4. Materials and Methods
4.1. Study Area
4.2. Seagrass Distribution Data
4.2.1. In Situ Data and Mapping
4.2.2. Satellite Data
4.2.3. Auxiliary Data
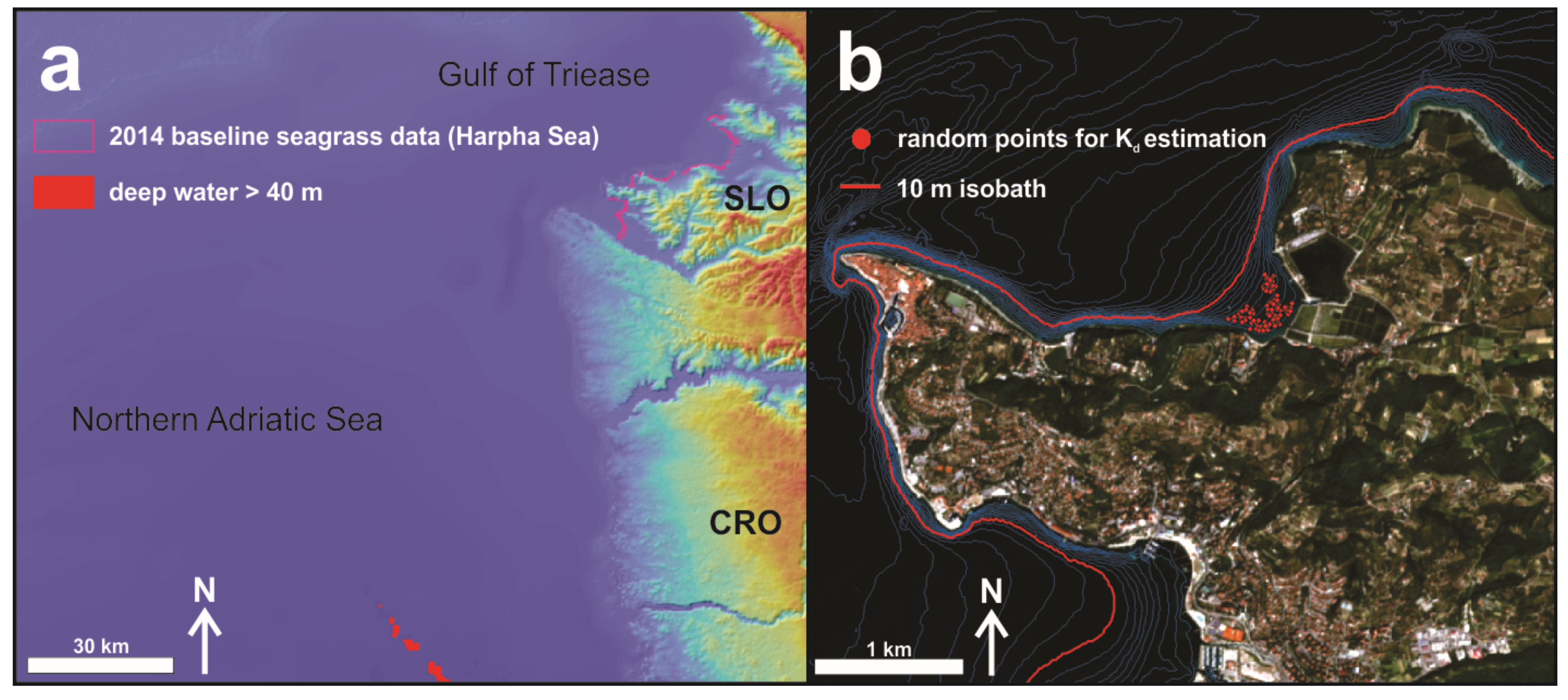
4.3. Water Column Correction
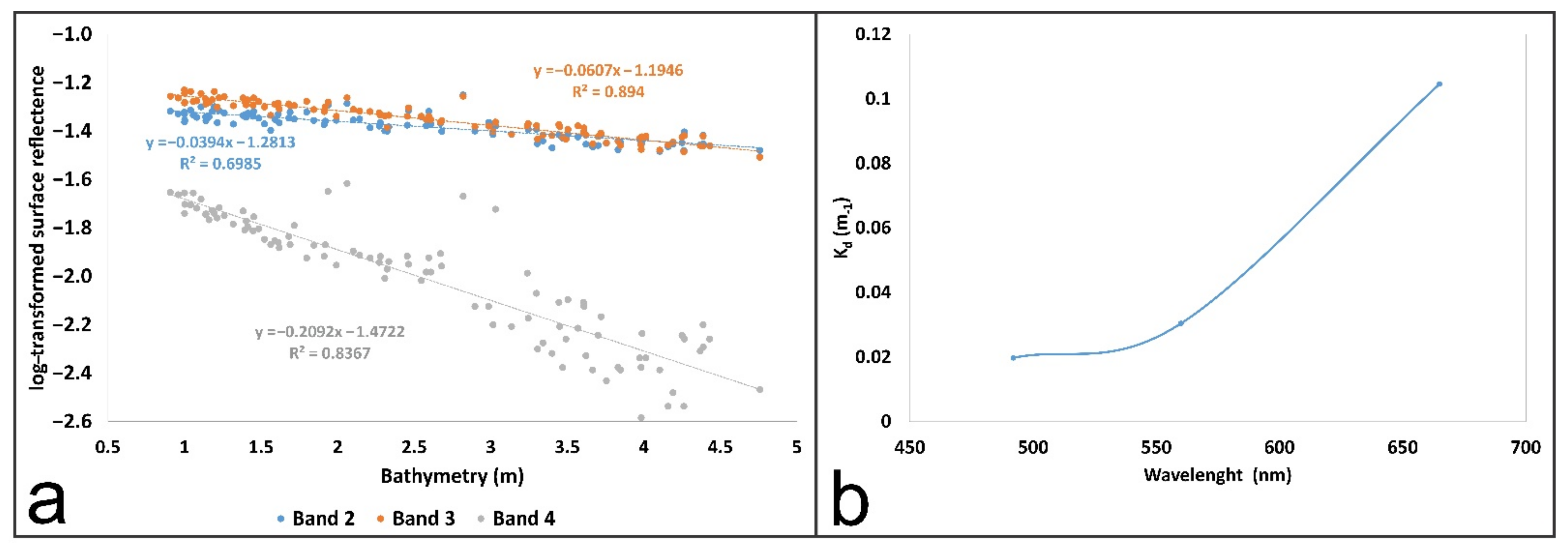
4.3.1. Infinitely Deep-Water Reflectance Estimation
4.3.2. Diffuse Attenuation Coefficient Estimation
4.4. Image Classification
4.5. Accuracy Assessment
4.6. Change Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Costanza, R.; d’Arge, R.; de Groot, R.; Farber, S.; Grasso, M.; Hannon, B.; Limburg, K.; Naeem, S.; O’Neill, R.V.; Paruelo, J.; et al. The Value of the World’s Ecosystem Services and Natural Capital. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- Spalding, M.; Taylor, M.; Ravilious, C.; Short, F.; Green, E. The Distribution and Status of Seagrasses. In World Atlas of Seagrasses; University of California Press: Berkeley, CA, USA, 2003; pp. 5–26. ISBN 0-520-24047-2. [Google Scholar]
- Brodersen, M.M.; Pantazi, M.; Kokkali, A.; Panayotidis, P.; Gerakaris, V.; Maina, I.; Kavadas, S.; Kaberi, H.; Vassilopoulou, V. Cumulative Impacts from Multiple Human Activities on Seagrass Meadows in Eastern Mediterranean Waters: The Case of Saronikos Gulf (Aegean Sea, Greece). Environ. Sci. Pollut. Res. 2018, 25, 26809–26822. [Google Scholar] [CrossRef] [PubMed]
- Short, F.T. Loss and restoration of seagrass ecosystems. In Proceedings of the 5th International Conference on Environmental Future, Zurich, Switzerland, 23–27 March 2003; pp. 23–27. [Google Scholar]
- Boudouresque, C.F.; Bernard, G.; Pergent, G.; Shili, A.; Verlaque, M. Regression of Mediterranean Seagrasses Caused by Natural Processes and Anthropogenic Disturbances and Stress: A Critical Review. Bot. Mar. 2009, 52, 395–418. [Google Scholar] [CrossRef]
- Telesca, L.; Belluscio, A.; Criscoli, A.; Ardizzone, G.; Apostolaki, E.T.; Fraschetti, S.; Gristina, M.; Knittweis, L.; Martin, C.S.; Pergent, G.; et al. Seagrass Meadows (Posidonia oceanica) Distribution and Trajectories of Change. Sci. Rep. 2015, 5, 12505. [Google Scholar] [CrossRef] [Green Version]
- Hughes, A.R.; Williams, S.L.; Duarte, C.M.; Heck, K.L.; Waycott, M. Associations of Concern: Declining Seagrasses and Threatened Dependent Species. Front. Ecol. Environ. 2009, 7, 242–246. [Google Scholar] [CrossRef] [Green Version]
- Waycott, M.; Duarte, C.M.; Carruthers, T.J.B.; Orth, R.J.; Dennison, W.C.; Olyarnik, S.; Calladine, A.; Fourqurean, J.W.; Heck, K.L.; Hughes, A.R.; et al. Accelerating Loss of Seagrasses across the Globe Threatens Coastal Ecosystems. Proc. Natl. Acad. Sci. USA 2009, 106, 12377–12381. [Google Scholar] [CrossRef] [Green Version]
- de los Santos, C.B.; Krause-Jensen, D.; Alcoverro, T.; Marbà, N.; Duarte, C.M.; van Katwijk, M.M.; Pérez, M.; Romero, J.; Sánchez-Lizaso, J.L.; Roca, G.; et al. Recent Trend Reversal for Declining European Seagrass Meadows. Nat. Commun. 2019, 10, 3356. [Google Scholar] [CrossRef] [Green Version]
- Palinkas, C.; Mistri, M.; Staver, L.; Lipej, L.; Kružić, P.; Stevenson, J.C.; Tamburri, M.; Munari, C.; Orlando-Bonaca, M. Status of critical habitats and invasive species. In Coastal Ecosystems in Transition: A Comparative Analysis of the Northern Adriatic and Chesapeake Bay; Malone, T.C., Malej, A., Faganeli, J., Eds.; Geophysical Monograph Series; Wiley & Sons: New York, NY, USA, 2020; pp. 177–202. ISBN 978-1-119-54358-9. [Google Scholar]
- Beck, M.W.; Heck, K.L.; Able, K.W.; Childers, D.L.; Eggleston, D.B.; Gillanders, B.M.; Halpern, B.; Hays, C.G.; Hoshino, K.; Minello, T.J.; et al. The Identification, Conservation, and Management of Estuarine and Marine Nurseries for Fish and Invertebrates. BioScience 2001, 51, 633. [Google Scholar] [CrossRef]
- Tuya, F.; Pérez, J.; Medina, L.; Luque, A. Seasonal Variation of the Macrofauna from Three Seagrass Meadows of Cymodocea nodosa of Gran Canaria (Central-Eastern Atlantic Ocean). Cienc. Mar. 2001, 27, 223–234. [Google Scholar] [CrossRef] [Green Version]
- Espino, F.; Tuya, F.; Brito, A.; Haroun, R. Ichthyofauna Associated with Cymodocea nodosa Meadows in the Canarian Archipelago (Central Eastern Atlantic): Community Structure and Nursery Role. Cienc. Mar. 2011, 37, 157–174. [Google Scholar] [CrossRef] [Green Version]
- Cabaço, S.; Santos, R.; Duarte, C.M. The Impact of Sediment Burial and Erosion on Seagrasses: A Review. Estuar. Coast. Shelf Sci. 2008, 79, 354–366. [Google Scholar] [CrossRef]
- Cabaco, S.; Santos, R. Human-Induced Changes of the Seagrass Cymodocea nodosa in Ria Formosa Lagoon (Southern Portugal) after a Decade. Cah. Biol. Mar. 2014, 55, 101–108. [Google Scholar]
- Terrados, J.; Borum, J. Why Are Seagrasses Important? Goods and Services Provided by Seagrass Meadows. In European Seagrasses: An Introduction to Monitoring and Management; M&MS Project: CA, USA, 2004; pp. 8–10. ISBN 87-89143-21-3. [Google Scholar]
- Peduzzi, P.; Vukovič, A. Primary Production of Cymodocea nodosa in the Gulf of Trieste (Northern Adriatic Sea): A Comparison of Methods. Mar. Ecol. Prog. Ser. 1990, 64, 197–207. [Google Scholar] [CrossRef]
- Duarte, C.M.; Marbà, N.; Gacia, E.; Fourqurean, J.W.; Beggins, J.; Barrón, C.; Apostolaki, E.T. Seagrass Community Metabolism: Assessing the Carbon Sink Capacity of Seagrass Meadows: SEAGRASS COMMUNITY METABOLISM. Glob. Biogeochem. Cycles 2010, 24, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Burkholder, J.M.; Tomasko, D.A.; Touchette, B.W. Seagrasses and Eutrophication. J. Exp. Mar. Biol. Ecol. 2007, 350, 46–72. [Google Scholar] [CrossRef]
- Eklöf, J.S.; de la Torre-Castro, M.; Gullström, M.; Uku, J.; Muthiga, N.; Lyimo, T.; Bandeira, S.O. Sea Urchin Overgrazing of Seagrasses: A Review of Current Knowledge on Causes, Consequences, and Management. Estuar. Coast. Shelf Sci. 2008, 79, 569–580. [Google Scholar] [CrossRef]
- Francour, P.; Ganteaume, A.; Poulain, M. Effects of Boat Anchoring in Posidonia oceanica Seagrass Beds in the Port-Cros National Park (North-western Mediterranean Sea). Aquat. Conserv. Mar. Freshw. Ecosyst. 1999, 9, 391–400. [Google Scholar] [CrossRef]
- Muehlstein, L.K.; Porter, D.; Short, F.T. Labyrinthula zosterae sp. nov., the Causative Agent of Wasting Disease of Eelgrass, Zostera marina. Mycologia 1991, 83, 180. [Google Scholar] [CrossRef]
- Garcias-Bonet, N.; Sherman, T.D.; Duarte, C.M.; Marbà, N. Distribution and Pathogenicity of the Protist labyrinthula Sp. in Western Mediterranean Seagrass Meadows. Estuaries Coasts 2011, 34, 1161–1168. [Google Scholar] [CrossRef]
- Short, F.T.; Wyllie-Echeverria, S. Natural and Human-Induced Disturbance of Seagrasses. Environ. Conserv. 1996, 23, 17–27. [Google Scholar] [CrossRef]
- Bianchi, C.N. Biodiversity Issues for the Forthcoming Tropical Mediterranean Sea. Hydrobiologia 2007, 580, 7–21. [Google Scholar] [CrossRef]
- Thomson, J.A.; Burkholder, D.A.; Heithaus, M.R.; Fourqurean, J.W.; Fraser, M.W.; Statton, J.; Kendrick, G.A. Extreme Temperatures, Foundation Species, and Abrupt Ecosystem Change: An Example from an Iconic Seagrass Ecosystem. Glob. Change Biol. 2015, 21, 1463–1474. [Google Scholar] [CrossRef] [PubMed]
- Borum, J.; Duarte, C.M.; Krause-Jensen, D.; Greve, T.M. European Seagrasses: An Introduction to Monitoring and Management; The EU Project Monitoring and Management of European Seagrasses: Hilleroed, Danmark, 2004; ISBN 87-89143-21-3. [Google Scholar]
- Lipej, L.; Turk, R.; Makovec, T.; Orlando-Bonaca, M.; Bettoso, N.; Malej, A.; Ciglič, H.; Bernetič, V.; Furlan, B.; Genov, T. Ogrožene Vrste in Habitatni Tipi v Slovenskem Morju: Endangered Species and Habitat Types in the Slovenian Sea; Zavod RS za Varstvo Narave: Ljubljana, Slovenia, 2006; ISBN 961-91505-4-6. [Google Scholar]
- Agostini, S.; Pergent, G.; Marchand, B. Growth and Primary Production of Cymodocea nodosa in a Coastal Lagoon. Aquat. Bot. 2003, 76, 185–193. [Google Scholar] [CrossRef]
- Buia, M.; Russo, G.; Mazzella, L. Interrelazioni Tra Cymodocea nodosa (Ucria) Aschers. e Zostera noltii Hornem. in Un Prato Misto Superficiale Dell’isola d’Ischia. Nova Thalass. 1985, 7, 406–408. [Google Scholar]
- Relini, G.; Giaccone, G. Priority Habitats According to the SPA/BIO Protocol (Barcelona Convention) Present in Italy. Identification Sheets. Biol. Mar. Mediterr. 2009, 16, 1–372. [Google Scholar]
- Orlando-Bonaca, M.; Francé, J.; Mavrič, B.; Grego, M.; Lipej, L.; Flander-Putrle, V.; Šiško, M.; Falace, A. A New Index (MediSkew) for the Assessment of the Cymodocea nodosa (Ucria) Ascherson Meadow’s Status. Mar. Environ. Res. 2015, 110, 132–141. [Google Scholar] [CrossRef]
- Duarte, C.M.; Marbà, N.; Krause-Jensen, D.; Sánchez-Camacho, M. Testing the Predictive Power of Seagrass Depth Limit Models. Estuaries Coasts 2007, 30, 652–656. [Google Scholar] [CrossRef]
- Papathanasiou, V.; Orfanidis, S.; Brown, M.T. Cymodocea nodosa Metrics as Bioindicators of Anthropogenic Stress in N. Aegean, Greek Coastal Waters. Ecol. Indic. 2016, 63, 61–70. [Google Scholar] [CrossRef]
- Traganos, D.; Reinartz, P. Mapping Mediterranean Seagrasses with Sentinel-2 Imagery. Mar. Pollut. Bull. 2018, 134, 197–209. [Google Scholar] [CrossRef] [Green Version]
- Casas, E.; Martín-García, L.; Otero-Ferrer, F.; Tuya, F.; Haroun, R.; Arbelo, M. Economic Mapping and Assessment of Cymodocea nodosa Meadows as Nursery Grounds for Commercially Important Fish Species. A Case Study in the Canary Islands. One Ecosyst. 2021, 6, e70919. [Google Scholar] [CrossRef]
- Mačić, V.; Zordan, C. Mapping of the Cymodocea nodosa (Ucria) Asch. Meadows in the Kotor Bay and Data Comparison over the Last Four Decades. Stud. Mar. 2018, 31, 5–15. [Google Scholar]
- Calleja, F.; Galván, C.; Silió-Calzada, A.; Juanes, J.A.; Ondiviela, B. Long-Term Analysis of Zostera noltei: A Retrospective Approach for Understanding Seagrasses’ Dynamics. Mar. Environ. Res. 2017, 130, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Duffy, J.P.; Pratt, L.; Anderson, K.; Land, P.E.; Shutler, J.D. Spatial Assessment of Intertidal Seagrass Meadows Using Optical Imaging Systems and a Lightweight Drone. Estuar. Coast. Shelf Sci. 2018, 200, 169–180. [Google Scholar] [CrossRef]
- Pu, R.; Bell, S.; Meyer, C.; Baggett, L.; Zhao, Y. Mapping and Assessing Seagrass along the Western Coast of Florida Using Landsat TM and EO-1 ALI/Hyperion Imagery. Estuar. Coast. Shelf Sci. 2012, 115, 234–245. [Google Scholar] [CrossRef]
- Montefalcone, M.; Rovere, A.; Parravicini, V.; Albertelli, G.; Morri, C.; Bianchi, C.N. Reprint of “Evaluating Change in Seagrass Meadows: A Time-Framed Comparison of Side Scan Sonar Maps”. Aquat. Bot. 2014, 115, 36–44. [Google Scholar] [CrossRef]
- Hedley, J.D.; Roelfsema, C.; Brando, V.; Giardino, C.; Kutser, T.; Phinn, S.; Mumby, P.J.; Barrilero, O.; Laporte, J.; Koetz, B. Coral Reef Applications of Sentinel-2: Coverage, Characteristics, Bathymetry and Benthic Mapping with Comparison to Landsat 8. Remote Sens. Environ. 2018, 216, 598–614. [Google Scholar] [CrossRef]
- Zoffoli, M.L.; Gernez, P.; Rosa, P.; Le Bris, A.; Brando, V.E.; Barillé, A.-L.; Harin, N.; Peters, S.; Poser, K.; Spaias, L.; et al. Sentinel-2 Remote Sensing of Zostera noltei-Dominated Intertidal Seagrass Meadows. Remote Sens. Environ. 2020, 251, 112020. [Google Scholar] [CrossRef]
- Dekker, T.; Greve, G.D.; Ter Laak, T.L.; Boivin, M.E.; Veuger, B.; Gortzak, G.; Dumfries, S.; Lücker, S.M.G.; Kraak, M.H.S.; Admiraal, W.; et al. Development and Application of a Sediment Toxicity Test Using the Benthic Cladoceran Chydorus sphaericus. Environ. Pollut. 2006, 140, 231–238. [Google Scholar] [CrossRef] [Green Version]
- Traganos, D.; Aggarwal, B.; Poursanidis, D.; Topouzelis, K.; Chrysoulakis, N.; Reinartz, P. Towards Global-Scale Seagrass Mapping and Monitoring Using Sentinel-2 on Google Earth Engine: The Case Study of the Aegean and Ionian Seas. Remote Sens. 2018, 10, 1227. [Google Scholar] [CrossRef] [Green Version]
- Traganos, D.; Reinartz, P. Interannual Change Detection of Mediterranean Seagrasses Using RapidEye Image Time Series. Front. Plant. Sci. 2018, 9, 96. [Google Scholar] [CrossRef] [Green Version]
- Orth, S.; Carruthers, T.; Dennison, W.; Duarte, C.; Fourqurean, J.; Heck, K.; Hughes, A.; Kendrick, G.; Kenworthy, W. A Global Crisis for Seagrass Ecosystems Bioscience. BioScience 2006, 56, 987–996. [Google Scholar] [CrossRef] [Green Version]
- Giannoulaki, M.; Belluscio, A.; Colloca, F.; Fraschetti, S.; Scardi, M.; Smith, C.; Spedicato, M. Mediterranean Sensitive Habitats: DG MARE Specific Contract SI2; Hellenic Centre for Marine Research: Heraklion, Greece, 2013. [Google Scholar]
- Knudby, A.; Nordlund, L. Remote Sensing of Seagrasses in a Patchy Multi-Species Environment. Int. J. Remote Sens. 2011, 32, 2227–2244. [Google Scholar] [CrossRef]
- Hossain, M.S.; Bujang, J.S.; Zakaria, M.H.; Hashim, M. The Application of Remote Sensing to Seagrass Ecosystems: An Overview and Future Research Prospects. Int. J. Remote Sens. 2015, 36, 61–114. [Google Scholar] [CrossRef]
- Topouzelis, K.; Makri, D.; Stoupas, N.; Papakonstantinou, A.; Katsanevakis, S. Seagrass Mapping in Greek Territorial Waters Using Landsat-8 Satellite Images. Int. J. Appl. Earth Obs. Geoinf. 2018, 67, 98–113. [Google Scholar] [CrossRef]
- McKenzie, L.J.; Nordlund, L.M.; Jones, B.L.; Cullen-Unsworth, L.C.; Roelfsema, C.; Unsworth, R.K. The Global Distribution of Seagrass Meadows. Environ. Res. Lett. 2020, 15, 074041. [Google Scholar] [CrossRef]
- Borfecchia, F.; De Cecco, L.; Martini, S.; Ceriola, G.; Bollanos, S.; Vlachopoulos, G.; Valiante, L.M.; Belmonte, A.; Micheli, C. Posidonia Oceanica Genetic and Biometry Mapping through High-Resolution Satellite Spectral Vegetation Indices and Sea-Truth Calibration. Int. J. Remote Sens. 2013, 34, 4680–4701. [Google Scholar] [CrossRef]
- Bonacorsi, M.; Pergent-Martini, C.; Breand, N.; Pergent, G. Is Posidonia oceanica Regression a General Feature in the Mediterranean Sea? Mediterr. Mar. Sci. 2013, 14, 193. [Google Scholar] [CrossRef]
- Matta, E.; Aiello, M.; Bresciani, M.; Gianinetto, M.; Musanti, M.; Giardino, C. Mapping Posidonia Meadow from High Spatial Resolution Images in the Gulf of Oristano (Italy). In Proceedings of the 2014 IEEE Geoscience and Remote Sensing Symposium, Quebec City, QC, Canada, 13–18 July 2014; pp. 5152–5155. [Google Scholar]
- Pergent-Martini, C.; Pergent, G.; Monnier, B.; Boudouresque, C.-F.; Mori, C.; Valette-Sansevin, A. Contribution of Posidonia oceanica Meadows in the Context of Climate Change Mitigation in the Mediterranean Sea. Mar. Environ. Res. 2021, 165, 105236. [Google Scholar] [CrossRef]
- Garrido, M.; Lafabrie, C.; Torre, F.; Fernandez, C.; Pasqualini, V. Resilience and Stability of Cymodocea nodosa Seagrass Meadows over the Last Four Decades in a Mediterranean Lagoon. Estuar. Coast. Shelf Sci. 2013, 130, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Chefaoui, R.M.; Assis, J.; Duarte, C.M.; Serrão, E.A. Large-Scale Prediction of Seagrass Distribution Integrating Landscape Metrics and Environmental Factors: The Case of Cymodocea nodosa (Mediterranean–Atlantic). Estuaries Coasts 2016, 39, 123–137. [Google Scholar] [CrossRef]
- Kaufman, K.A.; Bell, S.S. The Use of Imagery and GIS Techniques to Evaluate and Compare Seagrass Dynamics across Multiple Spatial and Temporal Scales. Estuaries Coasts 2020, 43, 1–17. [Google Scholar] [CrossRef]
- Kuhwald, K.; Schneider von Deimling, J.; Schubert, P.; Oppelt, N. How Can Sentinel-2 Contribute to Seagrass Mapping in Shallow, Turbid Baltic Sea Waters? Remote Sens. Ecol. Conserv. 2021, 7, rse2.246. [Google Scholar] [CrossRef]
- Carpenter, S.; Byfield, V.; Felgate, S.L.; Price, D.M.; Andrade, V.; Cobb, E.; Strong, J.; Lichtschlag, A.; Brittain, H.; Barry, C.; et al. Using Unoccupied Aerial Vehicles (UAVs) to Map Seagrass Cover from Sentinel-2 Imagery. Remote Sens. 2022, 14, 477. [Google Scholar] [CrossRef]
- Fauzan, M.A.; Wicaksono, P. Hartono Characterizing Derawan Seagrass Cover Change with Time-Series Sentinel-2 Images. Reg. Stud. Mar. Sci. 2021, 48, 102048. [Google Scholar] [CrossRef]
- Raziskava z Oceno Stanja Morskih Travnikov v Krajinskem Parku Strunjan: Zaključno Poročilo o Izvedenem Delu; Morska Bološka Postaja, Nacionalni Inštitut Za Biologijo: Piran, Slovenia, 2018.
- Saunders, M.I.; Atkinson, S.; Klein, C.J.; Weber, T.; Possingham, H.P. Increased Sediment Loads Cause Non-Linear Decreases in Seagrass Suitable Habitat Extent. PLoS ONE 2017, 12, e0187284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staggl, M.; Egger, C.; Schwienbacher, B.; Heß, M.; Mavrič, B.; Lipej, L.; Orlando-Bonaca, M.; Ivajnšič, D.; Dujmović, S.; Smodlaka Tanković, M.; et al. Habitat loss in the Northern Adriatic: Monitoring of seagrass beds on the Eastern coast of the northern Adriatic using GPS tracking and satellite image analysis reveals a dramatic die-off between 2013 and 2020. Sci. Rep. Manuscript submitted for publication.
- Diaz-Almela, E.; Marbà, N.; Duarte, C.M. Consequences of Mediterranean Warming Events in Seagrass (Posidonia oceanica) Flowering Records. Glob. Change Biol. 2007, 13, 224–235. [Google Scholar] [CrossRef]
- Jueterbock, A.; Franssen, S.U.; Bergmann, N.; Gu, J.; Coyer, J.A.; Reusch, T.B.H.; Bornberg-Bauer, E.; Olsen, J.L. Phylogeographic Differentiation versus Transcriptomic Adaptation to Warm Temperatures in Zostera marina, a Globally Important Seagrass. Mol. Ecol. 2016, 25, 5396–5411. [Google Scholar] [CrossRef] [Green Version]
- Repolho, T.; Duarte, B.; Dionísio, G.; Paula, J.R.; Lopes, A.R.; Rosa, I.C.; Grilo, T.F.; Caçador, I.; Calado, R.; Rosa, R. Seagrass Ecophysiological Performance under Ocean Warming and Acidification. Sci. Rep. 2017, 7, 41443. [Google Scholar] [CrossRef]
- Duarte, B.; Martins, I.; Rosa, R.; Matos, A.R.; Roleda, M.Y.; Reusch, T.B.H.; Engelen, A.H.; Serrão, E.A.; Pearson, G.A.; Marques, J.C.; et al. Climate Change Impacts on Seagrass Meadows and Macroalgal Forests: An Integrative Perspective on Acclimation and Adaptation Potential. Front. Mar. Sci. 2018, 5, 190. [Google Scholar] [CrossRef] [Green Version]
- Verdura, J.; de Caralt, S.; Santamaria, J. Seventh European Phycological Congress. Eur. J. Phycol. 2019, 54, 63. [Google Scholar] [CrossRef]
- Assis, J.; Fragkopoulou, E.; Frade, D.; Neiva, J.; Oliveira, A.; Abecasis, D.; Faugeron, S.; Serrão, E.A. A Fine-Tuned Global Distribution Dataset of Marine Forests. Sci. Data 2020, 7, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, M.; Romero, J. Photosynthetic Response to Light and Temperature of the Seagrass Cymodocea nodosa and the Prediction of Its Seasonality. Aquat. Bot. 1992, 43, 51–62. [Google Scholar] [CrossRef]
- Hendriks, I.E.; Olsen, Y.S.; Ramajo, L.; Basso, L.; Steckbauer, A.; Moore, T.S.; Howard, J.; Duarte, C.M. Photosynthetic Activity Buffers Ocean Acidification in Seagrass Meadows. Biogeosciences 2014, 11, 333–346. [Google Scholar] [CrossRef] [Green Version]
- Olsen, Y.S.; Potouroglou, M.; Garcias-Bonet, N.; Duarte, C.M. Warming Reduces Pathogen Pressure on a Climate-Vulnerable Seagrass Species. Estuaries Coasts 2015, 38, 659–667. [Google Scholar] [CrossRef]
- Ogorelec, B.; Mišič, M.; Faganeli, J. Marine Geology of the Gulf of Trieste (Northern Adriatic): Sedimentological Aspects. Mar. Geol. 1991, 99, 79–92. [Google Scholar] [CrossRef]
- Malačič, V.; Petelin, B. Climatic Circulation in the Gulf of Trieste (Northern Adriatic). J. Geophys. Res. Oceans 2009, 114, 1–15. [Google Scholar] [CrossRef]
- Turk, R. An Assessment of the Vulnerability of the Slovene Coastal Belt and Its Categorisation in View of in (Admissible) Human Pressure, Various Activities and Land-Use. Annales 1999, 1, 37–50. [Google Scholar]
- Solidoro, C.; Bastianini, M.; Bandelj, V.; Codermatz, R.; Cossarini, G.; Melaku Canu, D.; Ravagnan, E.; Salon, S.; Trevisani, S. Current State, Scales of Variability, and Trends of Biogeochemical Properties in the Northern Adriatic Sea. J. Geophys. Res. 2009, 114, C07S91. [Google Scholar] [CrossRef]
- Mozetič, P.; Francé, J.; Kogovšek, T.; Talaber, I.; Malej, A. Plankton Trends and Community Changes in a Coastal Sea (Northern Adriatic): Bottom-up vs. Top-down Control in Relation to Environmental Drivers. Estuar. Coast. Shelf Sci. 2012, 115, 138–148. [Google Scholar] [CrossRef]
- Mozetič, P.; Solidoro, C.; Cossarini, G.; Socal, G.; Precali, R.; Francé, J.; Bianchi, F.; De Vittor, C.; Smodlaka, N.; Fonda Umani, S. Recent Trends Towards Oligotrophication of the Northern Adriatic: Evidence from Chlorophyll a Time Series. Estuaries Coasts 2010, 33, 362–375. [Google Scholar] [CrossRef] [Green Version]
- Žerjal, A.; Kolega, N.; Poglajen, S.; Rant, I.; Jeklar, M.; Lovrič, E.; Vranac, D.; Mozetič, D.; Slavec, P.; Zrimec, M.B. Zajem Naravnih Geomorfoloških Značilnosti Morskega dna, Analiza Antropogenih Fizičnih Poškodb Morskega dna in Klasifikacija Tipov Morskega dna z Določitvijo Obsežnejšega Morskega Rastja na Morskem dnu; Inštitut za Vode Republike Slovenije: Ljubljana, Slovenia, 2014. [Google Scholar]
- Ministry of the Environment and Spatial Planning, The Surveying and Mapping Authority of the Republic of Slovenia E-Surveying Data. Available online: https://egp.gu.gov.si/egp/?lang=en (accessed on 2 August 2020).
- Ballesteros, E.; Torras, X.; Pinedo, S.; García, M.; Mangialajo, L.; de Torres, M. A New Methodology Based on Littoral Community Cartography Dominated by Macroalgae for the Implementation of the European Water Framework Directive. Mar. Pollut. Bull. 2007, 55, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Nikolić, V.; Žuljević, A.; Mangialajo, L.; Antolić, B.; Kušpilić, G.; Ballesteros, E. Cartography of Littoral Rocky-Shore Communities (CARLIT) as a Tool for Ecological Quality Assessment of Coastal Waters in the Eastern Adriatic Sea. Ecol. Indic. 2013, 34, 87–93. [Google Scholar] [CrossRef]
- Orlando-Bonaca, M.; Lipej, L.; Orfanidis, S. Benthic Macrophytes as a Tool for Delineating, Monitoring and Assessing Ecological Status: The Case of Slovenian Coastal Waters. Mar. Pollut. Bull. 2008, 56, 666–676. [Google Scholar] [CrossRef]
- Orlando-Bonaca, M.; Pitacco, V.; Lipej, L. Loss of Canopy-Forming Algal Richness and Coverage in the Northern Adriatic Sea. Ecol. Indic. 2021, 125, 107501. [Google Scholar] [CrossRef]
- Stumpf, R.P.; Holderied, K.; Sinclair, M. Determination of Water Depth with High-Resolution Satellite Imagery over Variable Bottom Types. Limnol. Oceanogr. 2003, 48, 547–556. [Google Scholar] [CrossRef]
- Maritorena, S.; Morel, A.; Gentili, B. Diffuse Reflectance of Oceanic Shallow Waters: Influence of Water Depth and Bottom Albedo. Limnol. Oceanogr. 1994, 39, 1689–1703. [Google Scholar] [CrossRef]
- Bierwirth, P.; Lee, T.; Burne, R. Shallow Sea-Floor Reflectance and Water Depth Derived by Unmixing Multispectral Imagery. Photogramm. Eng. Remote Sens. 1993, 59, 331–338. [Google Scholar]
- Congalton, R.G. A Review of Assessing the Accuracy of Classifications of Remotely Sensed Data. Remote Sens. Environ. 1991, 37, 35–46. [Google Scholar] [CrossRef]
- Green, E.; Mumby, P.; Edwards, A.; Clark, C. Remote Sensing: Handbook for Tropical Coastal Management; United Nations Educational, Scientific and Cultural Organization (UNESCO): Paris, France, 2000. [Google Scholar]
- ESRI. ArcGIS Desktop: Release 10.8.; Environmental Systems Research Institute: Redlands, CA, USA, 2020. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SVM | Seagrass | Other | Total | User Accuracy |
| Seagrass | 324 | 21 | 345 | 0.94 |
| Other | 60 | 97 | 157 | 0.62 |
| Total | 384 | 118 | 502 | |
| Producer Accuracy | 0.8420 | 0.82 | ||
| Overall Accuracy | 0.84 | |||
| Kappa | 0.60 | |||
| MLC | Seagrass | Other | Total | User Accuracy |
| Seagrass | 319 | 20 | 339 | 0.94 |
| Other | 63 | 97 | 160 | 0.61 |
| Total | 382 | 117 | 499 | |
| Producer Accuracy | 0.83 | 0.83 | ||
| Overall Accuracy | 0.83 | |||
| Kappa | 0.59 | |||
| RTC | Seagrass | Other | Total | User Accuracy |
| Seagrass | 303 | 15 | 318 | 0.95 |
| Other | 79 | 102 | 181 | 0.56 |
| Total | 382 | 117 | 499 | |
| Producer Accuracy | 0.79 | 0.87 | ||
| Overall Accuracy | 0.81 | |||
| Kappa | 0.56 | |||
| Kappa | 0.56 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivajnšič, D.; Orlando-Bonaca, M.; Donša, D.; Grujić, V.J.; Trkov, D.; Mavrič, B.; Lipej, L. Evaluating Seagrass Meadow Dynamics by Integrating Field-Based and Remote Sensing Techniques. Plants 2022, 11, 1196. https://doi.org/10.3390/plants11091196
Ivajnšič D, Orlando-Bonaca M, Donša D, Grujić VJ, Trkov D, Mavrič B, Lipej L. Evaluating Seagrass Meadow Dynamics by Integrating Field-Based and Remote Sensing Techniques. Plants. 2022; 11(9):1196. https://doi.org/10.3390/plants11091196
Chicago/Turabian StyleIvajnšič, Danijel, Martina Orlando-Bonaca, Daša Donša, Veno Jaša Grujić, Domen Trkov, Borut Mavrič, and Lovrenc Lipej. 2022. "Evaluating Seagrass Meadow Dynamics by Integrating Field-Based and Remote Sensing Techniques" Plants 11, no. 9: 1196. https://doi.org/10.3390/plants11091196
APA StyleIvajnšič, D., Orlando-Bonaca, M., Donša, D., Grujić, V. J., Trkov, D., Mavrič, B., & Lipej, L. (2022). Evaluating Seagrass Meadow Dynamics by Integrating Field-Based and Remote Sensing Techniques. Plants, 11(9), 1196. https://doi.org/10.3390/plants11091196