

Characterizing Root Morphological Responses to Exogenous Tryptophan in Soybean (Glycine max) Seedlings Using a Scanner-Based Rhizotron System


Abstract
:1. Introduction
2. Results
2.1. Canopy Projected Area
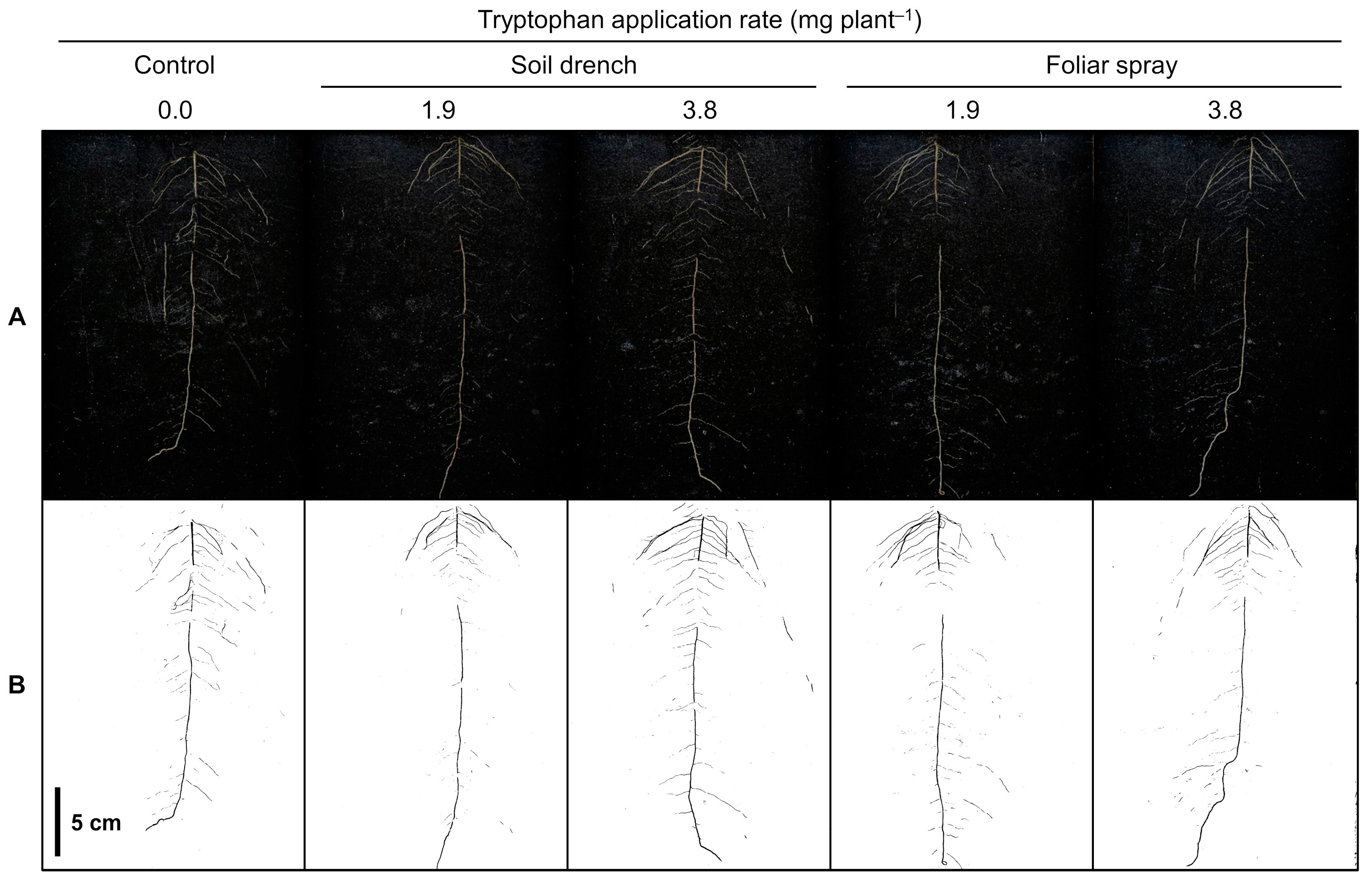
2.2. Root Projected Area (Rhizotron Image Analysis)
2.3. Root Diameter and Lateral Root Number (WinRHIZO Image Analysis)
2.4. Root Length (WinRHIZO Image Analysis)
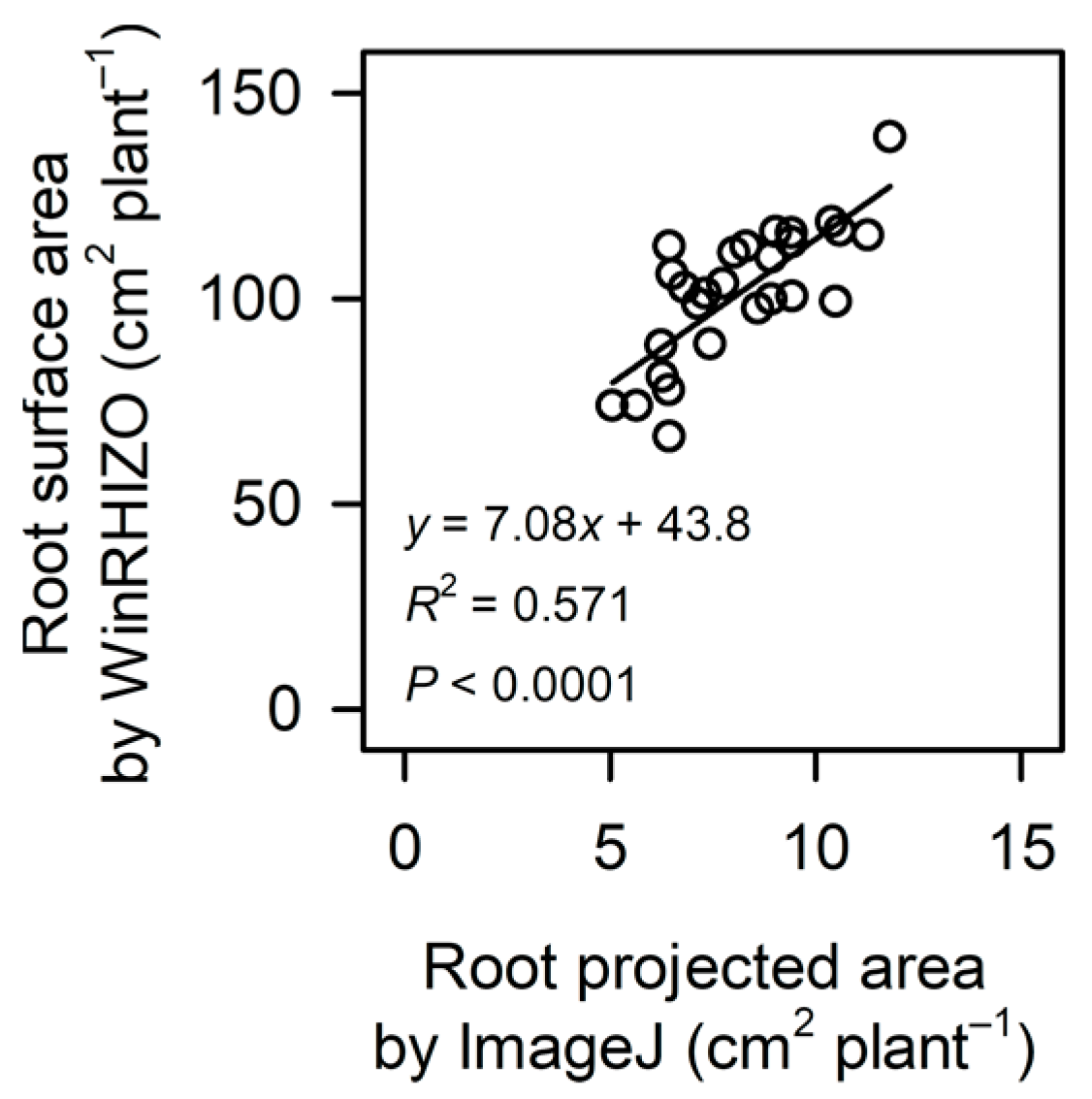
2.5. Root Surface Area (WinRHIZO Image Analysis)
2.6. Shoot and Root Growth (Plant Sampling)
3. Discussion
3.1. Exogenous Tryptophan Can Induce Auxin-Like Activities in Root Development
3.2. Tryptophan Is More Effective in Promoting Fine Root Development When Applied by Soil Drenching Than by Foliar Spray
3.3. The Optimum Application Rate of Tryptophan Depends on the Application Method
3.4. Practical Implementation
4. Materials and Methods
4.1. Tryptophan Treatments
4.2. Rhizotron Experiment
4.3. Canopy Image Analysis
4.4. Rhizotron Root Image Analysis
4.5. Plant Sampling and Growth Measurements
4.6. Image Analysis of Washed Roots
4.7. Experiment Design and Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant Physiology and Development, 6th ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2015. [Google Scholar]
- Blakely, L.; Durham, M.; Evans, T.; Blakely, R.M. Experimental studies on lateral root formation in radish seedling roots. I. General methods, developmental stages, and spontaneous formation of laterals. Bot. Gaz. 1982, 143, 341–352. [Google Scholar] [CrossRef]
- Laskowski, M.J.; Williams, M.E.; Nusbaum, H.C.; Sussex, I.M. Formation of lateral root meristems is a two-stage process. Development 1995, 121, 3303–3310. [Google Scholar] [CrossRef]
- Casimiro, I.; Marchant, A.; Bhalerao, R.P.; Beeckman, T.; Dhooge, S.; Swarup, R.; Graham, N.; Inzé, D.; Sandberg, G.; Casero, P.J. Auxin transport promotes Arabidopsis lateral root initiation. Plant Cell 2001, 13, 843–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashiguchi, K.; Tanaka, K.; Sakai, T.; Sugawara, S.; Kawaide, H.; Natsume, M.; Hanada, A.; Yaeno, T.; Shirasu, K.; Yao, H.; et al. The main auxin biosynthesis pathway in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 18512–18517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Won, C.; Shen, X.; Mashiguchi, K.; Zheng, Z.; Dai, X.; Cheng, Y.; Kasahara, H.; Kamiya, Y.; Chory, J.; Zhao, Y. Conversion of tryptophan to indole-3-acetic acid by TRYPTOPHAN AMINOTRANSFERASES OF ARABIDOPSIS and YUCCAs in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 18518–18523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y. Auxin biosynthesis: A simple two-step pathway converts tryptophan to indole-3-acetic acid in plants. Mol. Plant 2012, 5, 334–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frankenberger, W.T., Jr.; Brunner, W. Method of detection of auxin-indole-3-acetic acid in soils by high performance liquid chromatography. Soil Sci. Soc. Am. J. 1983, 47, 237–241. [Google Scholar] [CrossRef]
- Patten, C.L.; Glick, B.R. Bacterial biosynthesis of indole-3-acetic acid. Can. J. Microbiol. 1996, 42, 207–220. [Google Scholar] [CrossRef]
- Davies, P.J. Plant Hormones, 3rd ed.; Springer Dordrecht: Berlin, Germany, 2010. [Google Scholar]
- Abou Dahab, T.A.M.; Abd El-Aziz, N.G. Physiological effect of diphenylamin and tryptophan on the growth and chemical constituents of Philodendron erubescens plants. World J. Agric. Sci. 2006, 2, 75–81. [Google Scholar]
- Sudadi, S.; Suryono, S. Exogenous application of tryptophan and indole acetic acid (IAA) to induce root nodule formation and increase soybean yield in acid, neutral and alkaline soil. AGRIVITA J. Agric. Sci. 2015, 37, 37–44. [Google Scholar] [CrossRef]
- Mustafa, A.; Hussain, A.; Naveed, M.; Ditta, A.; Nazli, Z.-e.-H.; Sattar, A. Response of okra (Abelmoschus esculentus L.) to soil and foliar applied L-tryptophan. Soil Environ. 2016, 35, 76–84. [Google Scholar]
- Schuurman, J.J.; Goedewaagen, M.A.J. Methods for the Examination of Root Systems and Roots; Centre for Agricultural Publications and Documentation: Wageningen, The Netherlands, 1965. [Google Scholar]
- Seethepalli, A.; Guo, H.; Liu, X.; Griffiths, M.; Almtarfi, H.; Li, Z.; Liu, S.; Zare, A.; Fritschi, F.B.; Blancaflor, E.B. RhizoVision Crown: An integrated hardware and software platform for root crown phenotyping. Plant Phenomics 2020, 2020, 3074916. [Google Scholar] [CrossRef] [PubMed]
- Adu, M.O.; Yawson, D.O.; Bennett, M.J.; Broadley, M.R.; Dupuy, L.X.; White, P.J. A scanner-based rhizobox system enabling the quantification of root system development and response of Brassica rapa seedlings to external P availability. Plant Root 2017, 11, 16–32. [Google Scholar] [CrossRef] [Green Version]
- Davies, M.J.; Hipps, N.A.; Kingswell, G. The effects of indole-3-butyric acid root dips on the root development and shoot growth of transplanted Fagus sylvatica L. and Quercus robur L. seedlings. J. Hortic. Sci. Biotechnol. 2002, 77, 209–216. [Google Scholar] [CrossRef]
- Nagel, K.A.; Putz, A.; Gilmer, F.; Heinz, K.; Fischbach, A.; Pfeifer, J. GROWSCREEN-Rhizo is a novel phenotyping robot enabling simultaneous measurements of root and shoot growth for plants grown in soil-filled rhizotrons. Funct. Plant Biol. 2012, 39, 891–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agehara, S.; Sanada, A. Quick and inexpensive root and shoot evaluation methods using a scanner-based rhizotron system and ImageJ in soybean (Glycine max). Root Res. 2020, 29, 5–19. [Google Scholar] [CrossRef]
- Qin, H.; Huang, R. Auxin controlled by ethylene steers root development. Int. J. Mol. Sci. 2018, 19, 3656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, H.; Strader, L.C. Interplay of auxin and cytokinin in lateral root development. Int. J. Mol. Sci. 2019, 20, 486. [Google Scholar] [CrossRef] [Green Version]
- Poulet, A.; Kriechbaumer, V. Bioinformatics analysis of phylogeny and transcription of TAA/YUC auxin biosynthetic genes. Int. J. Mol. Sci. 2017, 18, 1791. [Google Scholar] [CrossRef] [Green Version]
- Gondek, K.; Mierzwa-Hersztek, M. Effect of soil-applied L-tryptophan on the amount of biomass and nitrogen and sulfur utilization by maize. Agronomy 2021, 11, 2582. [Google Scholar] [CrossRef]
- Mustafa, A.; Imran, M.; Ashraf, M.; Mahmood, K. Perspectives of using L-tryptophan for improving productivity of agricultural crops: A review. Pedosphere 2018, 28, 16–34. [Google Scholar] [CrossRef]
- Rao, S.R.; Qayyum, A.; Razzaq, A.; Ahmad, M.; Mahmood, I.; Sher, A. Role of foliar application of salicylic acid and l-tryptophan in drought tolerance of maize. J. Anim. Plant Sci. 2012, 22, 768–772. [Google Scholar]
- Yang, G.; Wei, Q.; Huang, H.; Xia, J. Amino acid transporters in plant cells: A brief review. Plants 2020, 9, 967. [Google Scholar] [CrossRef] [PubMed]
- Benjamins, R.; Scheres, B. Auxin: The looping star in plant development. Annu. Rev. Plant Biol. 2008, 59, 443–465. [Google Scholar] [CrossRef] [PubMed]
- Overvoorde, P.; Fukaki, H.; Beeckman, T. Auxin control of root development. Cold Spring Harb. Perspect. Biol. 2010, 2, a001537. [Google Scholar] [CrossRef] [Green Version]
- Janas, T.; Janas, T.; Yarus, M. A membrane transporter for tryptophan composed of RNA. RNA 2004, 10, 1541–1549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radwanski, E.R.; Last, R.L. Tryptophan biosynthesis and metabolism: Biochemical and molecular genetics. Plant Cell 1995, 7, 921. [Google Scholar] [CrossRef] [Green Version]
- Stepanova, A.N.; Alonso, J.M. Auxin catabolism unplugged: Role of IAA oxidation in auxin homeostasis. Proc. Natl. Acad. Sci. USA 2016, 113, 10742–10744. [Google Scholar] [CrossRef] [Green Version]
- Taiz, L.; Zeiger, E. Plant Physiology, 5th ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2010. [Google Scholar]
- Xu, W.; Cui, K.; Xu, A.; Nie, L.; Huang, J.; Peng, S. Drought stress condition increases root to shoot ratio via alteration of carbohydrate partitioning and enzymatic activity in rice seedlings. Acta Physiol. Plant. 2015, 37, 9. [Google Scholar] [CrossRef]
- Bacher, H.; Sharaby, Y.; Walia, H.; Peleg, Z. Modifying root-to-shoot ratio improves root water influxes in wheat under drought stress. J. Expt. Bot. 2021, 73, 1643–1654. [Google Scholar] [CrossRef]
- Mardanov, A.; Samedovam, A.; Shirvany, T. Root-shoot relationships in plant adaptation to nitrogen deficiency. In Root Demographics and Their Efficiencies in Sustainable Agriculture, Grasslands and Forest Ecosystems; Springer: Berlin/Heidelberg, Germany, 1998; pp. 147–154. [Google Scholar]
- Agehara, S. Simple imaging techniques for plant growth assessment. EDIS 2020, 2020, HS1353. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | TP Application Rate | Canopy Projected Area (cm2 plant−1) | ||
|---|---|---|---|---|
| Method 2 | (mg plant−1) | 11 DAS | 14 DAS | 18 DAS |
| Control | 0.0 | 11.7 | 21.7 | 25.3 b |
| Soil drench | 1.9 | 11.7 | 21.9 | 28.5 ab |
| 3.8 | 15.0 | 26.7 | 32.2 a | |
| Foliar spray | 1.9 | 11.3 | 22.7 | 29.3 ab |
| 3.8 | 11.2 | 20.8 | 25.5 b | |
| Pooled data | ||||
| All TP treatments | 12.3 | 23.0 | 28.9 | |
| All TP soil drench treatments | 13.3 | 24.3 | 30.3 | |
| All TP foliar spray treatments | 11.2 | 21.8 | 27.4 | |
| p value | ||||
| Treatment effect | 0.1929 | 0.1508 | 0.0440 | |
| Untreated vs. All TP | 0.6606 | 0.4930 | 0.0672 | |
| Untreated vs. All TP soil drench | 0.2796 | 0.2226 | 0.0218 | |
| Untreated vs. All TP foliar spray | 0.7691 | 0.9873 | 0.3056 | |
| All TP soil drench vs. All TP foliar spray | 0.0998 | 0.1436 | 0.0985 | |
| Treatment | TP Application Rate | Root Projected Area (cm2 plant−1) | ||||
|---|---|---|---|---|---|---|
| Method 2 | (mg plant−1) | 5 DAS | 8 DAS | 11 DAS | 14 DAS | 18 DAS |
| Control | 0.0 | 1.21 | 3.47 | 5.15 | 6.27 | 7.42 B |
| Soil drench | 1.9 | 0.99 | 2.87 | 4.25 | 5.61 | 6.93 B |
| 3.8 | 1.14 | 3.78 | 5.84 | 7.35 | 9.60 A | |
| Foliar spray | 1.9 | 1.12 | 3.67 | 5.34 | 6.85 | 8.36 AB |
| 3.8 | 1.10 | 3.56 | 5.60 | 6.75 | 8.58 AB | |
| Pooled data | ||||||
| All TP treatments | 1.09 | 3.47 | 5.26 | 6.64 | 8.37 | |
| All TP soil drench treatments | 1.07 | 3.32 | 5.05 | 6.48 | 8.27 | |
| All TP foliar spray treatments | 1.11 | 3.62 | 5.47 | 6.80 | 8.47 | |
| p value | ||||||
| Treatment effect | -- | 0.3371 | 0.3601 | 0.3863 | 0.0670 | |
| Untreated vs. All TP | -- | 0.3725 | 0.3754 | 0.2474 | 0.0570 | |
| Untreated vs. All TP soil drench | -- | 0.6593 | 0.5984 | 0.3719 | 0.0936 | |
| Untreated vs. All TP foliar spray | -- | 0.2383 | 0.2778 | 0.2256 | 0.0717 | |
| All TP soil drench vs. All TP foliar spray | -- | 0.3871 | 0.5152 | 0.7176 | 0.9108 | |
| Treatment | TP Application Rate | Root Diameter 3 | Root Number per Plant | |
|---|---|---|---|---|
| Method 2 | (mg plant−1) | (µm) | Secondary Roots | Tertiary Roots |
| Control | 0.0 | 425 | 120 C | 1590 |
| Soil drench | 1.9 | 400 | 135 AB | 1786 |
| 3.8 | 421 | 128 BC | 1844 | |
| Foliar spray | 1.9 | 412 | 142 A | 1591 |
| 3.8 | 410 | 137 AB | 1475 | |
| Pooled data | ||||
| All TP treatments | 411 | 136 | 1674 | |
| All TP soil drench treatments | 410 | 132 | 1815 | |
| All TP foliar spray treatments | 411 | 139 | 1533 | |
| p value | ||||
| Treatment effect | 0.3571 | 0.0594 | 0.9583 | |
| Control vs. All TP | 0.1591 | 0.0144 | 0.9386 | |
| Control vs. All TP soil drench | 0.2051 | 0.0841 | 0.7546 | |
| Control vs. All TP foliar spray | 0.1965 | 0.0066 | 0.7524 | |
| All TP soil drench vs. All TP foliar spray | 0.9948 | 0.1822 | 0.5371 | |
| Root Length (cm plant−1) | ||||||
|---|---|---|---|---|---|---|
| Treatment | TP Application Rate | Primary | Lateral Roots 3 | Primary + | ||
| Method 2 | (mg plant−1) | Root | ≥0.2 mm | <0.2 mm | Total | Lateral |
| Control | 0.0 | 28.8 BC | 497 | 205 | 702 | 731 |
| Soil drench | 1.9 | 30.2 AB | 547 | 251 | 798 | 828 |
| 3.8 | 27.5 C | 580 | 266 | 847 | 874 | |
| Foliar spray | 1.9 | 31.7 A | 523 | 222 | 745 | 777 |
| 3.8 | 31.2 AB | 505 | 202 | 707 | 738 | |
| Pooled data | ||||||
| All TP treatments | 30.2 | 527 | 231 | 758 | 804 | |
| All TP soil drench treatments | 28.9 | 554 | 257 | 812 | 851 | |
| All TP foliar spray treatments | 31.4 | 499 | 205 | 704 | 757 | |
| p value | ||||||
| Treatment effect (ANOVA) | 0.0518 | 0.5605 | 0.1934 | 0.3423 | 0.3554 | |
| Untreated vs. All TP | 0.0221 | 0.3177 | 0.2148 | 0.2422 | 0.2337 | |
| Untreated vs. All TP soil drench | 0.9276 | 0.1589 | 0.0522 | 0.0860 | 0.0858 | |
| Untreated vs. All TP foliar spray | 0.0384 | 0.7061 | 0.8348 | 0.7192 | 0.6895 | |
| All TP soil drench vs. All TP foliar spray | 0.0228 | 0.2150 | 0.0428 | 0.1037 | 0.1128 | |
| Root Surface Area (cm2 plant−1) | ||||||
|---|---|---|---|---|---|---|
| Treatment | TP Application Rate | Primary | Lateral Roots 3 | Primary + | ||
| Method 2 | (mg plant−1) | Roots | ≥0.2 mm | <0.2 mm | Total | Lateral |
| Control | 0.0 | 14.4 | 67.9 | 9.1 | 77.0 | 91.4 |
| Soil drench | 1.9 | 13.6 | 73.3 | 10.7 | 84.0 | 97.6 |
| 3.8 | 13.4 | 80.5 | 11.6 | 92.1 | 105.5 | |
| Foliar spray | 1.9 | 14.3 | 70.5 | 9.6 | 80.2 | 94.5 |
| 3.8 | 14.6 | 66.6 | 8.7 | 75.4 | 90.0 | |
| Pooled data | ||||||
| All TP treatments | 14.0 | 72.8 | 10.2 | 82.9 | 96.9 | |
| All TP soil drench treatments | 13.5 | 76.9 | 11.1 | 88.0 | 101.5 | |
| All TP foliar spray treatments | 14.4 | 68.6 | 9.2 | 77.8 | 92.2 | |
| p value | ||||||
| Treatment effect | 0.4372 | 0.3626 | 0.2279 | 0.3006 | 0.4072 | |
| Untreated vs. All TP | 0.4522 | 0.3814 | 0.2857 | 0.3438 | 0.3941 | |
| Untreated vs. All TP soil drench | 0.1607 | 0.1497 | 0.0770 | 0.1198 | 0.1643 | |
| Untreated vs. All TP foliar spray | 0.9371 | 0.9120 | 0.9370 | 0.9062 | 0.9034 | |
| All TP soil drench vs. All TP foliar spray | 0.0848 | 0.1172 | 0.0509 | 0.0905 | 0.1344 | |
| Treatment | TP Application Rate | Stem Diameter | Leaf Area | Dry wt (mg plant−1) | Root:Shoot | |
|---|---|---|---|---|---|---|
| Method 2 | (mg plant−1) | (mm) | (cm2 plant−1) | Shoot | Roots | Ratio (wt/wt) |
| Control | 0.0 | 2.56 ab | 36.9 ABC | 194 | 60.5 c | 0.315 b |
| Soil drench | 1.9 | 2.40 c | 38.1 AB | 188 | 72.3 bc | 0.389 a |
| 3.8 | 2.65 a | 41.6 A | 213 | 86.3 a | 0.406 a | |
| Foliar spray | 1.9 | 2.60 ab | 36.3 BC | 195 | 75.7 ab | 0.388 a |
| 3.8 | 2.45 bc | 32.0 C | 183 | 69.8 bc | 0.379 a | |
| Pooled data | ||||||
| All TP treatments | 2.52 | 37.0 | 195 | 76.0 | 0.390 | |
| All TP soil drench treatments | 2.52 | 39.8 | 201 | 79.3 | 0.397 | |
| All TP foliar spray treatments | 2.52 | 34.2 | 189 | 72.8 | 0.384 | |
| p value | ||||||
| Treatment effect | 0.0132 | 0.0597 | 0.2796 | 0.0046 | 0.0085 | |
| Untreated vs. All TP | 0.5731 | 0.9659 | 0.9850 | 0.0030 | 0.0005 | |
| Untreated vs. All TP soil drench | 0.6162 | 0.2734 | 0.6071 | 0.0013 | 0.0005 | |
| Untreated vs. All TP foliar spray | 0.5974 | 0.3078 | 0.6311 | 0.0261 | 0.0027 | |
| All TP soil drench vs. All TP foliar spray | 0.9734 | 0.0139 | 0.2290 | 0.1322 | 0.4287 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanada, A.; Agehara, S. Characterizing Root Morphological Responses to Exogenous Tryptophan in Soybean (Glycine max) Seedlings Using a Scanner-Based Rhizotron System. Plants 2023, 12, 186. https://doi.org/10.3390/plants12010186
Sanada A, Agehara S. Characterizing Root Morphological Responses to Exogenous Tryptophan in Soybean (Glycine max) Seedlings Using a Scanner-Based Rhizotron System. Plants. 2023; 12(1):186. https://doi.org/10.3390/plants12010186
Chicago/Turabian StyleSanada, Atsushi, and Shinsuke Agehara. 2023. "Characterizing Root Morphological Responses to Exogenous Tryptophan in Soybean (Glycine max) Seedlings Using a Scanner-Based Rhizotron System" Plants 12, no. 1: 186. https://doi.org/10.3390/plants12010186
APA StyleSanada, A., & Agehara, S. (2023). Characterizing Root Morphological Responses to Exogenous Tryptophan in Soybean (Glycine max) Seedlings Using a Scanner-Based Rhizotron System. Plants, 12(1), 186. https://doi.org/10.3390/plants12010186