
The Electrome of a Parasitic Plant in a Putative State of Attention Increases the Energy of Low Band Frequency Waves: A Comparative Study with Neural Systems

 , ,
, , {kind=link}
{kind=link}
Abstract
:1. Introduction
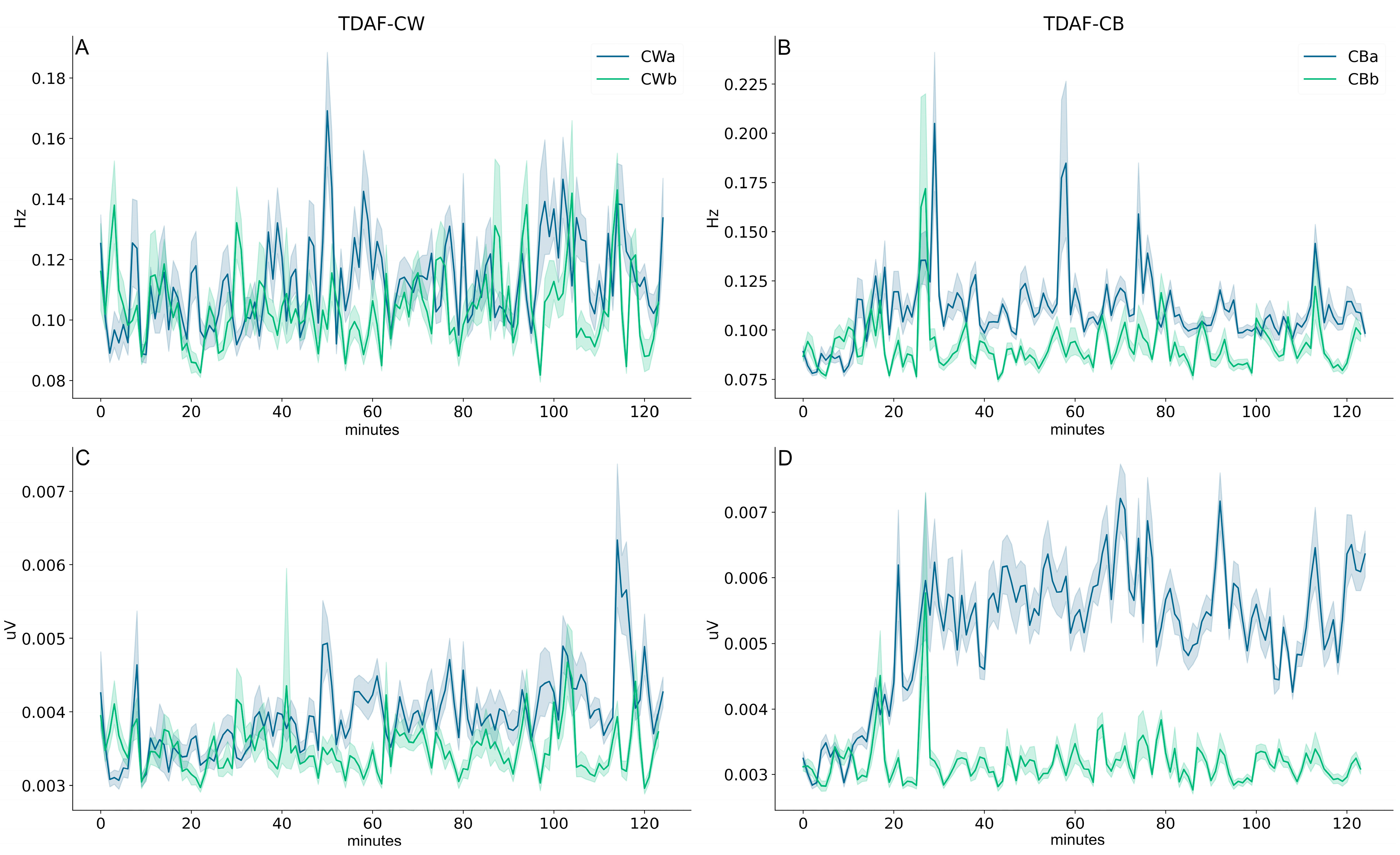
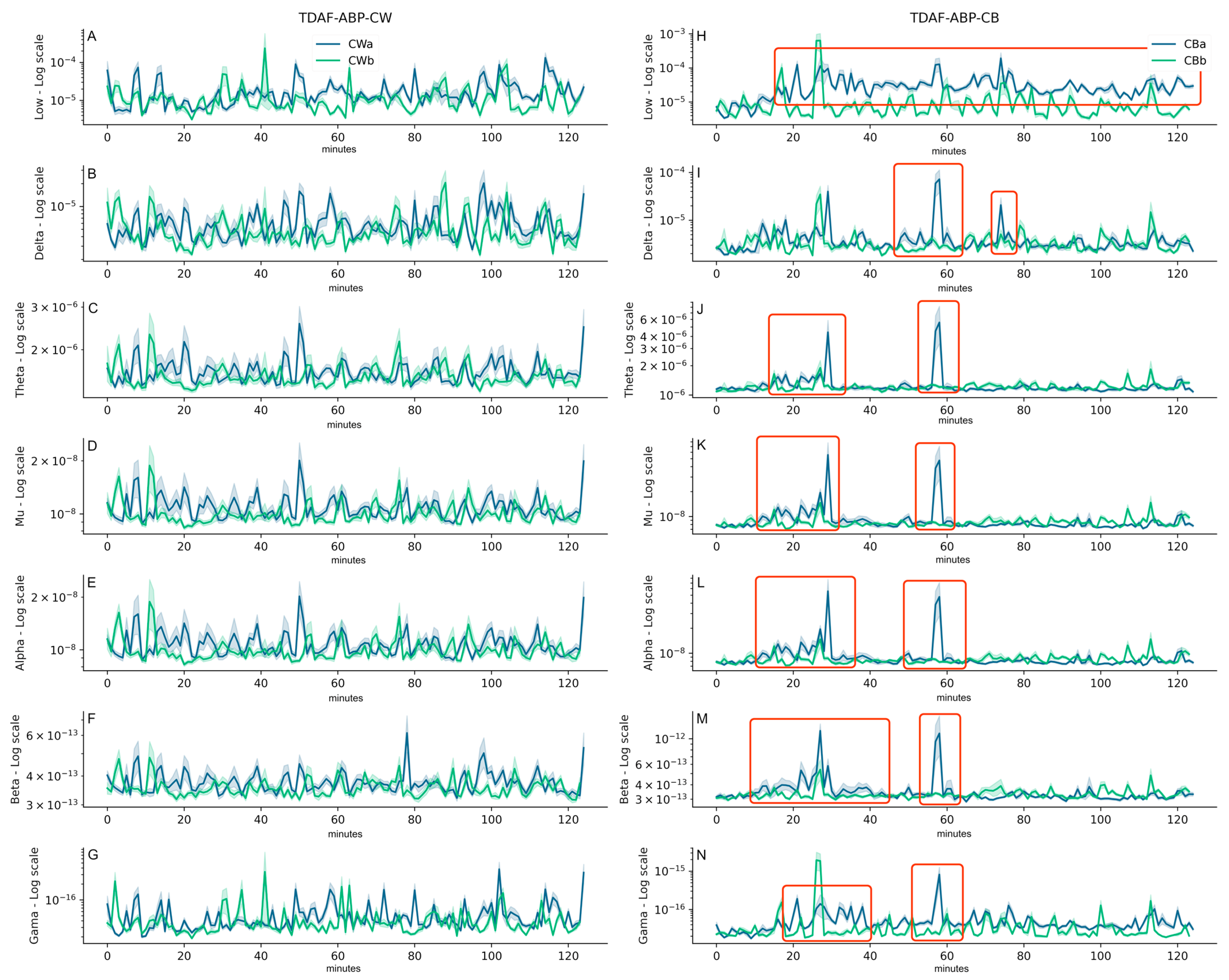
2. Results
3. Discussion
- (1)
- The complex functioning of ion channels and dipoles in cell membranes (apart from differences in the ions involved in plant and animal signals), whose direct influence on the shape of the waves has been confirmed at least in human brains with computer models [50]. How plant ion channels influence bioelectrical oscillation in plants remains to be elucidated.
- (2)
- The generation of spontaneous low-voltage bioelectrical signals (background EPG activity) through layers of tissue dipoles that correspond for brainwaves to extracellular EEG ionic currents [51,52], and intracellular electromagnetic field potentials that can be recorded by MEG [53]. Spontaneous EPGs, as part of the electrome, may synchronise when plant tissues are stimulated [23,24,54,55].
- (3)
- The nature of the signals collected in the plant tissues, ranging within the microvolt amplitude (5–250 μV) and relatively low frequencies (0.5–15 Hz)—despite being, as shown here, lower, and less diversified than in humans and non-human animals.
4. Materials and Methods
4.1. Plant Material and Experimental Setup
4.2. Electrophysiological Recordings
4.3. Electrophysiological Analyses
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neisser, U. Cognition and Reality; W. H. Freeman and Company: San Francisco, CA, USA, 1976. [Google Scholar]
- Maturana, H.R.; Varela, F.J. Autopoiesis and Cognition: The Realization of the Living; D. Reidel Publishing Company: Dordrecht, The Netherlands, 1980. [Google Scholar]
- Baluška, F.; Reber, A.S.; Miller, W.B., Jr. Cellular sentience as the primary source of biological order and evolution. BioScience 2022, 218, 104694. [Google Scholar] [CrossRef]
- James, W. The Principles of Psychology; 1981 Reprint; Harvard University Press: Cambridge, MA, USA, 1890. [Google Scholar]
- Driver, J. A selective review of selective attention research from the past century. Br. J. Psychol. 2001, 92, 53–78. [Google Scholar] [CrossRef] [PubMed]
- Chun, M.M.; Golomb, J.D.; Turk-Browne, N.B. A taxonomy of external and internal attention. Annu. Rev. Psychol. 2011, 62, 73–101. [Google Scholar] [CrossRef] [PubMed]
- Gibson, J.J. The Ecological Approach to Visual Perception: Classic Edition; Psychology Press: New York, NY, USA, 2014. [Google Scholar] [CrossRef]
- Lev-Ari, T.; Beeri, H.; Gutfreund, Y. The ecological view of selective attention. Front. Integr. Neurosci. 2022, 16, 856207. [Google Scholar] [CrossRef] [PubMed]
- Parise, A.G.; de Toledo, G.R.A.; Oliveira, T.F.d.C.; Souza, G.M.; Castiello, U.; Gagliano, M.; Marder, M. Do plants pay attention? A possible phenomenological-empirical approach. Prog. Biophys. Mol. Biol. 2022, 173, 11–23. [Google Scholar] [CrossRef]
- Karban, R. Plant Sensing and Communication; The University of Chicago Press: Chicago, IL, USA, 2014. [Google Scholar]
- Gagliano, M. Green symphonies: A call for studies on acoustic communication in plants. Behav. Ecol. 2013, 24, 789–796. [Google Scholar] [CrossRef]
- Schwartz, A.; Koller, D. Diurnal phototropism in solar tracking leaves of Lavatera cretica. Plant Physiol. 1986, 80, 771–778. [Google Scholar] [CrossRef]
- Trewavas, A. Aspects of plant intelligence. Ann. Bot. 2003, 92, 1–20. [Google Scholar] [CrossRef]
- Lüttge, U. Clusia: Holy grail and enigma. J. Exp. Bot. 2008, 59, 1503–1514. [Google Scholar] [CrossRef]
- Gagliano, M.; Vyazovskiy, V.V.; Borbély, A.A.; Grimonprez, M.; Depczynski, M. Learning by association in plants. Sci. Rep. 2016, 6, 38427. [Google Scholar] [CrossRef]
- Munné-Bosh, S. Spatiotemporal limitations in plant biology research. Trends Plant Sci. 2022, 27, P346–P354. [Google Scholar] [CrossRef] [PubMed]
- Guerra, S.; Peressotti, A.; Peressotti, F.; Bulgheroni, M.; Baccinelli, W.; D’Amico, E.; Gómez, A.; Massaccesi, S.; Ceccarini, F.; Castiello, U. Flexible control of movement in plants. Sci. Rep. 2019, 9, 16570. [Google Scholar] [CrossRef] [PubMed]
- Raja, V.; Silva, P.L.; Holghoomi, R.; Calvo, P. The dynamics of plant nutation. Sci. Rep. 2020, 10, 19465. [Google Scholar] [CrossRef]
- Wang, Q.; Guerra, S.; Bonato, B.; Simonetti, V.; Bulgheroni, M.; Castiello, U. Decision-making underlying support-searching in pea plants. Plants 2023, 12, 1597. [Google Scholar] [CrossRef]
- Marder, M. Plant intelligence and attention. Plant Signal. Behav. 2013, 8, e23902. [Google Scholar] [CrossRef] [PubMed]
- De Toledo, G.R.A.; Parise, A.G.; Simmi, F.Z.; Costa, A.V.L.; Senko, L.G.S.; Debono, M.-W.; Souza, G.M. Plant electrome: The electrical dimension of plant life. Theor. Exp. Plant Physiol. 2019, 31, 21–46. [Google Scholar] [CrossRef]
- Volkov, A. Plant Electrophysiology: Theory and Methods, 1st ed.; Springer: Berlin, Germany, 2006. [Google Scholar]
- Debono, M.-W.; Bouteau, F. Spontaneous and evoked surface potentials in Kalanchoë tissues. Life Sci. Adv. Plant. Physiol. 1992, 11, 107–117. [Google Scholar]
- Debono, M.-W. Dynamic protoneural networks in plants: A new approach of extracellular spontaneous potential variations. Plant Signal. Behav. 2013, 8, e24207. [Google Scholar] [CrossRef]
- Debono, M.-W. Perceptive levels in plants: A transdisciplinary challenge in living organism’s plasticity. Transdiscipl. J. Eng. Sci. 2013, 4, 21–39. [Google Scholar] [CrossRef]
- Souza, G.M.; Ferreira, A.S.; Saraiva, G.F.R.; Toledo, G.R.A. Plant “electrome” can be pushed toward a self-organized critical state by external cues: Evidences from a study with soybean seedlings subject to different environmental conditions. Plant Signal. Behav. 2017, 12, e1290040. [Google Scholar] [CrossRef]
- Pereira, D.R.; Papa, J.P.; Saraiva, G.F.R.; Souza, G.M. Automatic classification of plant electrophysiological responses to environmental stimuli using machine learning and interval arithmetic. Comput. Electron. Agric. 2018, 145, 35–42. [Google Scholar] [CrossRef]
- Simmi, F.; Dallagnol, L.; Ferreira, A.; Pereira, D.; Souza, G. Electrome alterations in a plant-pathogen system: Toward early diagnosis. Bioelectrochemistry 2020, 133, 107493. [Google Scholar] [CrossRef] [PubMed]
- Najdenovska, E.; Dutoit, F.; Tran, D.; Plummer, C.; Wallbridge, N.; Camps, C.; Raileanu, L.E. Classification of plant electrophysiology signals for detection of spider mites infestation in tomatoes. Appl. Sci. 2021, 11, 1414. [Google Scholar] [CrossRef]
- Gimenez, V.M.M.; Pauletti, P.M.; Silva, A.C.S.; Costa, E.J.X. Bioelectrical pattern discrimination of Miconia plants by spectral analysis and machine learning. Theor. Exp. Plant Physiol. 2021, 33, 329–342. [Google Scholar] [CrossRef]
- Parise, A.G.; Reissig, G.N.; Basso, L.F.; Senko, L.G.S.; Oliveira, T.F.d.C.; de Toledo, G.R.A.; Ferreira, A.S.; Souza, G.M. Detection of different hosts from a distance alters the behaviour and bioelectrical activity of Cuscuta racemosa. Front. Plant Sci. 2021, 12, 409. [Google Scholar] [CrossRef]
- Reissig, G.N.; Oliveira, T.F.C.; Oliveira, R.P.; Posso, D.A.; Parise, A.G.; Nava, D.E.; Souza, G.M. Fruit herbivory alters plant electrome: Evidence for fruit-shoot long-distance electrical signaling in tomato plants. Front. Sustain. Food Syst. 2021, 5, 657401. [Google Scholar] [CrossRef]
- Mokeichev, A.; Segev, R.; Ben-Shahar, O. Orientation saliency without visual cortex and target selection in archer fish. Proc. Natl. Acad. Sci. USA 2010, 107, 16726–16731. [Google Scholar] [CrossRef] [PubMed]
- Sridharan, D.; Schwarz, J.S.; Knudsen, E.I. Selective attention in birds. Curr. Biol. 2010, 24, R510–R513. [Google Scholar] [CrossRef]
- Sareen, P.; Wolf, R.; Heisenberg, M. Attracting the attention of a fly. Proc. Natl. Acad. Sci. USA 2011, 108, 7230–7235. [Google Scholar] [CrossRef]
- Paulk, A.C.; Stacey, J.A.; Pearson, T.W.; Taylor, G.J.; Moore, R.J.; Srinivasan, M.V.; van Swinderen, B. Selective attention in the honeybee optic lobes precedes behavioral choices. Proc. Natl. Acad. Sci. USA 2014, 111, 5006–5011. [Google Scholar] [CrossRef]
- Van Swinderen, B. Attention-like processes in Drosophila require short-term memory genes. Science 2007, 315, 1590–1593. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Guo, J.Z.; Peng, Y.; Xi, W.; Guo, A. Dopamine-mushroom body circuit regulates saliency-based decision-making in Drosophila. Science 2007, 316, 1901–1904. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.V.L.; Oliveira, T.F.C.; Posso, D.A.; Reissig, G.N.; Parise, A.G.; Barros, W.S.; Souza, G.M. Systemic signals induced by single and combined abiotic stimuli in common bean plants. Plants 2023, 12, 924. [Google Scholar] [CrossRef] [PubMed]
- Debono, M.-W.; Souza, G.M. Plants as electromic plastic interfaces: A mesological approach. Prog. Biophys. Mol. Biol. 2019, 146, 123–133. [Google Scholar] [CrossRef]
- Harmony, T. The functional significance of delta oscillations in cognitive processing. Front. Integr. Neurosci. 2013, 7, 0083. [Google Scholar] [CrossRef]
- De Arcangelis, L.; Herrmann, H.J. Learning as a phenomenon occurring in a critical state. Proc. Natl. Acad. Sci. USA 2010, 107, 3977–3981. [Google Scholar] [CrossRef]
- Saraiva, G.F.R.; Ferreira, A.S.; Souza, G.M. Osmotic stress decreases complexity underlying the electrophysiological dynamic in soybean. Plant Biol. 2017, 19, 702–708. [Google Scholar] [CrossRef]
- Debono, M.-W. The plant-environment interface: A mesological approach to plant cognition. In Proceedings of the Third World Congress of Transdisciplinarity, CTU Weeks, Transdisciplinary Unesco Chair “Human Development and Culture of Peace”, Florence, Italy, 13–15 October 2021. [Google Scholar]
- Debono, M.-W. Mesological plasticity as a new model to study plant evolution, interactive ecosystems & self-organized evolutionary processes. In Self-Organization as a New Paradigm in Evolutionary Biology: From Theory to Applied Cases in the Tree of Life; Dambricourt-Malassé, A., Ed.; Springer-Nature: Cham, Switzerland, 2022; Volume 5, pp. 253–290. [Google Scholar] [CrossRef]
- Simmi, F.Z.; Dallagnol, L.J.; Almeida, R.O.; Dorneles, K.R.; Souza, G.M. Barley systemic bioelectrical changes detect pathogenic infection days before the first disease symptoms. Comput. Electron. Agric. 2023, 209, 107832. [Google Scholar] [CrossRef]
- Freeman, W.J. Origin, structure, and role of background EEG activity. Part 1. Analytic amplitude. Clin. Neurophysiol. 2004, 115, 2077–2088. [Google Scholar] [CrossRef]
- Freeman, W.J. Origin, structure, and role of background EEG activity. Part 2. Analytic phase. Clin. Neurophysiol. 2004, 115, 2089–2107. [Google Scholar] [CrossRef]
- Freeman, W.J. Origin, structure, and role of background EEG activity. Part 3. Neural frame classification. Clin. Neurophysiol. 2005, 116, 1118–1129. [Google Scholar] [CrossRef]
- Reimann, M.W.; Anastassiou, C.A.; Perin, R.; Hill, S.L.; Markram, H.; Koch, C. A biophysically detailed model of neocortical local field potentials predicts the critical role of active membrane currents. Neuron 2013, 79, 375–390. [Google Scholar] [CrossRef] [PubMed]
- Nunez, P.L.; Srinivasan, R. Electric Fields of the Brain: The Neurophysics of EEG; Oxford University Press: Oxford, UK, 2006. [Google Scholar]
- Buzsáki, G.; Anastassiou, C.; Koch, C. The origin of extracellular fields and currents—EEG, ECoG, LFP and spikes. Nat. Rev. Neurosci. 2012, 13, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Amzica, F.; Lopes da Silva, F.H. Cellular substrates of brain rhythms. In Niedermeyer’s Electroencephalography: Basic Principles, Clinical Applications, and Related Fields, 7th ed.; Schomer, D.L., Lopez da Silva, F.H., Eds.; Oxford University Press: Oxford, UK, 2017; Volume 1, pp. 20–62. [Google Scholar] [CrossRef]
- Masi, E.; Ciszak, M.; Stefano, G.; Rena, L.; Azzarello, E.; Pandolfi, C.; Mugnai, S.; Baluška, F.; Arecchi, F.T.; Mancuso, S. Spatiotemporal dynamics of the electrical network activity in the root apex. Proc. Natl. Acad. Sci. USA 2009, 106, 4048–4053. [Google Scholar] [CrossRef]
- Cabral, E.F.; Pecora, P.C.; Arce, A.I.C.; Tech, A.R.B.; Costa, E.J.X. The oscillatory bioelectrical signal from plants explained by a simulated electrical model and tested using Lempel-Ziv complexity. Comput. Electron. Agric. 2011, 76, 1–5. [Google Scholar] [CrossRef]
- Kandel, E.R.; Koester, J.; Mack, S.; Siegelbaum, S. Principles of Neural Science, 6th ed.; McGraw Hill: New York, NY, USA, 2021. [Google Scholar]
- Remmers, J.E.; Gautier, H. Neural and mechanical mechanisms of feline purring. Respir. Physiol. 1972, 16, 351–361. [Google Scholar] [CrossRef]
- Da Silva, F.L. EEG: Origin and Measurement. In EEG–fMRI; Mulert, C., Lemieux, L., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 19–38. [Google Scholar] [CrossRef]
- Steriade, M. Grouping of brain rhythms in corticothalamic systems. Neuroscience 2006, 137, 1087–1106. [Google Scholar] [CrossRef] [PubMed]
- Turner, D.C.; Bateson, P. (Eds.) The Domestic Cat: The Biology of Its Behaviour, 2nd ed.; The University of Cambridge Press: Cambridge, UK, 2000. [Google Scholar]
- Canales, J.; Henriquez-Valencia, C.; Brauchi, S. The integration of electrical signals originating in the root of vascular plants. Front. Plant Sci. 2018, 8, 2173. [Google Scholar] [CrossRef]
- Baluška, F.; Mancuso, S. Deep evolutionary origins of neurobiology: Turning the essence of ‘neural’ upside-down. Commun. Integr. Biol. 2009, 2, 60–65. [Google Scholar] [CrossRef]
- Gagliano, M.; Renton, M.; Depczynski, M.; Mancuso, S. Experience teaches plants to learn faster and forget slower in environments where it matters. Oecologia 2014, 175, 63–72. [Google Scholar] [CrossRef]
- Calvo, P.; Gagliano, M.; Souza, G.M.; Trewavas, A. Plants are intelligent: Here’s how. Ann. Bot. 2020, 125, 11–28. [Google Scholar] [CrossRef]
- Calvo Garzón, P.; Keijzer, F.A. Plants: Adaptive behavior, root-brains, and minimal cognition. Adapt. Behav. 2011, 19, 155–171. [Google Scholar] [CrossRef]
- Damineli, D.S.C.; Portes, M.T.; Feijó, J.A. Electrifying rhythms in plant cells. Curr. Opin. Cell Biol. 2022, 77, 102113. [Google Scholar] [CrossRef] [PubMed]
- Ala, T.S.; Ahmadi-Pajouh, M.A.; Nasrabadi, A.M. Cumulative effects of theta binaural beats on brain power and functional connectivity. Biomed. Signal Process. Control. 2018, 42, 242–252. [Google Scholar] [CrossRef]
- Von der Malsburg, C. Synaptic plasticity as basis of brain organization. In The Neural and Molecular Bases of Learning; Changeux, J.P., Konishi, M., Eds.; John Wiley & Sons: Chichester, UK, 1987; pp. 411–431. [Google Scholar]
- Engel, A.K.; Konig, P.; Kreiter, A.K.; Gray, C.M.; Singer, W. Temporal coding by coherent oscillations as a potential solution to the binding problem. In NONLINEAR Dynamics and Neural Networks; Schuster, H.G., Ed.; VCH Pub: New York, NY, USA, 1990. [Google Scholar]
- Engel, A.K.; Singer, W. Temporal binding and the neural correlates of sensory awareness. Trends. Cogn. Sci. 2001, 5, 16–25. [Google Scholar] [CrossRef]
- Hazledine, S.; Sun, J.; Wysham, D.; Downie, J.A.; Oldroyd, G.E.D.; Morris, R.J. Nonlinear time series analysis of nodulation factor induced calcium oscillations: Evidence for deterministic chaos? PLoS ONE 2009, 4, e6637. [Google Scholar] [CrossRef]
- Sukhova, E.; Akinchits, E.; Sukhov, V. Mathematical models of electrical activity in plants. J. Membr. Biol. 2017, 250, 407–423. [Google Scholar] [CrossRef]
- Debono, M.-W. Electrome & Cognition modes in plants: A transdisciplinary approach to the eco-sensitiveness of the world. Transdiscipl. J. Eng. Sci. 2020, 11, 213–239. Available online: https://www.atlas-tjes.org/index.php/tjes/article/view/169 (accessed on 10 May 2023).
- Tian, W.; Wang, C.; Gao, Q.; Li, L.; Luan, S. Calcium spikes, waves and oscillations in plant development and biotic interactions. Nat. Plants 2020, 6, 750–759. [Google Scholar] [CrossRef]
- Amzica, F.; Steriade, M. The K-complex: Its slow (<1-Hz) rhythmicity and relation to delta waves. Neurology 1997, 49, 952–959. [Google Scholar] [CrossRef]
- Neske, G.T. The slow oscillation in cortical and thalamic networks: Mechanisms and functions. Front. Neural Circuits 2015, 9, 88. [Google Scholar] [CrossRef] [PubMed]
- Achermann, P.; Borbély, A.A. Low-frequency (<1 Hz) oscillations in the human sleep electroencephalogram. Neuroscience 1997, 81, 213–222. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, L.F.C.; Oliveira, M.G.C.; Wendland, A.; Heinemann, A.B.; Guimarães, C.M.; Ferreira, E.P.B.; Quintela, E.D.; Barbosa, F.R.; Carvalho, M.C.S.; Lobo Junior, M.; et al. Conhecendo a Fenologia do Feijoeiro e Seus Aspectos Fitotécnicos; Embrapa: Brasília, DF, Brazil, 2018. [Google Scholar]
- Costea, M.; Tardif, F.J. The biology of Canadian weeds. 133. Cuscuta campestris Yuncker, C. gronovii Willd. ex Schult., C. umbrosa Beyr. ex Hook., C. epithymum (L.) L. and C. epilinum Weihe. Can. J. Plant Sci. 2006, 86, 293–316. [Google Scholar] [CrossRef]
- Runyon, J.B.; Mescher, M.C.; de Moraes, C.M. Volatile chemical cues guide host location and host selection by parasitic plants. Science 2006, 313, 1964–1967. [Google Scholar] [CrossRef]
- Albert, M.; Belastegui-Macadam, X.; Bleischwitz, M.; Kaldenhoff, R. Cuscuta spp: “parasitic plants in the spotlight of plant physiology, economy and ecology”. In Progress in Botany; Lüttge, U., Beyschlag, W., Murata, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 267–277. [Google Scholar] [CrossRef]
- Bluestein, L.A. Linear filtering approach to the computation of discrete Fourier transform. IEEE Trans. Audio Electroacoust. 1970, 18, 451–455. [Google Scholar] [CrossRef]
- Virtanen, P.; Gommers, R.; Oliphant, T.E.; Haberland, M.; Reddy, T.; Cournapeau, D.; Burovski, E.; Peterson, P.; Weckesser, W.; Bright, J.; et al. SciPy 1.0: Fundamental algorithms for scientific computing in Python. Nat. Methods 2020, 17, 261–272. [Google Scholar] [CrossRef]
- Welch, P. The use of fast Fourier transform for the estimation of power spectra: A method based on time averaging over short, modified periodograms. IEEE Trans. Audio Electroacoust. 1967, 15, 70–73. [Google Scholar] [CrossRef]
- Evans, J.R. Neurofeedback. In Encyclopedia of the Human Brain; Elsevier: Amsterdam, The Netherlands, 2002; pp. 465–477. [Google Scholar]
- Heraz, A.; Razaki, R.; Frasson, C. Using machine learning to predict learner emotional state from brainwaves. In Proceedings of the Seventh IEEE International Conference on Advanced Learning Technologies (ICALT 2007), Niigata, Japan, 18–20 July 2007. [Google Scholar] [CrossRef]
- Abhang, P.A.; Gawali, B.W.; Mehrotra, S.C. Technical aspects of brain rhythms and speech parameters. In Introduction to EEG- and Speech-Based Emotion Recognition; Abhang, P.A., Gawali, B.W., Mehrotra, S.C., Eds.; Elsevier: London, UK, 2016; pp. 51–79. [Google Scholar]
- Abhang, P.A.; Gawali, B.W.; Mehrotra, S.C. Technological basics of EEG recording and operation of apparatus. In Introduction to EEG- and Speech-Based Emotion Recognition; Abhang, P.A., Gawali, B.W., Mehrotra, S.C., Eds.; Elsevier: London, UK, 2016; pp. 19–50. [Google Scholar]
- Delimayanti, M.K.; Purnama, B.; Nguyen, N.G.; Faisal, M.R.; Mahmudah, K.R.; Indriani, F.; Kubo, M.; Satou, K. Classification of brainwaves for sleep stages by high-dimensional FFT characteristics from EEG signals. Appl. Sci. 2020, 10, 1797. [Google Scholar] [CrossRef]
- Savadkoohi, M.; Oladunni, T.; Thompson, L. A machine learning approach to epileptic seizure prediction using electroencephalogram (EEG) signal. Biocybern. Biomed. Eng. 2020, 40, 1328–1341. [Google Scholar] [CrossRef]
- Kora, P.; Meenakshi, K.; Swaraja, K.; Rajani, A.; Raju, M.S. EEG based interpretation of human brain activity during yoga and meditation using machine learning: A systematic review. Complement. Ther. Clin. Pract. 2021, 43, 101329. [Google Scholar] [CrossRef]
- Zhong, G. Analysis of healthy people’s attention based on EEG spectrum. In Proceedings of the 2015 International Conference on Mechatronics, Electronic, Industrial and Control Engineering, Shenyang, China, 1–3 April 2015. [Google Scholar] [CrossRef]
- Kirmizi-Alsan, E.; Bayraktaroglu, Z.; Guruvit, H.; Keskin, Y.H.; Emre, M.; Demiralp, T. Comparative analysis of event-related potentials during Go/NoGo and CPT: Decomposition of electrophysiological markers of response inhibition and sustained attention. Brain Res. 2006, 1104, 114–128. [Google Scholar] [CrossRef] [PubMed]
- Karamacoska, D.; Barry, R.J.; De Blasio, F.M.; Steiner, G.Z. EEG-ERP dynamics in a visual continuous performance test. Int. J. Psychophysiol. 2019, 146, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Press, W.H.; Flannery, B.P.; Teukolsky, S.A.; Vetterling, W.T. Numerical Recipes in Pascal: The Art of Scientific Computing; Cambridge University Press: Cambridge, UK, 1989. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parise, A.G.; Oliveira, T.F.d.C.; Debono, M.-W.; Souza, G.M. The Electrome of a Parasitic Plant in a Putative State of Attention Increases the Energy of Low Band Frequency Waves: A Comparative Study with Neural Systems. Plants 2023, 12, 2005. https://doi.org/10.3390/plants12102005
Parise AG, Oliveira TFdC, Debono M-W, Souza GM. The Electrome of a Parasitic Plant in a Putative State of Attention Increases the Energy of Low Band Frequency Waves: A Comparative Study with Neural Systems. Plants. 2023; 12(10):2005. https://doi.org/10.3390/plants12102005
Chicago/Turabian StyleParise, André Geremia, Thiago Francisco de Carvalho Oliveira, Marc-Williams Debono, and Gustavo Maia Souza. 2023. "The Electrome of a Parasitic Plant in a Putative State of Attention Increases the Energy of Low Band Frequency Waves: A Comparative Study with Neural Systems" Plants 12, no. 10: 2005. https://doi.org/10.3390/plants12102005
APA StyleParise, A. G., Oliveira, T. F. d. C., Debono, M. -W., & Souza, G. M. (2023). The Electrome of a Parasitic Plant in a Putative State of Attention Increases the Energy of Low Band Frequency Waves: A Comparative Study with Neural Systems. Plants, 12(10), 2005. https://doi.org/10.3390/plants12102005