
Integrating BLUP, AMMI, and GGE Models to Explore GE Interactions for Adaptability and Stability of Winter Lentils (Lens culinaris Medik.)

 ,
,  ,
,  ,
,  ,
,  , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Stability Analysis for Different Yield Contributing Characters
2.2. AMMI Analysis of Variance
2.3. Additive Main Effects and Multiplicative Interaction 1 (AMMI1)
2.4. Additive Main Effects and Multiplicative Interaction 2 (AMMI2)
2.5. Non-Parametric Measures
2.6. AMMI Stability Measure
2.7. Parametric Stability Measures
2.8. Simultaneous Selection Indices (SSI) of Genotypes for Yield
2.9. Best Linear Unbiased Prediction-Based Stability Indices (BLUP)
2.10. Correlation between Stability Statistics
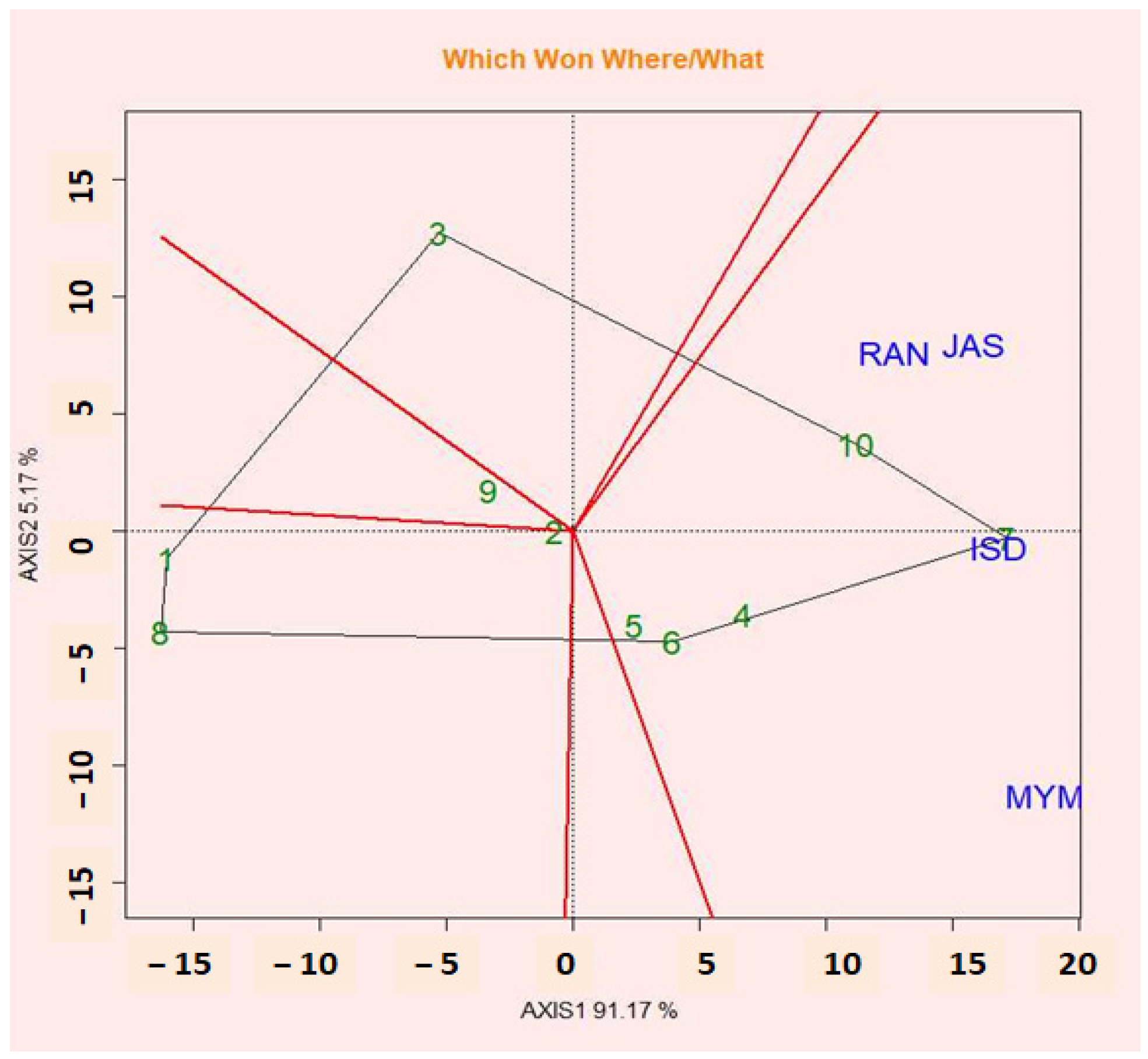
2.11. Polygon View of GGE Biplot (Which-Won-Where Pattern)
2.12. Mean vs. Stability Views of GGE Biplot
2.13. Evaluation of Genotypes Relative to Ideal Genotypes
2.14. Evaluation of Environments Relative to Ideal Environments
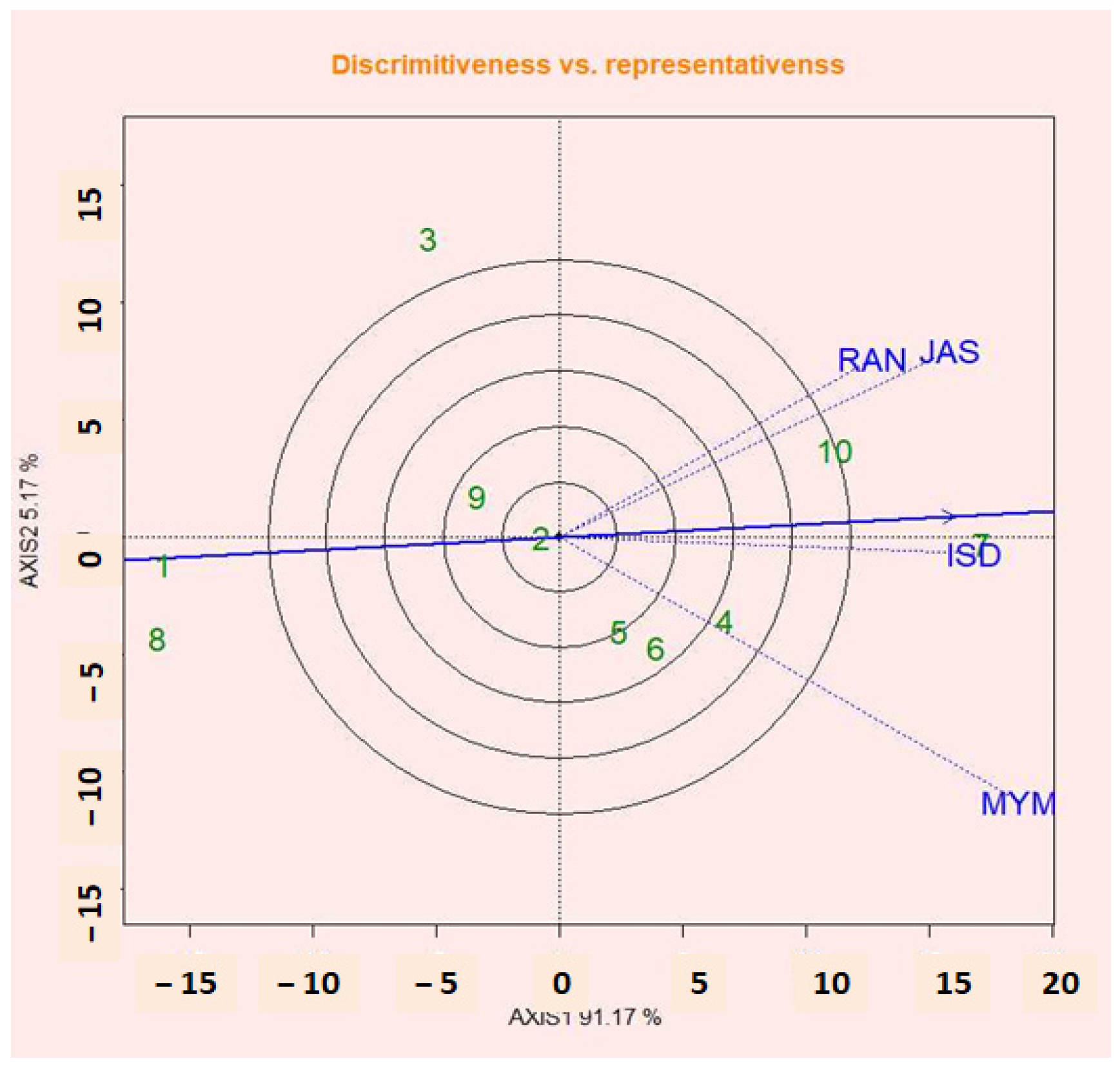
2.15. Discrimination Abilities vs. Representation of Environments
3. Discussions
4. Materials and Methods
4.1. Planting Sites and Plant Materials
4.2. Experimental Design and Data Collection
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Negussie, T.; Pretorius, Z.A. Lentil Rust: Present Status and Future Prospects. Crop Prot. 2012, 32, 119–128. [Google Scholar] [CrossRef]
- Parihar, A.K.; Basandrai, A.K.; Saxena, D.R.; Kushwaha, K.P.S.; Chandra, S.; Sharma, K.; Singha, K.D.; Singh, D.; Lal, H.C.; Gupta, S. Biplot Evaluation of Test Environments and Identification of Lentil Genotypes with Durable Resistance to Fusarium Wilt in India. Crop Pasture Sci. 2017, 68, 1024–1030. [Google Scholar] [CrossRef]
- Arumuganathan, K.; Earle, E.D. Nuclear DNA Content of Some Important Plant Species. Plant Mol. Biol. Report. 1991, 9, 208–218. [Google Scholar] [CrossRef]
- Shrestha, R.; Rizvi, A.H.; Sarker, A.; Darai, R.; Paneru, R.B.; Vandenberg, A.; Singh, M. Genotypic Variability and Genotype× Environment Interaction for Iron and Zinc Content in Lentil under Nepalese Environments. Crop Sci. 2018, 58, 2503–2510. [Google Scholar] [CrossRef]
- Grusak, M.A. Nutritional and Health-Beneficial Quality. Lentil Bot. Prod. Uses 2009, 1418, 368. [Google Scholar]
- Anonymous. Agriculture Information Service (AIS); Department of Agricultural Extension, Ministry of Agriculture, Government of the People’s Republic of Bangladesh: Dhaka, Bangladesh, 2020.
- Martey, E.; Wiredu, A.N.; Oteng-Frimpong, R. Baseline Study of Groundnut in Northern Ghana; LAP Lambert Academic Publishing: Saarbrücken, Germany, 2015. [Google Scholar]
- McGuire, S.; Sperling, L. Seed Systems Smallholder Farmers Use. Food Secur. 2016, 8, 179–195. [Google Scholar] [CrossRef]
- Hasan, M.J.; Kulsum, M.U.; Sarker, U.; Matin, M.Q.I.; Shahin, N.H.; Kabir, M.S.; Ercisli, S.; Marc, R.A. Assessment of GGE, AMMI, Regression, and Its Deviation Model to Identify Stable Rice Hybrids in Bangladesh. Plants 2022, 11, 2336. [Google Scholar] [CrossRef]
- Sarker, U.; Biswas, P.S.; Prasad, B. Stability for Grain Yield and Yield Components in Rice (Oryza sativa L.). Bangladesh J. Agril. Res. 2007, 32, 559–564. [Google Scholar]
- Islam, M.A.; Johora, F.T.; Sarker, U.; Mian, M.A.K. Adaptation of Chinese CMS Lines Interaction with Seedling Age and Row Ratio on Hybrid Seed Production of Rice (Oryza sativa L.). Bangladesh J. Agron. 2012, 25, 178–183. [Google Scholar]
- Islam, M.A.; Sarker, U.; Hasan, M.R.; Islam, M.R.; Mahmud, M.N.H. Genetype × Environment (Fertilizer Dose) Interaction and Stability Analysis of Hybrid Seed Production of Rice (Oryza sativa L.). Eco-Friendly Agric. J. 2012, 5, 183–185. [Google Scholar]
- Islam, M.A.; Sarker, U.; Mian, M.A.K.; Ahmed, J.U. Genotype Fertilizer Doses Interaction for Hybrid Seed Yield of Rice (Oryza sativa L.). Bangladesh J. Plant Breed. Genet. 2011, 24, 41–44. [Google Scholar] [CrossRef]
- Islam, M.A.; Mian, M.A.K.; Rasul, G.; Johora, F.T.; Sarker, U. Interaction Effect between Genotypes, Row Ratio and Fertilizer Dose on Hybrid Seed Production of Rice (Oryza sativa L.). In Proceedings of the 10th Conference Proceeding of Bangladesh Society of Agronomy, Gazipur, Bangladesh, 8 October 2011; pp. 134–141. [Google Scholar]
- Islam, M.A.; Sarker, U.; Mian, M.A.K.; Ahmed, J.U. Genotype Seedling Age Interaction for Hybrid Seed Yield of Rice (Oryza sativa L.). Bangladesh J. Plant Breed. Genet. 2011, 24, 23–26. [Google Scholar] [CrossRef]
- Sarker, U. Stability for Grain Yield under Different Planting Times in Rice. Bangladesh J. Agril. Res. 2002, 27, 425–430. [Google Scholar]
- Sarker, U.; Ferdous, N. Genotype× seedling age interaction in rice (Oryza sativa L.). Pak. J. Biol. Sci. 2002, 5, 275–277. [Google Scholar] [CrossRef]
- Kulsum, M.U.; Sarker, U.; Karim, M.A.; Mian, M.A.K. Additive Main Effects and Multiplicative Interaction (AMMI) Analysis for Yield of Hybrid Rice in Bangladesh. Trop. Agric. Dev. 2012, 6, 53–61. [Google Scholar]
- Wricke, G. Über eine Methode zur Erfassung der ökologischen Streubreite in Feldversuchen. Z. Pflanzenzüchtg 1962, 47, 9296. [Google Scholar]
- Eberhart, S.A.; Russell, W.A. Stability Parameters for Comparing Varieties 1. Crop Sci. 1966, 6, 36–40. [Google Scholar] [CrossRef]
- Finlay, K.W.; Wilkinson, G.N. The Analysis of Adaptation in a Plant-Breeding Programme. Aust. J. Agric. Res. 1963, 14, 742–754. [Google Scholar] [CrossRef]
- Shukla, G.K. Some Statistical Aspects of Partitioning Genotype-Environmental Components of Variability. Heredity 1972, 29, 237–245. [Google Scholar] [CrossRef]
- Francis, T.R.; Kannenberg, L.W. Yield Stability Studies in Short-Season Maize. I. A Descriptive Method for Grouping Genotypes. Can. J. Plant Sci. 1978, 58, 1029–1034. [Google Scholar] [CrossRef]
- Purchase, J.L. Parametric Analysis to Describe Genotype× Environment Interaction and Yield Stability in Winter Wheat. Ph.D. Thesis, University of the Free State, Bloemfontein, South Africa, 1997. [Google Scholar]
- Purchase, J.L.; Hatting, H.; Van Deventer, C.S. Genotype× Environment Interaction of Winter Wheat (Triticum aestivum L.) in South Africa: II. Stability Analysis of Yield Performance. S. Afr. J. Plant Soil 2000, 17, 101–107. [Google Scholar] [CrossRef]
- Huehn, M. Nonparametric Measures of Phenotypic Stability. Part 1: Theory. Euphytica 1990, 47, 189–194. [Google Scholar] [CrossRef]
- Huehn, M. Beiträge zur Erfassung der phänotypischen Stabilität. EDV Med. Biol. 1979, 10, 112–117. [Google Scholar]
- Nassar, R.; Huehn, M. Studies on Estimation of Phenotypic Stability: Tests of Significance for Nonparametric Measures of Phenotypic Stability. Biometrics 1987, 43, 45–53. [Google Scholar] [CrossRef]
- Thennarasu, K. On Certain Non-Parametric Procedures for Studying Genotype-Environment Interactions and Yield Stability; IARI, Division of Agricultural Statistics: New Delhi, India, 1995.
- Vaezi, B.; Pour-Aboughadareh, A.; Mohammadi, R.; Mehraban, A.; Hossein-Pour, T.; Koohkan, E.; Ghasemi, S.; Moradkhani, H.; Siddique, K.H.M. Integrating Different Stability Models to Investigate Genotype× Environment Interactions and Identify Stable and High-Yielding Barley Genotypes. Euphytica 2019, 215, 63. [Google Scholar] [CrossRef]
- Malosetti, M.; Ribaut, J.-M.; van Eeuwijk, F.A. The Statistical Analysis of Multi-Environment Data: Modeling Genotype-by-Environment Interaction and Its Genetic Basis. Front. Physiol. 2013, 4, 44. [Google Scholar] [CrossRef]
- Mukherjee, A.K.; Mohapatra, N.K.; Bose, L.K.; Jambhulkar, N.N.; Nayak, P. Additive Main Effects and Multiplicative Interaction (AMMI) Analysis of G x E Interactions in Rice-Blast Pathosystem to Identify Stable Resistant Genotypes. Afr. J. Agric. Res. 2013, 8, 5492–5507. [Google Scholar]
- Tekalign, A.; Sibiya, J.; Derera, J.; Fikre, A. Analysis of Genotype× Environment Interaction and Stability for Grain Yield and Chocolate Spot (Botrytis Fabae) Disease Resistance in Faba Bean (Vicia Faba). Aust. J. Crop Sci. 2017, 11, 1228–1235. [Google Scholar] [CrossRef]
- Ghazvini, H.; Pour-Aboughadareh, A.; Sharifalhosseini, M.; Razavi, S.A.; Mohammadi, S.; Ghasemi Kalkhoran, M.; Fathi Hafshejani, A.; Khakizadeh, G. Phenotypic Stability Analysis of Barley Promising Lines in the Cold Regions of Iran. Crop Breed. J. 2018, 8, 17–29. [Google Scholar]
- Crossa, J. Statistical Analyses of Multilocation Trials. Adv. Agron. 1990, 44, 55–85. [Google Scholar]
- Yan, W.; Hunt, L.A.; Sheng, Q.; Szlavnics, Z. Cultivar Evaluation and Mega-Environment Investigation Based on the GGE Biplot. Crop Sci. 2000, 40, 597–605. [Google Scholar] [CrossRef]
- Yan, W.; Kang, M.S. GGE Biplot Analysis: A Graphical Tool for Breeders, Geneticists, and Agronomists; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Rao, A.R.; Prabhakaran, V.T. Use of AMMI in Simultaneous Selection of Genotypes for Yield and Stability. J. Ind. Soc. Agri. Statis. 2005, 59, 76–82. [Google Scholar]
- Farshadfar, E. Incorporation of AMMI Stability Value and Grain Yield in a Single Non-Parametric Index (GSI) in Bread Wheat. Pak. J. Biol. Sci. 2008, 11, 1791. [Google Scholar] [CrossRef] [PubMed]
- Verardi, S.; Dahourou, D.; Ah-Kion, J.; Bhowon, U.; Tseung, C.N.; Amoussou-Yeye, D.; Adjahouisso, M.; Bouatta, C.; Dougoumalé Cissé, D.; Mbodji, M. Psychometric Properties of the Marlowe-Crowne Social Desirability Scale in Eight African Countries and Switzerland. J. Cross. Cult. Psychol. 2010, 41, 19–34. [Google Scholar] [CrossRef]
- De Resende, M.D.V. SELEGEN-REML/BLUP: Sistema Estatístico e Seleção Genética Computadorizada via Modelos Lineares Mistos; Embrapa Florestas: Colombo, Brasil, 2007. [Google Scholar]
- Yan, W. Mega-environment Analysis and Test Location Evaluation Based on Unbalanced Multiyear Data. Crop Sci. 2015, 55, 113–122. [Google Scholar] [CrossRef]
- Gauch, H.G.; Piepho, H.P.; Annicchiarico, P. Statistical Analysis of Yield Trials by AMMI and GGE: Further Considerations. Crop Sci. 2008, 48, 866–889. [Google Scholar] [CrossRef]
- Yan, W.; Holland, J.B. A Heritability-Adjusted GGE Biplot for Test Environment Evaluation. Euphytica 2010, 171, 355–369. [Google Scholar] [CrossRef]
- Da Cruz, D.P.; de Amaral Gravina, G.; Vivas, M.; Entringer, G.C.; Rocha, R.S.; da Costa Jaeggi, M.E.P.; Gravina, L.M.; Pereira, I.M.; do Amaral Junior, A.T.; de Moraes, R. Analysis of the Phenotypic Adaptability and Stability of Strains of Cowpea through the GGE Biplot Approach. Euphytica 2020, 216, 160. [Google Scholar] [CrossRef]
- Singh, B.; Das, A.; Parihar, A.K.; Bhagawati, B.; Singh, D.; Pathak, K.N.; Dwivedi, K.; Das, N.; Keshari, N.; Midha, R.L.; et al. Delineation of Genotype-by-Environment Interactions for Identification and Validation of Resistant Genotypes in Mungbean to Root-Knot Nematode (Meloidogyne Incognita) Using GGE Biplot. Sci. Rep. 2020, 10, 4108. [Google Scholar] [CrossRef]
- Biswas, A.; Sarker, U.; Banik, B.R.; Rohman, M.M.; Talukder, M.Z.A. Genotype × environment interaction for grain yield of maize (Zea mays L.) inbreds under salinity stress. Bangladesh J. Agril. Res. 2014, 39, 293–301. [Google Scholar] [CrossRef]
- Zobel, R.W.; Wright, M.J.; Gauch, H.G., Jr. Statistical Analysis of a Yield Trial. Agron. J. 1988, 80, 388–393. [Google Scholar] [CrossRef]
- Alam, M.A.; Sarker, Z.I.; Farhad, M.; Hakim, M.A.; Barma, N.C.D.; Hossain, M.I.; Rahman, M.M.; Islam, R. Yield Stability of Newly Released Wheat Varieties in Multi-Environments of Bangladesh. Int. J. Plant Soil Sci. 2015, 6, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Akter, A.; Jamil, H.M.; Umma, K.M.; Islam, M.R.; Hossain, K.; Mamunur, R.M. AMMI Biplot Analysis for Stability of Grain Yield in Hybrid Rice (Oryza sativa L.). J. Rice Res. 2014, 2, 126. [Google Scholar] [CrossRef]
- Azam, M.G.; Iqbal, M.S.; Hossain, M.A.; Hossain, M.F. Stability Investigation and Genotype× Environment Association in Chickpea Genotypes Utilizing AMMI and GGE Biplot Model. Genet. Mol. Res. 2020, 19, gmr16039980. [Google Scholar]
- Murphy, S.E.; Lee, E.A.; Woodrow, L.; Seguin, P.; Kumar, J.; Rajcan, I.; Ablett, G.R. Genotype× Environment Interaction and Stability for Isoflavone Content in Soybean. Crop Sci. 2009, 49, 1313–1321. [Google Scholar] [CrossRef]
- Farshadfar, E.; Zali, H.; Mohammadi, R. Evaluation of Phenotypic Stability in Chickpea Genotypes Using GGE-Biplot. Ann. Biol. Res. 2011, 2, 282–292. [Google Scholar]
- Zali, H.; Farshadfar, E.; Sabaghpour, S.H.; Karimizadeh, R. Evaluation of Genotype× Environment Interaction in Chickpea Using Measures of Stability from AMMI Model. Ann. Biol. Res. 2012, 3, 3126–3136. [Google Scholar]
- Yan, W.; Kang, M.S.; Ma, B.; Woods, S.; Cornelius, P.L. GGE Biplot vs. AMMI Analysis of Genotype-by-Environment Data. Crop Sci. 2007, 47, 643–653. [Google Scholar] [CrossRef]
- Yan, W.; Rajcan, I. Biplot Analysis of Test Sites and Trait Relations of Soybean in Ontario. Crop Sci. 2002, 42, 11–20. [Google Scholar] [CrossRef]
- Yan, W.; Tinker, N.A. Biplot Analysis of Multi-Environment Trial Data: Principles and Applications. Can. J. Plant Sci. 2006, 86, 623–645. [Google Scholar] [CrossRef]
- Solonechnyi, P.; Vasko, N.; Naumov, A.; Solonechnaya, O.; Vazhenina, O.; Bondareva, O.; Logvinenko, Y. GGE Biplot Analysis of Genotype by Environment Interaction of Spring Barley Varieties. Zemdirb. Agric. 2015, 102, 431–436. [Google Scholar] [CrossRef]
- Alam, M.A.; Farhad, M.; Hakim, M.A.; Barma, N.C.D.; Malaker, P.K.; Reza, M.M.A.; Hossain, M.A.; Li, M. AMMI and GGE Biplot Analysis for Yield Stability of Promising Bread Wheat Genotypes in Bangladesh. Pak. J. Bot. 2017, 49, 1049–1056. [Google Scholar]
- Sousa, M.B.E.; Damasceno-Silva, K.J.; Rocha, M.D.M.; Nogueira De Menezes Júnior, J.Â.; Lima, L.R.L. Genotype by Environment Interaction in Cowpea Lines Using GGE Biplot Method. Rev. Caatinga 2018, 31, 64–71. [Google Scholar] [CrossRef]
- Fayeun, L.S.; Alake, G.C.; Akinlolu, A.O. GGE Biplot Analysis of Fluted Pumpkin (Telfairia Occidentalis) Landraces Evaluated for Marketable Leaf Yield in Southwest Nigeria. J. Saudi Soc. Agric. Sci. 2018, 17, 416–423. [Google Scholar] [CrossRef]
- Tolessa, T.T.; Gela, T.S. Sites Regression GGE Biplot Analysis of Haricot Bean (Phaseolus Vulgaris L.) Genotypes in Three Contrasting Environments. World J. Agric. Res. 2014, 2, 228–236. [Google Scholar] [CrossRef]
- Azam, M.G.; Hossain, M.A.; Sarker, U.; Alam, A.K.M.M.; Nair, R.M.; Roychowdhury, R.; Ercisli, S.; Golokhvast, K.S. Genetic Analyses of Mungbean [Vigna radiata (L.) Wilczek] Breeding Traits for Selecting Superior Genotype(s) Using Multivariate and Multi-Traits Indexing Approaches. Plants 2023, 12, 1984. [Google Scholar] [CrossRef]
- Azam, M.D.G.; Sarker, U.; Uddin, M.D.S. Screening maize (Zea mays L.) genotypes for phosphorus deficiency at the seedling stage. Turk. J. Agric. For. 2022, 46, 802–821. [Google Scholar] [CrossRef]
- Azam, M.G.; Sarker, U.; Maniruzzam; Banik, B.R. Genetic variability of yield and its contributing characters of CIMMYT maize inbreds under drought stress. Bangladesh J. Agri. Res. 2014, 39, 419–426. [Google Scholar] [CrossRef]
- Biswas, A.; Sarker, U.; Banik, B.R.; Rohman, M.M.; Mian, M.A.K. Genetic Divergence Study in Salinity Stress Resistant Maize (Zea mays L.). Bangladesh J. Agric. Res. 2014, 39, 621–630. [Google Scholar] [CrossRef]
- Azam, M.G.A.; Sarker, U.; Mian, M.A.K.; Banik, B.R.; Talukder, M.Z.A. Genetic Divergence on Quantitative Characters of Exotic Maize Inbreds (Zea mays L.). Bangladesh J. Plant Breed. Genet. 2013, 26, 9–14. [Google Scholar] [CrossRef]
- Faysal, A.S.M.; Ali, L.; Azam, M.G.; Sarker, U.; Ercisli, S.; Golokhvast, K.S.; Marc, R.A. Genetic Variability, Character Association, and Path Coefficient Analysis in Transplant Aman Rice Genotypes. Plants 2022, 11, 2952. [Google Scholar] [CrossRef] [PubMed]
- Kulsum, U.; Sarker, U.; Rasul, M. Genetic variability, heritability and interrelationship in salt-tolerant lines of T. aman rice. Genetika 2022, 54, 761–776. [Google Scholar] [CrossRef]
- Hasan-Ud-Daula, M.; Sarker, U. Variability, Heritability, Character Association, and Path Coefficient Analysis in Advanced Breeding Lines of Rice (Oryza sativa L.). Genetika 2020, 52, 711–726. [Google Scholar] [CrossRef]
- Ali, M.A.; Sarker, U.; MAK, M.; Islam, M.A.; Johora, F.T. Estimation of Genetic Divergence in Boro Rice (Oryza sativa L.). Int. J. BioRes. 2014, 16, 28–36. [Google Scholar]
- Rai, P.K.; Sarker, U.; Roy, P.C.; Islam, A. Character Association in F4 Generation of Rice (Oryza sativa L.). Bangladesh J. Plant Breed. Genet. 2013, 26, 39–44. [Google Scholar] [CrossRef]
- Hasan, M.R.; Sarker, U.; Hossain, M.A.; Huda, K.M.K.; Mian, M.A.K.; Hossain, T.; Zahan, M.S.; Mahmud, M.N.H. Genetic Diversity in Micronutrient Dense Rice and Its Implication in Breeding Program. Eco-Friendly Agril. J. 2012, 5, 168–174. [Google Scholar]
- Hasan, M.R.; Sarker, U.; Mian, M.A.K.; Hossain, T.; Mahmud, M.N.H. Genetic Variation in Micronutrient Dense Rice and Its Implication in Breeding for Higher Yield. Eco-Friendly Agril. J. 2012, 5, 175–182. [Google Scholar]
- Siddique, M.N.A.; Sarker, U.; Mian, M.A.K. Genetic diversity in restorer line of rice. In Proceedings of the International Conference on Plant Breeding and Seed for Food Security; Bhuiyan, M.S.R., Rahman, L., Eds.; Plant Breeding and Genetics Society of Bangladesh: Dhaka, Bangladesh, 2009; pp. 137–142. [Google Scholar]
- Nath, J.K.; Sarker, U.; Mian, M.A.K.; Hossain, T. Genetic Divergence in T. aman Rice. Ann. Bangladesh Agric. 2008, 12, 51–60. [Google Scholar]
- Rahman, M.H.; Sarker, U.; Main, M.A.K. Assessment of Variability of Floral and Yield Traits; I Restorer Lines of Rice. Ann. Bangladesh Agric. 2007, 11, 87–94. [Google Scholar]
- Rahman, M.H.; Sarker, U.; Main, M.A.K. Assessment of Variability of Floral and Yield Traits; II Maintainer Lines of Rice. Ann. Bangladesh Agric. 2007, 11, 95–102. [Google Scholar]
- Ganapati, R.K.; Rasul, M.G.; Mian, M.A.K. Genetic Variability and Character Association of T-Aman Rice (Oryza sativa L.). Intl. J. Plant Biol. Res. 2014, 2, 1–4. [Google Scholar]
- Sarker, U.; Mian, M.A.K. Genetic Variations and Correlations between Floral Traits in Rice. Bangladesh J. Agril. Res 2004, 29, 553–558. [Google Scholar]
- Biswas, P.S.; Sarker, U.; Bhuiyan, M.A.R.; Khatun, S. Genetic Divergence in Cold Tolerant Irrigated Rice (Oryza sativa L.). Agriculturists 2006, 4, 15–20. [Google Scholar]
- Sarker, U.; Biswas, P.S.; Prasad, B.; Mian, M.A.K. Correlated Response, Relative Selection Efficiency and Path Analysis in Cold Tolerant Rice. Bangladesh J. Pl. Breed. Genet. 2001, 14, 33–36. [Google Scholar]
- Sarker, U.; Mian, M.A.K. Genetic Variability, Character Association and Path Analysis for Yield and Its Components in Rice. J. Asiat. Soc. Bangladesh Sci. 2003, 29, 47–54. [Google Scholar]
- Karim, D.; Siddique, M.N.A.; Sarkar, U.; Hasnat, Z.; Sultana, J. Phenotypic and Genotypic Correlation Co-Efficient of Quantitative Characters and Character Association of Aromatic Rice. J. Biosci. Agric. Res. 2014, 1, 34–46. [Google Scholar] [CrossRef]
- Ashraf, A.T.M.; Rahman, M.M.; Hossain, M.M.; Sarker, U. Study of Correlation and Path Analysis in the Selected Okra Genotypes. Asian Res. J. Agric. 2020, 12, 1–11. [Google Scholar] [CrossRef]
- Ashraf, A.T.M.; Rahman, M.M.; Hossain, M.M.; Sarker, U. Study of the Genetic Analysis of Some Selected Okra Genotypes. Int. J. Adv. Res. 2020, 8, 549–556. [Google Scholar] [CrossRef]
- Ashraf, A.T.M.; Rahman, M.M.; Hossain, M.M.; Sarker, U. Performance Evaluation of Some Selected Okra Genotypes. Int. J. Plant Soil Sci. 2020, 32, 13–20. [Google Scholar] [CrossRef]
- Rashad, M.M.I.; Sarker, U. Genetic variations in yield and yield contributing traits of green amaranth. Genetika 2020, 52, 393–407. [Google Scholar] [CrossRef]
- Sarker, U.; Azam, M.; Talukder, M.; Alam, Z. Genetic variation in mineral profiles, yield contributing agronomic traits, and foliage yield of stem amaranth. Genetika 2022, 54, 91–108. [Google Scholar] [CrossRef]
- Kayesh, E.; Sharker, M.S.; Roni, M.S.; Sarker, U. Integrated Nutrient Management for Growth, Yield and Profitability of Broccoli. Bangladesh J. Agric. Res. 2019, 44, 13–26. [Google Scholar] [CrossRef]
- Talukder, M.Z.A.; Sarker, U.; Harun-Or-Rashid, M.; Zakaria, M. Genetic Diversity of Coconut (Cocos nucifera L.) in Barisal Region. Ann Bangladesh Agric 2015, 19, 13–21. [Google Scholar]
- Talukder, M.Z.A.; Sarker, U.; Khan ABM, M.M.; Moniruzzaman, M.; Zaman, M.M. Genetic Variability and Correlation Coefficient of Coconut (Cocos nucifera L.) in Barisal Region. Intl. J. BioRes. 2011, 11, 15–21. [Google Scholar]
- Jamshidmoghaddam, M.; Pourdad, S.S. Genotype× Environment Interactions for Seed Yield in Rainfed Winter Safflower (Carthamus tinctorius L.) Multi-Environment Trials in Iran. Euphytica 2013, 190, 357–369. [Google Scholar] [CrossRef]
- Islam, M.R.; Anisuzzaman, M.; Khatun, H.; Sharma, N.; Islam, M.Z.; Akter, A.; Biswas, P.S. AMMI Analysis of Yield Performance and Stability of Rice Genotypes across Different Haor Areas. Eco-Friendly Agril. J. 2014, 7, 20–24. [Google Scholar]
- Miranda, G.V.; De Souza, L.V.; Guimarães, L.J.M.; Namorato, H.; Oliveira, L.R.; Soares, M.O. Multivariate Analyses of Genotype x Environment Interaction of Popcorn. Pesqui. Agropecuária Bras. 2009, 44, 45–50. [Google Scholar] [CrossRef]
- Gauch, H.G.; Kang, M.S. Genotype-by-Environment Interaction/Genotype by Environment Interaction; CRC Press: Boca Raton, FL, USA, 1996. [Google Scholar]
- Da Silveira, L.C.I.; Kist, V.; de Paula, T.O.M.; Barbosa, M.H.P.; Peternelli, L.A.; Daros, E. AMMI Analysis to Evaluate the Adaptability and Phenotypic Stability of Sugarcane Genotypes. Sci. Agric. 2013, 70, 27–32. [Google Scholar] [CrossRef]
- Dehghani, H.; Ebadi, A.; Yousefi, A. Biplot Analysis of Genotype by Environment Interaction for Barley Yield in Iran. Agron. J. 2006, 98, 388–393. [Google Scholar] [CrossRef]
- Singamsetti, A.; Shahi, J.P.; Zaidi, P.H.; Seetharam, K.; Vinayan, M.T.; Kumar, M.; Singla, S.; Shikha, K.; Madankar, K. Genotype× Environment Interaction and Selection of Maize (Zea mays L.) Hybrids across Moisture Regimes. Field Crop. Res. 2021, 270, 108224. [Google Scholar] [CrossRef]
- Becker, H.C.; Leon, J. Stability Analysis in Plant Breeding. Plant Breed. 1988, 101, 1–23. [Google Scholar] [CrossRef]
- Ahmadi, J.; Vaezi, B.; Shaabani, A.; Khademi, K.; Fabriki Ourang, S.; Pour-Aboughadareh, A. Non-Parametric Measures for Yield Stability in Grass Pea (Lathyrus sativus L.) Advanced Lines in Semi-Warm Regions. J. Agric. Sci. Technol. 2015, 17, 1825–1838. [Google Scholar]
- Dalló, S.C.; Zdziarski, A.D.; Woyann, L.G.; Milioli, A.S.; Zanella, R.; Conte, J.; Benin, G. Across Year and Year-by-Year GGE Biplot Analysis to Evaluate Soybean Performance and Stability in Multi-Environment Trials. Euphytica 2019, 215, 113. [Google Scholar] [CrossRef]
- Bangladesh Agricultural Research Institute. Krishi Projukyi Hatboi (Handbook on Agro-Technology), 11th ed.; Farm Technology Group: Gazipur, Bangladesh, 2020. [Google Scholar]
- Sarker, U.; Hossain, M.N.; Oba, S.; Ercisli, S.; Marc, R.A.; Golokhvast, K.S. Salinity Stress Ameliorates Pigments, Minerals, Polyphenolic Profiles, and Antiradical Capacity in Lalshak. Antioxidants 2023, 12, 173. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S.; Ercisli, S.; Assouguem, A.; Alotaibi, A.; Ullah, R. Bioactive Phytochemicals and Quenching Activity of Radicals in Selected Drought-Resistant Amaranthus tricolor Vegetable Amaranth. Antioxidants 2022, 11, 578. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Rabbani, M.G.; Oba, S.; Eldehna, W.M.; Al-Rashood, S.T.; Mostafa, N.M.; Eldahshan, O.A. Phytonutrients, Colorant Pigments, Phytochemicals, and Antioxidant Potential of Orphan Leafy Amaranthus Species. Molecules 2022, 27, 2899. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S.; Alsanie, W.F.; Gaber, A. Characterization of Phytochemicals, Nutrients, and Antiradical Potential in Slim Amaranth. Antioxidants 2022, 11, 1089. [Google Scholar] [CrossRef]
- Sarker, U.; Iqbal, M.A.; Hossain, M.N.; Oba, S.; Ercisli, S.; Muresan, C.C.; Marc, R.A. Colorant Pigments, Nutrients, Bioactive Components, and Antiradical Potential of Danta Leaves (Amaranthus lividus). Antioxidants 2022, 11, 1206. [Google Scholar] [CrossRef]
- Sarker, U.; Ercisli, S. Salt Eustress Induction in Red Amaranth (Amaranthus gangeticus) Augments Nutritional, Phenolic Acids and Antiradical Potential of Leaves. Antioxidants 2022, 11, 2434. [Google Scholar] [CrossRef]
- Mamun, M.A.A.; Julekha; Sarker, U.; Mannan, M.A.; Rahman, M.M.; Karim, M.A.; Ercisli, S.; Marc, R.A.; Golokhvast, K.S. Application of Potassium after Waterlogging Improves Quality and Productivity of Soybean Seeds. Life 2022, 12, 1816. [Google Scholar] [CrossRef]
- Rahman, M.M.; Sarker, U.; Swapan, M.A.H.; Raihan, M.S.; Oba, S.; Alamri, S.; Siddiqui, M.H. Combining Ability Analysis and Marker-Based Prediction of Heterosis in Yield Reveal Prominent Heterotic Combinations from Diallel Population of Rice. Agronomy 2022, 12, 1797. [Google Scholar] [CrossRef]
- Prodhan, M.M.; Sarker, U.; Hoque, M.A.; Biswas, M.S.; Ercisli, S.; Assouguem, A. Foliar Application of GA3 Stimulates Seed Production in Cauliflower. Agronomy 2022, 12, 1394. [Google Scholar] [CrossRef]
- Fatema, M.K.; Mamun, M.A.A.; Sarker, U.; Hossain, M.S.; Mia, M.A.B.; Roychowdhury, R.; Ercisli, S.; Marc, R.A.; Babalola, O.O.; Karim, M.A. Assessing Morpho-Physiological and Biochemical Markers of Soybean for Drought Tolerance Potential. Sustainability 2023, 15, 1427. [Google Scholar] [CrossRef]
- Azad, A.K.; Sarker, U.; Ercisli, S.; Assouguem, A.; Ullah, R.; Almeer, R. Evaluation of Combining Ability and Heterosis of Popular Restorer and Male Sterile Lines for the Development of Superior Rice Hybrids. Agronomy 2022, 12, 965. [Google Scholar] [CrossRef]
- Hasan, M.J.; Kulsum, U.M.; Majumder, R.R.; Sarker, U. Genotypic variability for grain quality attributes in restorer lines of hybrid rice. Genetika 2020, 52, 973–989. [Google Scholar] [CrossRef]
- Hossain, M.N.; Sarker, U.; Raihan, M.S.; Al-Huqail, A.A.; Siddiqui, M.H.; Oba, S. Influence of Salinity Stress on Color Parameters, Leaf Pigmentation, Polyphenol and Flavonoid Contents, and Antioxidant Activity of Amaranthus lividus Leafy Vegetables. Molecules 2022, 27, 1821. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2009; Available online: http//www.r-project.org/ (accessed on 11 December 2022).
- Olivoto, T.; Lúcio, A.D. Metan: An R Package for Multi-environment Trial Analysis. Methods Ecol. Evol. 2020, 11, 783–789. [Google Scholar] [CrossRef]
- Pour-Aboughadareh, A.; Yousefian, M.; Moradkhani, H.; Poczai, P.; Siddique, K.H.M. STABILITYSOFT: A New Online Program to Calculate Parametric and Non-parametric Stability Statistics for Crop Traits. Appl. Plant Sci. 2019, 7, e01211. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-E Ects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entries | Code | Days to Flowering | Pi | bi | S2di | ||||
|---|---|---|---|---|---|---|---|---|---|
| Ish | Jas | Mym | Ran | Mean | |||||
| LRIL 18–102 | G1 | 63 | 55 | 57 | 60 | 59 | 29.7 | 1.50 | −3.18 |
| LRIL 21–139 | G2 | 61 | 56 | 56 | 61 | 59 | 29.8 | 1.15 | −1.67 |
| ILL 2580 | G3 | 66 | 62 | 60 | 65 | 63 | 8.96 | 0.92 | −1.05 |
| LRIL 22–165 | G4 | 62 | 56 | 57 | 59 | 58 | 29.4 | 1.17 | −3.18 |
| LRIL 21–198 | G5 | 60 | 51 | 53 | 53 | 54 | 69.3 | 1.50 | 0.76 |
| LRIL 22–158 | G6 | 61 | 55 | 53 | 58 | 57 | 46.8 | 1.34 | −1.08 |
| RL 12–171 | G7 | 64 | 63 | 58 | 65 | 62 | 14.2 | 0.77 | 5.39 * |
| LG 198 | G8 | 67 | 62 | 58 | 64 | 63 | 14.3 | 1.29 | 4.71 |
| LRIL 22–133 | G9 | 57 | 55 | 68 | 58 | 60 | 24.5 | −0.67 ** | 44.20 ** |
| BARI Masur-8 | G10 | 65 | 59 | 60 | 61 | 61 | 13 | 1.03 | −2.93 |
| Mean | 63 | 57 | 58 | 60 | 60 | ||||
| Environmental Index (Ij) | 3 | −3 | −2 | 0 | |||||
| CV (%) | 1.85 | 2.69 | 1.98 | 2.98 | 4.69 | ||||
| LSD (0.05) | 3.12 | 2.52 | 3.73 | 2.83 | 2.91 | ||||
| Entries | Code | Days to Maturity | Pi | bi | S2di | ||||
|---|---|---|---|---|---|---|---|---|---|
| Ish | Jas | Mym | Ran | Mean | |||||
| LRIL 18–102 | G1 | 93 | 86 | 98 | 88 | 91 | 53.3 | 1.05 | 28.40 * |
| LRIL 21–139 | G2 | 97 | 94 | 96 | 96 | 96 | 16.5 | 0.32 * | 0.14 |
| ILL 2580 | G3 | 100 | 91 | 96 | 90 | 94 | 23.8 | 1.70 * | 2.09 |
| LRIL 22–165 | G4 | 96 | 94 | 89 | 92 | 93 | 44.2 | 0.53 * | 8.17 * |
| LRIL 21–198 | G5 | 95 | 91 | 88 | 89 | 91 | 57.5 | 0.87 | 6.90 * |
| LRIL 22–158 | G6 | 98 | 94 | 91 | 94 | 94 | 28.4 | 0.68 | 5.20 * |
| RL 12–171 | G7 | 106 | 97 | 91 | 98 | 98 | 18.3 | 1.45 * | 29.80 * |
| LG 198 | G8 | 105 | 94 | 100 | 94 | 98 | 4.57 | 1.80 | 1.18 |
| LRIL 22–133 | G9 | 98 | 95 | 99 | 93 | 96 | 14.2 | 0.75 | 5.78 * |
| BARI Masur-8 | G10 | 102 | 98 | 103 | 96 | 100 | 1.61 | 0.86 | 6.10 * |
| Mean | 99 | 93 | 95 | 93 | 95 | ||||
| Environmental Index (Ij) | 5 | 2 | 0 | −2 | |||||
| CV (%) | 2.94 | 2.34 | 1.80 | 3.67 | 5.17 | ||||
| LSD (0.05) | 2.43 | 2.86 | 2.66 | 3.39 | 4.28 | ||||
| Entries | Code | Plant Height | Pi | bi | S2di | ||||
|---|---|---|---|---|---|---|---|---|---|
| Ish | Jas | Mym | Ran | Mean | |||||
| LRIL 18–102 | G1 | 38.0 | 37.0 | 40.0 | 39.0 | 38.5 | 2611 | 1.14 | −462.65 |
| LRIL 21–139 | G2 | 40.0 | 39.0 | 43.0 | 41.0 | 40.8 | −2489 | 1.20 | −472.34 |
| ILL 2580 | G3 | 42.0 | 41.0 | 42.0 | 43.0 | 42.0 | −2509 | 1.05 | −467.12 |
| LRIL 22–165 | G4 | 43.0 | 39.0 | 39.0 | 41.0 | 40.5 | 2647 | −1.18 | −470.36 |
| LRIL 21–198 | G5 | 35.0 | 32.0 | 34.0 | 31.0 | 33.0 | 2866 | 2.07 | −465.56 |
| LRIL 22–158 | G6 | 39.0 | 38.0 | 39.0 | 36.0 | 38.0 | −2629 | 1.08 | −476.50 |
| RL 12–171 | G7 | 42.0 | 44.0 | 44.0 | 45.0 | 43.8 | 2419 | 2.52 * | −466.47 |
| LG 198 | G8 | 44.0 | 41.0 | 40.0 | 42.0 | 41.8 | 2603 | −1.18 | −471.23 |
| LRIL 22–133 | G9 | 32.0 | 35.0 | 38.0 | 35.0 | 35.0 | 2711 | 1.27 | −458.45 |
| BARI Masur-8 | G10 | 35.0 | 40.0 | 38.0 | 38.0 | 37.8 | 2663 | 1.04 | −468.87 |
| Mean | 39.0 | 38.6 | 39.7 | 39.1 | 39.1 | ||||
| Environmental Index (Ij) | −0.2 | −0.5 | 0.6 | 0 | |||||
| CV (%) | 2.83 | 2.34 | 1.67 | 1.89 | 3.72 | ||||
| LSD (0.05) | 2.29 | 3.23 | 2.12 | 2.79 | 3.34 | ||||
| Entries | Code | Pods Per Plant | Pi | bi | S2di | ||||
|---|---|---|---|---|---|---|---|---|---|
| Ish | Jas | Mym | Ran | Mean | |||||
| LRIL 18–102 | G1 | 58 | 64 | 59 | 66 | 62 | 1049 | 0.38 | 10.60 |
| LRIL 21–139 | G2 | 74 | 68 | 64 | 70 | 69 | 727 | 0.75 | −0.16 |
| ILL 2580 | G3 | 122 | 106 | 87 | 107 | 106 | 113 | 2.63 | 55.20 * |
| LRIL 22–165 | G4 | 81 | 80 | 74 | 83 | 79 | 399 | 0.80 | −5.04 |
| LRIL 21–198 | G5 | 80 | 85 | 69 | 87 | 80 | 373 | 1.43 | 11.90 |
| LRIL 22–158 | G6 | 74 | 63 | 70 | 64 | 68 | 790 | −0.14 | 29.90 * |
| RL 12–171 | G7 | 60 | 59 | 53 | 62 | 58 | 1176 | 0.71 | −5.30 |
| LG 198 | G8 | 69 | 70 | 63 | 73 | 69 | 734 | 0.81 | −3.00 |
| LRIL 22–133 | G9 | 68 | 69 | 63 | 71 | 68 | 777 | 0.62 | −3.96 |
| BARI Masur-8 | G10 | 84 | 86 | 66 | 88 | 81 | 337 | 2.01 * | 3.39 |
| Mean | 77 | 75 | 67 | 77 | 74 | ||||
| Environmental Index (Ij) | 3 | 1 | −7 | 3 | |||||
| CV (%) | 2.35 | 3.16 | 2.51 | 3.14 | 4.23 | ||||
| LSD (0.05) | 4.98 | 2.59 | 3.34 | 5.94 | 3.61 | ||||
| Entries | Code | 100 Seed Weight | Pi | bi | S2di | ||||
|---|---|---|---|---|---|---|---|---|---|
| Ish | Jas | Mym | Ran | Mean | |||||
| LRIL 18–102 | G1 | 1.66 | 1.65 | 1.37 | 1.72 | 1.60 | 0.48 | 1.20 | 0.35 |
| LRIL 21–139 | G2 | 1.89 | 1.93 | 1.75 | 2.00 | 1.89 | 0.23 | 0.75 | 0.24 |
| ILL 2580 | G3 | 1.23 | 1.16 | 0.96 | 1.25 | 1.15 | 1.02 | 1.00 * | 0.99 |
| LRIL 22–165 | G4 | 1.86 | 1.78 | 1.64 | 1.87 | 1.79 | 0.31 | 0.81 | 0.38 |
| LRIL 21–198 | G5 | 2.19 | 2.04 | 1.92 | 2.13 | 2.07 | 0.13 | 0.82 | 0.40 |
| LRIL 22–158 | G6 | 2.03 | 2.10 | 1.87 | 2.19 | 2.05 | 0.14 | 0.95 | 0.81 |
| RL 12–171 | G7 | 1.69 | 1.57 | 1.36 | 1.67 | 1.57 | 0.51 | 1.13 | 0.55 |
| LG 198 | G8 | 1.75 | 1.71 | 1.50 | 1.81 | 1.69 | 0.39 | 1.04 | 0.84 |
| LRIL 22–133 | G9 | 2.43 | 2.64 | 2.41 | 2.74 | 2.55 | 0.04 | 0.78 | 0.32 * |
| BARI Masur-8 | G10 | 2.52 | 2.17 | 1.94 | 2.27 | 2.22 | 0.08 | 2.41 * | 0.03 * |
| Mean | 1.92 | 1.87 | 1.67 | 1.96 | 1.86 | ||||
| Environmental Index (Ij) | 0.06 | 0.01 | −0.19 | 0.10 | |||||
| CV (%) | 1.15 | 1.12 | 1.21 | 1.34 | 1.29 | ||||
| LSD (0.05) | 0.68 | 0.59 | 0.34 | 0.74 | 0.51 | ||||
| Entries | Seed Yield | Pi | bi | S2di | |||||
|---|---|---|---|---|---|---|---|---|---|
| Ish | Jas | Mym | Ran | Mean | |||||
| LRIL 18–102 | G1 | 1164 | 970 | 786 | 974 | 973 | 144,789 | 1.38 | 1885.0 * |
| LRIL 21–139 | G2 | 1320 | 1225 | 1108 | 1239 | 1223 | 41,953 | 0.76 | 1036.0* |
| ILL 2580 | G3 | 1275 | 1253 | 858 | 1251 | 1159 | 71,198 | 1.51 | 169.0 |
| LRIL 22–165 | G4 | 1550 | 1329 | 1238 | 1227 | 1336 | 15,345 | 1.24 | 6286.0 * |
| LRIL 21–198 | G5 | 1335 | 1259 | 1231 | 1255 | 1270 | 29,767 | 0.39 | 233.3 * |
| LRIL 22–158 | G6 | 1460 | 1269 | 1220 | 1218 | 1292 | 23,352 | 0.93 | 4202.0 * |
| RL 12–171 | G7 | 1658 | 1515 | 1425 | 1430 | 1507 | 5356 | 0.93 | 2291.0 * |
| LG 198 | G8 | 1065 | 999 | 861 | 942 | 967 | 146,946 | 0.80 | −346.3 |
| LRIL 22–133 | G9 | 1255 | 1270 | 1044 | 1148 | 1179 | 55,907 | 0.87 | 3049.0 * |
| BARI Masur-8 | G10 | 1550 | 1515 | 1266 | 1314 | 1411 | 6304 | 1.21 | 4245.0 * |
| Mean | 1363 | 1260 | 1104 | 1200 | 1232 | ||||
| Environmental Index (Ij) | 131 | 28 | −128 | −32 | |||||
| CV (%) | 5.45 | 7.52 | 6.71 | 8.84 | 6.89 | ||||
| LSD (0.05) | 45.56 | 48.66 | 52.43 | 49.87 | 38.98 | ||||
| Source | df | DF | DM | PH (cm) | PPP | Yield (kg/ ha) | Cumulative % | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MS | %Total SS | MS | %Total SS | MS | %Total SS | MS | %Total SS | MS | %Total SS | |||
| Environments | 2 | 256.07 ** | 30.69 | 237.21 | 24.58 ** | 2207.77 ns | 7.14 | 948.68 ** | 10.50 | 491,049.30 ** | 24.06 | 24.06 |
| Genotypes | 9 | 64.67 ** | 34.88 | 92.96 | 43.34 ** | 2428.43 ns | 35.33 | 1586.49 ** | 78.98 | 319,398.80 ** | 70.41 | 94.47 |
| G × E Interaction | 18 | 31.92 ** | 34.44 | 34.41 | 32.08 ** | 1976.93 ns | 57.53 | 105.71 ** | 10.53 | 12,555.00 ** | 5.53 | 100 |
| IPCA1 | 10 | 49.79 ** | 86.63 | 49.25 | 79.52 ** | 3550.62 ns | 99.78 | 123.29 ** | 64.79 | 15,370.29 ** | 68.01 | 68.01 |
| IPCA2 | 8 | 9.60 ns | 13.37 | 15.86 | 20.48 ** | 9.81 ns | 0.22 | 83.74 ** | 35.21 | 9035.90 ** | 31.99 | 100 |
| Residuals | 60 | 13.87778 | - | 2.96 | - | 1880.51 | - | 27.61 | - | 2152.37 | - | |
| Genotype | Mean Yield | S(1) | S(2) | S⁽³⁾ | S⁽⁶⁾ | N(1) | N(2) | N(3) | N(4) |
|---|---|---|---|---|---|---|---|---|---|
| G1 | 973 | 4.17 | 10.92 | 0.50 | 1.00 | 2.75 | 0.29 | 0.30 | 0.11 |
| G2 | 1223 | 3.00 | 6.33 | 1.00 | 0.71 | 1.50 | 0.25 | 0.35 | 0.11 |
| G3 | 1159 | 5.17 | 16.92 | 2.92 | 1.23 | 3.25 | 0.46 | 0.53 | 0.22 |
| G4 | 1336 | 5.17 | 16.25 | 0.86 | 0.57 | 3.25 | 1.08 | 0.93 | 0.27 |
| G5 | 1270 | 5.33 | 18.00 | 0.70 | 0.50 | 3.50 | 0.78 | 0.82 | 0.11 |
| G6 | 1292 | 4.67 | 13.67 | 0.50 | 0.50 | 3.00 | 0.60 | 0.61 | 0.16 |
| G7 | 1507 | 2.67 | 4.67 | 0.07 | 0.14 | 1.50 | 1.50 | 1.50 | 0.67 |
| G8 | 967 | 1.50 | 1.58 | 1.00 | 1.00 | 0.75 | 0.08 | 0.12 | 0.00 |
| G9 | 1179 | 3.17 | 6.25 | 1.86 | 1.00 | 1.75 | 0.23 | 0.32 | 0.02 |
| G10 | 1411 | 4.50 | 12.92 | 0.07 | 0.14 | 2.75 | 1.38 | 1.78 | 0.10 |
| Rank | |||||||||
| Genotype | Mean Yield | S(1) | S(2) | S⁽³⁾ | S⁽⁶⁾ | N(1) | N(2) | N(3) | N(4) |
| G1 | 9 | 5 | 5 | 3 | 7 | 5 | 4 | 2 | 4 |
| G2 | 6 | 3 | 4 | 7 | 6 | 2 | 3 | 4 | 5 |
| G3 | 8 | 8 | 9 | 10 | 10 | 8 | 5 | 5 | 8 |
| G4 | 3 | 9 | 8 | 6 | 5 | 9 | 8 | 8 | 9 |
| G5 | 5 | 10 | 10 | 5 | 3 | 10 | 7 | 7 | 6 |
| G6 | 4 | 7 | 7 | 4 | 4 | 7 | 6 | 6 | 7 |
| G7 | 1 | 2 | 2 | 1 | 1 | 3 | 10 | 9 | 10 |
| G8 | 10 | 1 | 1 | 8 | 7 | 1 | 1 | 1 | 1 |
| G9 | 7 | 4 | 3 | 9 | 7 | 4 | 2 | 3 | 2 |
| G10 | 2 | 6 | 6 | 2 | 2 | 6 | 9 | 10 | 3 |
| Genotype | Mean Yield | ASV | AMGE | ASI | ASTAB | AVAMGE | DA | DZ | FA | MASI | MASV | SIPC | Za |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| G1 | 973 | 8.16 | 9.95 | 1.90 | 53.73 | 182.30 | 95.01 | 0.58 | 9027.49 | 2.05 | 10.95 | 12.26 | 0.24 |
| G2 | 1223 | 3.07 | 1.78 | 1.03 | 26.95 | 121.37 | 65.65 | 0.41 | 4310.49 | 1.14 | 7.10 | 7.94 | 0.14 |
| G3 | 1159 | 4.41 | 3.20 | 8.18 | 180.66 | 377.03 | 222.66 | 0.81 | 49,578.69 | 8.18 | 35.17 | 15.73 | 0.53 |
| G4 | 1336 | 35.10 | 6.04 | 3.58 | 69.62 | 224.30 | 121.78 | 0.59 | 14,830.81 | 3.58 | 17.01 | 12.29 | 0.32 |
| G5 | 1270 | 15.40 | −2.84 | 2.88 | 75.30 | 212.03 | 119.05 | 0.64 | 14,172.99 | 2.90 | 15.21 | 13.18 | 0.30 |
| G6 | 1292 | 12.40 | 3.55 | 3.25 | 35.42 | 185.30 | 93.94 | 0.39 | 8824.79 | 3.27 | 14.27 | 9.13 | 0.25 |
| G7 | 1507 | 14.00 | 3.55 | 2.57 | 18.53 | 131.97 | 70.70 | 0.26 | 4998.25 | 2.57 | 11.09 | 5.50 | 0.18 |
| G8 | 967 | 11.00 | −3.42 | 0.74 | 9.60 | 65.88 | 40.47 | 0.24 | 1637.61 | 0.75 | 4.70 | 4.09 | 0.08 |
| G9 | 1179 | 3.18 | −8.53 | 1.50 | 43.37 | 125.37 | 83.40 | 0.53 | 6955.27 | 1.71 | 9.12 | 10.68 | 0.21 |
| G10 | 1411 | 6.43 | −6.75 | 0.72 | 72.19 | 165.63 | 101.50 | 0.71 | 10,302.25 | 1.42 | 9.18 | 11.13 | 0.16 |
| Rank | |||||||||||||
| Genotype | ryield | rASV | rAMGE | rASI | rASTAB | rAVAMGE | rDA | rDZ | rFA | rMASI | rMASV | rSIPC | rZa |
| G1 | 9 | 5 | 10 | 5 | 6 | 6 | 6 | 6 | 6 | 5 | 5 | 7 | 6 |
| G2 | 6 | 1 | 5 | 3 | 3 | 2 | 2 | 4 | 2 | 2 | 2 | 3 | 2 |
| G3 | 8 | 3 | 7 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 |
| G4 | 3 | 10 | 9 | 9 | 7 | 9 | 9 | 7 | 9 | 9 | 9 | 8 | 9 |
| G5 | 5 | 9 | 4 | 7 | 9 | 8 | 8 | 8 | 8 | 7 | 8 | 9 | 8 |
| G6 | 4 | 7 | 8 | 8 | 4 | 7 | 5 | 3 | 5 | 8 | 7 | 4 | 7 |
| G7 | 1 | 8 | 6 | 6 | 2 | 4 | 3 | 2 | 3 | 6 | 6 | 2 | 4 |
| G8 | 10 | 6 | 3 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| G9 | 7 | 2 | 1 | 4 | 5 | 3 | 4 | 5 | 4 | 4 | 3 | 5 | 5 |
| G10 | 2 | 4 | 2 | 1 | 8 | 5 | 7 | 9 | 7 | 3 | 4 | 6 | 3 |
| Genotype | Mean Yield | CV | bi | S2di | Tai | Wi2 | Pi | R2 | σ2 |
|---|---|---|---|---|---|---|---|---|---|
| G1 | 973 | 14.37 | 1.38 | 2004.06 | 0.38 | 9027.49 | 144,789 | 0.94 | 3184 |
| G2 | 1223 | 6.62 | 0.76 | 1155.24 | 0.21 | 4310.49 | 41,953 | 0.90 | 1219 |
| G3 | 1159 | 15.71 | 1.51 | 20,252.21 | −0.24 | 49,578.69 | 71,198 | 0.67 | 20081 |
| G4 | 1336 | 10.25 | 1.24 | 6405.73 | 0.51 | 14,830.81 | 15,345 | 0.81 | 5602 |
| G5 | 1270 | 3.40 | 0.39 | 395.07 | 0.24 | 14,172.99 | 29,767 | 0.87 | 5328 |
| G6 | 1292 | 8.12 | 0.93 | 4321.67 | −0.61 | 8824.79 | 23,352 | 0.78 | 3100 |
| G7 | 1507 | 6.61 | 0.93 | 2410.11 | −0.07 | 4998.25 | 5536. | 0.86 | 1506 |
| G8 | 967 | 8.35 | 0.80 | 73.23 | −0.07 | 1637.61 | 146,946 | 0.99 | 105 |
| G9 | 1179 | 8.22 | 0.87 | 3168.33 | −0.21 | 6955.27 | 55,907 | 0.81 | 2321 |
| G10 | 1411 | 9.15 | 1.21 | 4364.56 | −0.13 | 10,302.25 | 6304 | 0.86 | 3716 |
| Genotype | r yield | CV | bi | S2di | Tai | W2 | Pi | R2 | σ2i |
| G1 | 9 | 9 | 2 | 4 | 5 | 6 | 9 | 2 | 6 |
| G2 | 6 | 3 | 9 | 3 | 2 | 2 | 6 | 3 | 2 |
| G3 | 8 | 10 | 1 | 10 | 8 | 10 | 8 | 10 | 10 |
| G4 | 3 | 8 | 3 | 9 | 9 | 9 | 3 | 7 | 9 |
| G5 | 5 | 1 | 10 | 2 | 6 | 8 | 5 | 4 | 8 |
| G6 | 4 | 4 | 6 | 7 | 7 | 5 | 4 | 9 | 5 |
| G7 | 1 | 2 | 5 | 5 | 10 | 3 | 1 | 5 | 3 |
| G8 | 10 | 6 | 8 | 1 | 4 | 1 | 10 | 1 | 1 |
| G9 | 7 | 5 | 7 | 6 | 1 | 4 | 7 | 8 | 4 |
| G10 | 2 | 7 | 4 | 8 | 3 | 7 | 2 | 6 | 7 |
| Gen. | ASV | AMGE | ASI | ASTAB | AVAMGE | DA | DZ | FA | MASI | MASV | SIPC | Za |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| G1 | 14 | 19 | 14 | 15 | 15 | 15 | 15 | 15 | 14 | 14 | 16 | 15 |
| G2 | 9 | 11 | 9 | 9 | 8 | 8 | 10 | 8 | 8 | 8 | 9 | 8 |
| G3 | 18 | 15 | 18 | 18 | 18 | 18 | 18 | 18 | 18 | 18 | 18 | 18 |
| G4 | 12 | 12 | 12 | 10 | 12 | 12 | 10 | 12 | 12 | 12 | 11 | 12 |
| G5 | 12 | 9 | 12 | 14 | 13 | 13 | 13 | 13 | 12 | 13 | 14 | 13 |
| G6 | 12 | 12 | 12 | 8 | 11 | 9 | 7 | 9 | 12 | 11 | 8 | 11 |
| G7 | 7 | 7 | 7 | 3 | 5 | 4 | 3 | 4 | 7 | 7 | 3 | 5 |
| G8 | 12 | 13 | 12 | 11 | 11 | 11 | 11 | 11 | 11 | 11 | 11 | 11 |
| G9 | 11 | 8 | 11 | 12 | 10 | 11 | 12 | 11 | 11 | 10 | 12 | 12 |
| G10 | 3 | 4 | 3 | 10 | 7 | 9 | 11 | 9 | 5 | 6 | 8 | 5 |
| Genotypes | Mean Yield | HMRPGV | HMGV | RPGV | ||||
|---|---|---|---|---|---|---|---|---|
| Kg/ha | Rank | Score | Rank | Score | Rank | Score | Rank | |
| G1 | 973 | 9 | 0.78 | 10 | 955.00 | 10 | 0.79 | 9 |
| G2 | 1223 | 6 | 0.99 | 6 | 1218.00 | 6 | 0.99 | 6 |
| G3 | 1159 | 8 | 0.93 | 8 | 1129.00 | 8 | 0.94 | 8 |
| G4 | 1336 | 3 | 1.08 | 3 | 1324.00 | 3 | 1.08 | 3 |
| G5 | 1270 | 5 | 1.03 | 5 | 1269.00 | 5 | 1.03 | 5 |
| G6 | 1292 | 4 | 1.05 | 4 | 1285.00 | 4 | 1.05 | 4 |
| G7 | 1507 | 1 | 1.22 | 1 | 1501.00 | 1 | 1.23 | 1 |
| G8 | 967 | 10 | 0.79 | 9 | 961.00 | 9 | 0.79 | 10 |
| G9 | 1179 | 7 | 0.96 | 7 | 1172.00 | 7 | 0.96 | 7 |
| G10 | 1411 | 2 | 1.14 | 2 | 1401.00 | 2 | 1.15 | 2 |
| Sl. No. | Genotype Name | Genotype Code | Sources |
|---|---|---|---|
| 1 | LRIL 18–102 | G1 | ICARDA |
| 2 | LRIL 21–139 | G2 | ICARDA |
| 3 | ILL 2580 | G3 | ICARDA |
| 4 | LRIL 22–165 | G4 | ICARDA |
| 5 | LRIL 21–198 | G5 | ICARDA |
| 6 | LRIL 22–158 | G6 | ICARDA |
| 7 | RL 12–171 | G7 | ICARDA |
| 8 | LG 198 | G8 | ICARDA |
| 9 | LRIL 22–133 | G9 | ICARDA |
| 10 | BARI Masur-8 (Check variety) | G10 | PRC, BARI |
| Locations | Locations Code | Geographical Position | Altitude (m.a.s.l.) | Temperature (°C) | Relative Humidity (%) | Rainfall (mm) | Soil Type | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Latitude | Longitude | Max | Min | Max | Min | |||||
| PRC, Ishurdi | ISD | 24.1292° N | 89.0657° E | 18 | 30.5 | 11.75 | 94.5 | 64.2 | 27.3 | Clay loam |
| RARS, Jashore | JAS | 23.1778° N | 89.1801° E | 11 | 27.5 | 11.50 | 92.4 | 68.6 | 35.6 | Loamy to clay loam |
| BAU, Mymenshingh | MYM | 24.7851° N | 90.3560° E | 19 | 30.2 | 13.50 | 93.6 | 63.6 | 25.4 | Clay loam |
| OFRD, Rangpur | RAN | 25.7466° N | 89.2516° E | 34 | 28.2 | 11.30 | 91.5 | 60.6 | 20.4 | Sandy loamy to clay loam |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, M.A.; Sarker, U.; Azam, M.G.; Kobir, M.S.; Roychowdhury, R.; Ercisli, S.; Ali, D.; Oba, S.; Golokhvast, K.S. Integrating BLUP, AMMI, and GGE Models to Explore GE Interactions for Adaptability and Stability of Winter Lentils (Lens culinaris Medik.). Plants 2023, 12, 2079. https://doi.org/10.3390/plants12112079
Hossain MA, Sarker U, Azam MG, Kobir MS, Roychowdhury R, Ercisli S, Ali D, Oba S, Golokhvast KS. Integrating BLUP, AMMI, and GGE Models to Explore GE Interactions for Adaptability and Stability of Winter Lentils (Lens culinaris Medik.). Plants. 2023; 12(11):2079. https://doi.org/10.3390/plants12112079
Chicago/Turabian StyleHossain, Md. Amir, Umakanta Sarker, Md. Golam Azam, Md. Shahriar Kobir, Rajib Roychowdhury, Sezai Ercisli, Daoud Ali, Shinya Oba, and Kirill S. Golokhvast. 2023. "Integrating BLUP, AMMI, and GGE Models to Explore GE Interactions for Adaptability and Stability of Winter Lentils (Lens culinaris Medik.)" Plants 12, no. 11: 2079. https://doi.org/10.3390/plants12112079
APA StyleHossain, M. A., Sarker, U., Azam, M. G., Kobir, M. S., Roychowdhury, R., Ercisli, S., Ali, D., Oba, S., & Golokhvast, K. S. (2023). Integrating BLUP, AMMI, and GGE Models to Explore GE Interactions for Adaptability and Stability of Winter Lentils (Lens culinaris Medik.). Plants, 12(11), 2079. https://doi.org/10.3390/plants12112079