
Identification of Green-Leaf Volatiles Released from Cabbage Palms (Sabal palmetto) Infected with the Lethal Bronzing Phytoplasma

 ,
, 
 , , and
, , and 
Abstract
:1. Introduction
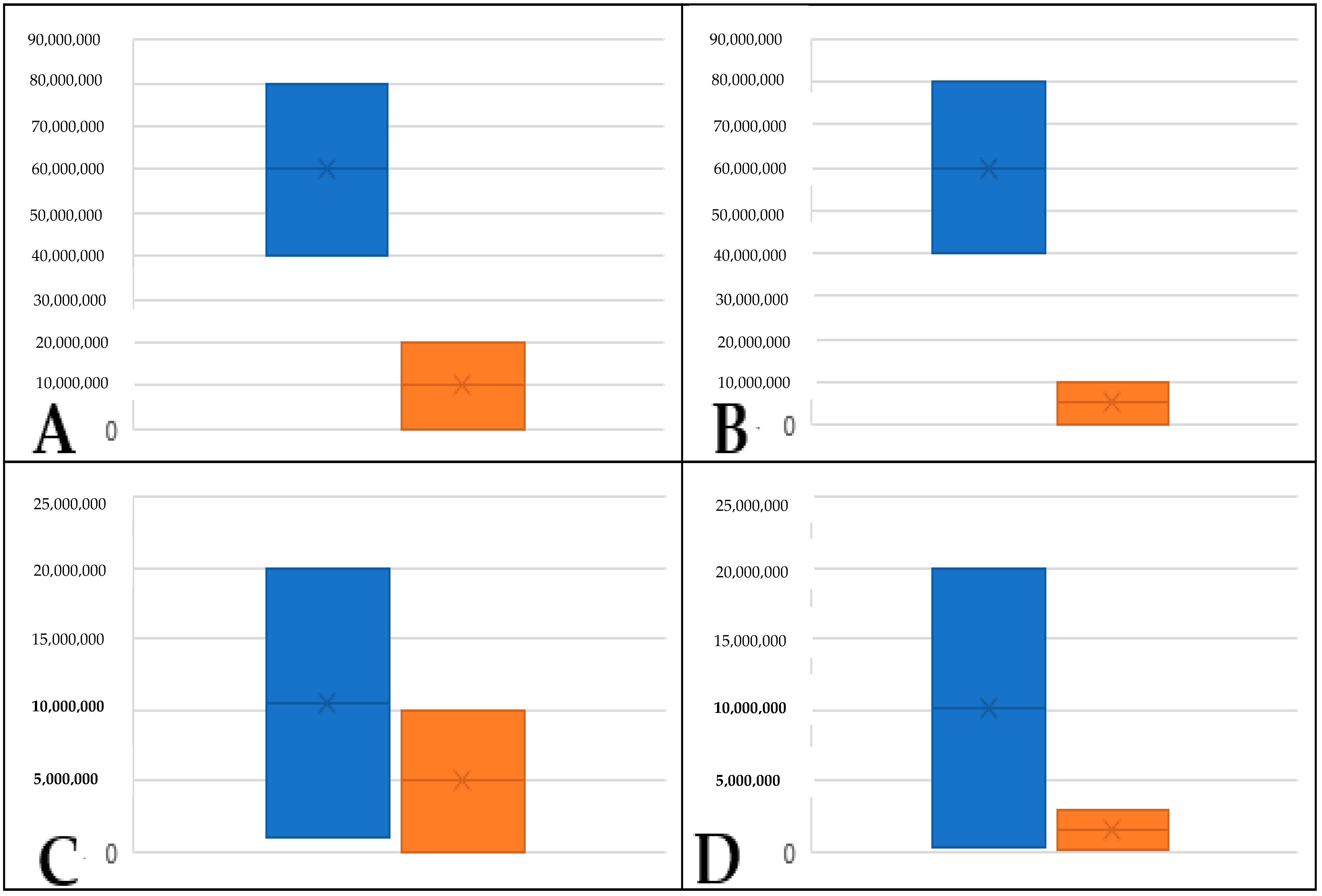
2. Results
2.1. qPCR Data and Confirmation of Infection Status
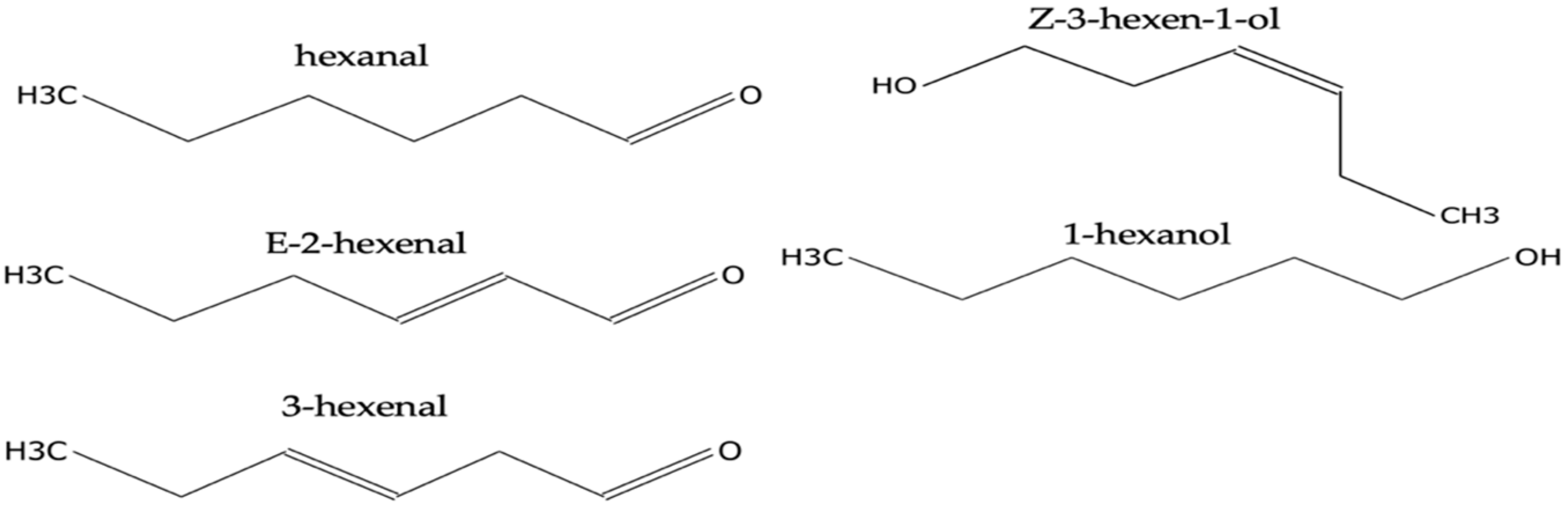
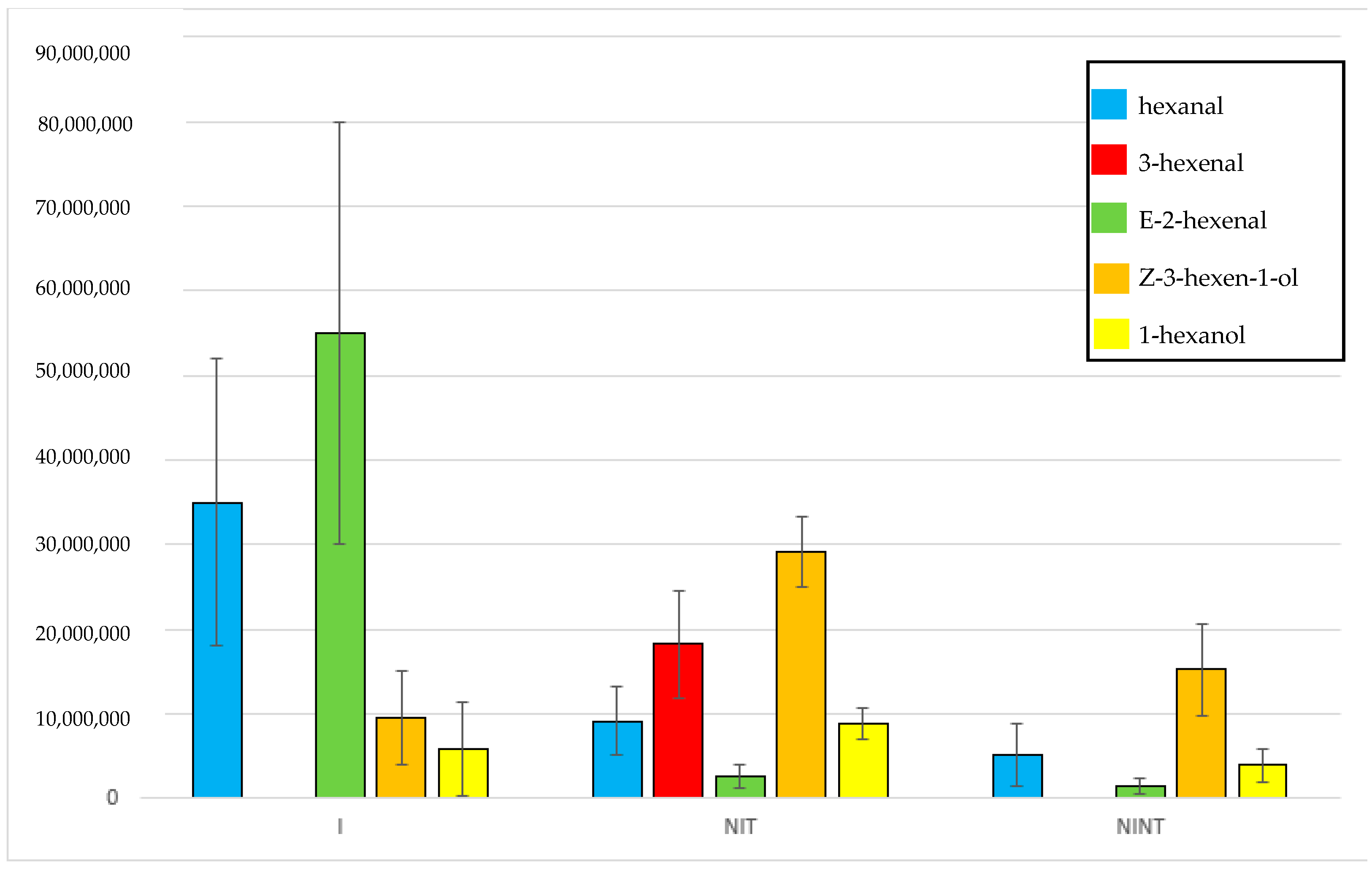
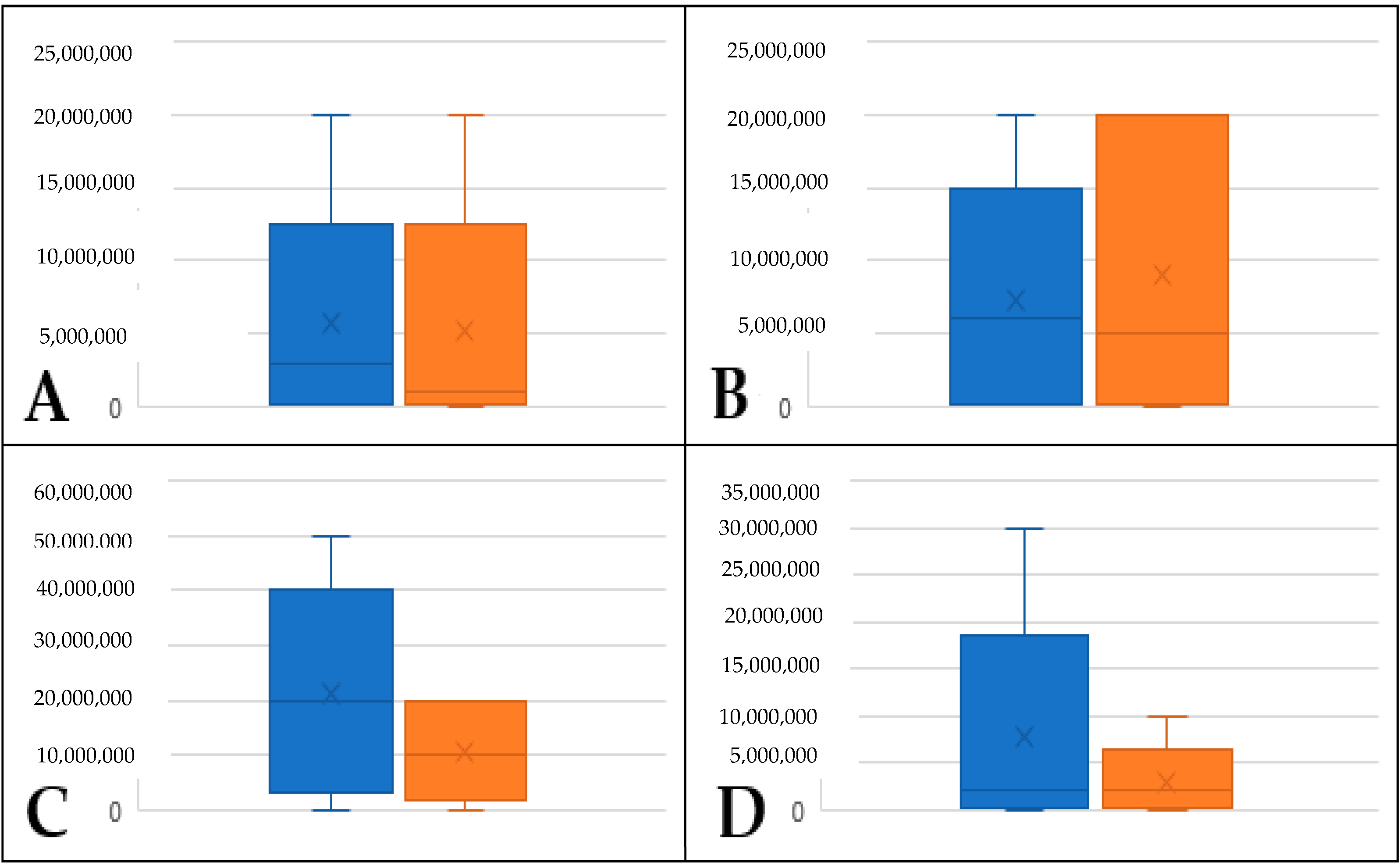
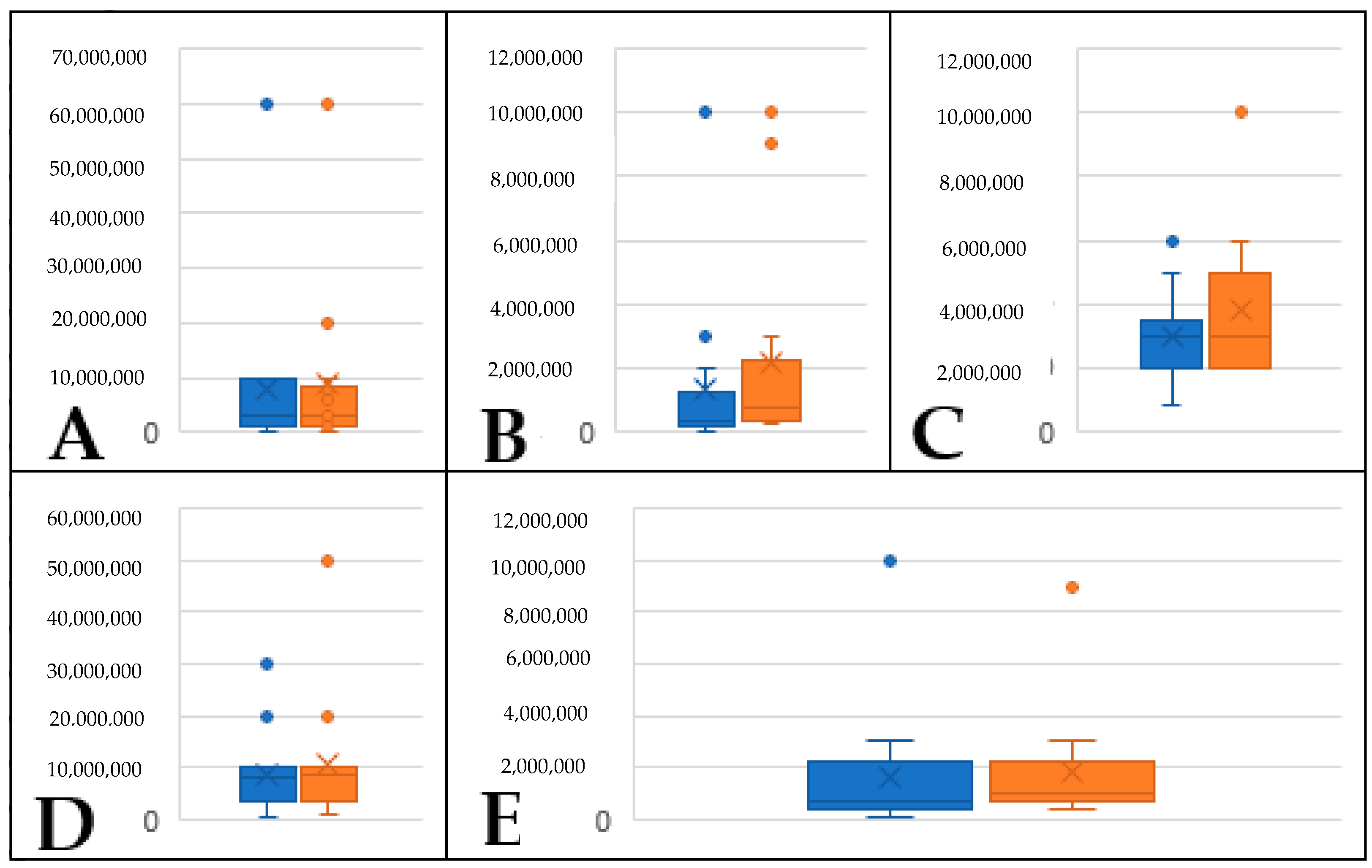
2.2. Identification of Volatiles Released from S. palmetto Leaves
2.3. Evaluation and Confirmation of GLVs Found in S. palmetto Leaf Samples
3. Discussion
4. Materials and Methods
4.1. Sample Selection and Verification of Infection Status
4.2. Identification of Volatile Profiles via HS-SPME/GC-MS
4.2.1. Reagents and Materials
4.2.2. Sample Preparation
4.2.3. HS-SPME Procedure
4.2.4. GC-MS Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mou, D.F.; Helmick, E.E.; Bahder, B.W. Multi-locus sequence analysis reveals new hosts of palm lethal decline phytoplasmas in Florida, U.S.A. Plant Health Prog. 2022, 23, 399–408. [Google Scholar] [CrossRef]
- Bahder, B.W.; Soto, N.; Helmick, E.E.; Dey, K.K.; Komondy, L.; Humphries, A.R.; Mou, D.F.; Bailey, R.; Ascunce, M.; Goss, E.M. A survey of declining palms (Arecaceae) with 16SrIV-D phytoplasma to evaluate the distribution and host range in Florida. Plant Dis. 2019, 103, 2512–2519. [Google Scholar] [CrossRef]
- Mou, D.F.; Di Lella, B.; Halbert, S.E.; Bextine, B.; Helmick, E.E.; Bahder, B.W. Acquisition and transmission of the lethal bronzing phytoplasma by Haplaxius crudus using infected palm spear leaves and artificial feeding media. Phytopathology 2022, 112, 2052–2061. [Google Scholar] [CrossRef]
- Humphries, A.R.; Ascunce, M.S.; Goss, E.M.; Helmick, E.E.; Bartlett, C.R.; Myrie, W.; Barrantes, E.A.B.; Zumbado, M.A.; Bustillo, A.E.; Bahder, B.W. Genetic variability of Haplaxius crudus, based on the 5′ region of the cyotchome c oxidase subunit I gene, sheds light on epidemiology of palm lethal decline phytoplasmas. Phytofrontiers 2021, 1, 127–134. [Google Scholar] [CrossRef]
- Mou, D.F.; Lee, C.C.; Hahn, P.G.; Soto, N.; Humphries, A.R.; Helmick, E.E.; Bahder, B.W. Effects of lethal bronzing disease, palm height and temperature on abundance and monitoring of Haplaxius crudus. Insects 2020, 11, 748. [Google Scholar] [CrossRef] [PubMed]
- Oates, M.J.; Abu-Khalaf, N.; Molina-Cabrera, C.; Ruiz-Canales, A.; Ramos, J.; Bahder, B.W. Detection of lethal bronzing disease in cabbage palms (Sabal palmetto) using a low-cost electronic nose. Biosensors 2020, 10, 188. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K. Green leaf volatiles: Hydroperoxide lyase pathway of oxylipin metabolism. Curr. Opin. Plant Biol. 2006, 9, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Ameye, M.; Allmann, S.; Verwaeren, J.; Smagghe, G.; Haesaert, G.; Schuurink, R.C.; Audenaert, K. Green leaf volatile production by plants: A meta-Analysis. New Phytol. 2018, 220, 666–668. [Google Scholar] [CrossRef]
- Shiojiri, K.; Kishimoto, K.; Ozawa, R.; Kugimiya, S.; Urashimo, S.; Arimura, G.; Horiuchi, J.; Nishioka, T.; Matsui, K.; Takabayashi, J. Changing green leaf volatile biosynthesis in plants: An approach for improving plant resistance against both herbivores and pathogens. Proc. Natl. Acad. Sci. USA 2006, 103, 16672–16676. [Google Scholar] [CrossRef]
- Chehab, E.W.; Kaspi, R.; Savchenko, T.; Rowe, H.; Negre-Zakharov, F.; Kliebenstein, D.; Dehesh, K. Distinct roles of jasmonates and aldehydes in plant-defenseresponses. PLoS ONE 2008, 3, e1904. [Google Scholar] [CrossRef]
- Schwartzberg, E.G.; Böröczky, K.; Tumlinson, J.H. Pea aphids, Acyrthosiphonpisum, suppress induced plant volatiles in broad bean, Vicia faba. J. Chem. Ecol. 2011, 37, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Kant, M.R.; Jonckheere, W.; Knegt, B.; Lemos, F.; Liu, J.; Schimmel, B.C.J.; Villarroel, C.A.; Ataide, L.M.S.; Dermauw, W.; Glas, J.J.; et al. Mechanisms and ecological consequences of plant defence induction and suppression in herbivore communities. Ann. Bot. 2015, 115, 1015–1051. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Huang, J.; Akutse, K.; Hou, Y. Phytopathogens increase the preference of insect vectors to volatiles emitted by healthy host plants. J. Agric. Food Chem. 2022, 70, 5262–5269. [Google Scholar] [CrossRef] [PubMed]
- Rid, M.; Mesca, C.; Ayasse, M.; Gross, J. Apple Proliferation Phytoplasma Influences the Pattern of Plant Volatiles Emitted Depending on Pathogen Virulence. Front. Ecol. Evol. 2016, 3, 152. [Google Scholar] [CrossRef]
- De Vleesschauwer, D.; Xu, J.; Höfte, M. Making sense of hormone-mediated defense networking: From rice to Arabidopsis. Front. Plant Sci. 2014, 5, 611. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.; Rieu, I.; Mariani, C.; van Dam, N.M. How plants handle multiplestresses: Hormonal interactions underlying responses to abiotic stress and insect herbivory. Plant Mol. Biol. 2016, 91, 727–740. [Google Scholar] [CrossRef]
- Halitschke, R.; Ziegler, J.; Keinänen, M.; Baldwin, I.T. Silencing of hydroperoxide lyase and allene oxide synthase reveals substrate and defense signaling crosstalk in Nicotiana attenuata. Plant J. 2004, 40, 35–46. [Google Scholar] [CrossRef]
- Allmann, S.; Baldwin, I.T. Insects betray themselves in nature to predators by rapid isomerization of green leaf volatiles. Science 2010, 329, 1075–1078. [Google Scholar] [CrossRef]
- Nakamura, S.; Hatanaka, A. Green-leaf-derived C6-aroma compounds with potent antibacterial action that act on both gram-negative and gram-positive bacteria. J. Agric. Food Chem. 2002, 50, 7639–7644. [Google Scholar] [CrossRef]
- Burzynski-Chang, E.A.; Ryona, I.; Reisch, B.I.; Gonda, I.; Foolad, M.R.; Giovannoni, J.J.; Sacks, G.L. HS-SPME-GC-MS Analyses of Volatiles in Plant Populations—Quantitating Compound × Individual Matrix Effects. Molecules 2018, 23, 2436. [Google Scholar] [CrossRef]
- Silva, E.A.S.; Saboia, G.; Jorge, N.C.; Hoffmann, C.; Isaias, R.M.S.; Soares, G.L.G.; Zini, C.A. Development of a HS-SPME-GC/MS protocol assisted by chemometric tools to study herbivore-induced volatiles in Myrcia splendens. Talanta 2017, 175, 9–20. [Google Scholar] [CrossRef]
- Lobo, A.P.; da Camara, C.A.G.; de Melo, J.P.R.; de Moraes, M.M. Chemical composition and repellent activity of essential oils from the leaves of Cinnamomum zylanicum and Eugenia uniflora against Diaphania hyalinata L. (Lepidoptera: Crambidae). J. Plant Dis. Prot. 2019, 126, 79–87. [Google Scholar] [CrossRef]
- Scarpati, M.L.; Scalzo, R.L.; Vita, G. Olea europaea volatiles attractive and repellant to the olive fruit fly (Dacus oleae, Gmelin). J. Chem. Ecol. 1993, 19, 881–891. [Google Scholar] [CrossRef] [PubMed]
- Giannoukos, S.; Agapiou, A.; Brkić, B.; Taylor, S. Volatolomics: A broad area of experimentation. J. Chromatogr. B 2019, 1105, 136–147. [Google Scholar] [CrossRef]
- Bahder, B.W.; Soto, N.; Mou, D.F.; Humphries, A.R.; Helmick, E.E. Quantification and distribution of the 16SrIV-D phytoplasma in the wild date palm, Phoenix sylvestris, at different stages of decline using quantitative PCR (qPCR) analysis. Plant Dis. 2020, 104, 1328–1334. [Google Scholar] [CrossRef] [PubMed]
- Velickovic, M.M.; Radivojevic, D.D.; Oparnica, C.D.; Nikicevic, N.J.; Zivkovic, M.B.; Dordevic, N.O.; Vajs, V.E.; Tesevic, V.V. Volatile compoundsin Medlar fruit (Mespilus germanica L.) at two ripening stages. Hemijskaindustrija 2013, 67, 437–441. [Google Scholar]
- Zhang, Y.; Kong, J.; Huang, F.; Xie, Y.; Guo, Y.; Cheng, Y.; Qian, H.; Yao, W. Hexanal as a QS inhibitor of extracellular enzyme activity of Erwinia carotovora and Pseudomonas fluorescens and its application in vegetables. Food Chem. 2018, 255, 1–7. [Google Scholar] [CrossRef]
- Dhakshinamoorthy, D.; Sundaresan, S.; Iyadurai, A.; Subramanian, K.S.; Janavi, G.J.; Paliyath, G.; Subramanian, J. Hexanal vapor induced resistance against major postharvest pathogens of banana (Musa acuminata L.). Plant Pathol. J. 2020, 36, 133–147. [Google Scholar] [CrossRef]
- Patrignani, F.; Iucci, L.; Belletti, N.; Gardini, F.; Guerzoni, M.E.; Lanciotti, R. Effects of sub-lethal concentrations of hexanal and 2-(E)-hexenal on membrane fatty acid composition and volatile compounds of Listeria monocytogenes, Staphylococcus aureus, Salmonella enteritidis and Escherichia coli. Int. J. Food Microbiol. 2008, 123, 1–8. [Google Scholar] [CrossRef]
- Li, Y.; Zhong, S.; Qin, Y.; Zhang, S.; Gao, Z.; Dang, Z.; Pan, W. Identification of plant chemicals attracting and repelling whiteflies. Arthropod-Plant Interact. 2014, 8, 183–190. [Google Scholar] [CrossRef]
- Shiojiri, K.; Ozawa, R.; Matsui, K.; Kishimoto, K.; Kugimiya, S.; Takabayashi, J. Role of the lipoxygenase/lyase pathway of host-food plants in thehost searching behavior of two parasitoid species, Cotesia glomerata and Cotesia plutellae. J. Chem. Ecol. 2006, 32, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Hirao, T.; Okazawa, A.; Harada, K.; Kobayashi, A.; Muranaka, T.; Hirata, K. Green leaf volatiles enhance methyl jasmonate response in Arabidopsis. J. Biosci. Bioeng. 2012, 114, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.S.; Heil, M.; Adame-Alvarez, R.M.; Ballhorn, D.J.; Ryu, C.M. Airborne induction and priming of plant defenses against a bacterialpathogen. Plant Physiol. 2009, 151, 2152–2161. [Google Scholar] [CrossRef] [PubMed]
- Engelberth, J.; Contreras, C.F.; Dalvi, C.; Li, T.; Engelberth, M. Early transcriptome analyses of Z-3-Hexenol-treated zea mays revealed distinct transcriptional networks and anti-herbivore defense potential of green leaf volatiles. PLoS ONE 2013, 8, e77465. [Google Scholar] [CrossRef]
- Wei, J.; Wang, L.; Zhu, J.; Zhang, S.; Nandi, O.I.; Kang, L. Plants attract parasitic wasps to defend themselves against insect pests by releasing hexenol. PLoS ONE 2007, 2, e852. [Google Scholar] [CrossRef]
- Wei, J.; Kang, L. Roles of (Z)-3-hexenol in plant-insect interactions. Plant Signal. Behav. 2011, 6, 369–371. [Google Scholar] [CrossRef]
- Oliveira, I.; Guedes de Pinho, P.; Malheiro, R.; Baptista, P.; Pereira, J.A. Volatile profile of Arbutus unedo L. fruits through ripening stage. Food Chem. 2011, 128, 667–673. [Google Scholar] [CrossRef]
- Mukherjee, K.; Tribedi, P.; Mukhopadhyay, B.; Sil, A.K. Antibacterial activity of long-chain fatty alcohols against mycobacteria. FEMS Microbiol. Lett. 2013, 338, 177–183. [Google Scholar] [CrossRef]
- Havlikova, M.; Bosakova, T.; Petschenka, G.; Cabala, R.; Exnerova, A.; Bosakova, Z. Analysis of defensive secretion of a milkweed bug Lygaeus equestris by 1D GC-MS and GC×GC-MS: Sex differences and host-plant effect. Sci. Rep. 2020, 10, 3092. [Google Scholar] [CrossRef]
- Kim, S.Y.; Shin, D.H. Volatile Constituents from the Leaves of Callicarpa japonica Thunb. and Their Antibacterial Activities. J. Agric. Food Chem. 2004, 52, 781–787. [Google Scholar] [CrossRef]
- Bahder, B.W.; Helmick, E.E. Sampling palms for lethal yellowing and Texas Phoenix palm decline phytoplasmas. In UF/IFAS Extension Document ENY-990; University of Florida: Gainesville, FL, USA, 2018. [Google Scholar]
- Soto, N.; Helmick, E.E.; Harrison, N.A.; Bahder, B.W. Genetic Variability of Palm Lethal Decline Phytoplasmas in the Caribbean Basin and Florida, USA, Based on a Multilocus Analysis. Phytopathology 2021, 111, 2203–2212. [Google Scholar] [CrossRef] [PubMed]
- Terra, L.R.; Ferreira, M.M.C.; Terao, D.; Queiroz, S.C.N. Otimização do processo não invasivo e não destrutivo de extração e análise de metabólitos voláteis de mamão papaia por SPME-GC-MS. Química Nova 2020, 43, 1240–1245. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| qPCR Results | |||||
|---|---|---|---|---|---|
| Palm Sample | Infection Status | TaqMan Ct | HRMA Ct | HRMA Tm (°C) | Result |
| Spa_1 | I | 24.33 | 22.43 | 77.6 | + |
| Spa_2 | NINT | No Ct | No Ct | 64.5 | - |
| Spa_3 | I | 33.97 | 30.21 | 77.6 | + |
| Spa_4 | NIT | No Ct | No Ct | 82.8 | - |
| Spa_5 | NIT | No Ct | No Ct | 82.8 | - |
| Spa_6 | NINT | No Ct | No Ct | 82.8 | - |
| Spa_7 | NIT | No Ct | No Ct | 82.7 | - |
| Spa_8 | NIT | No Ct | No Ct | 82.9 | - |
| Spa_9 | NINT | No Ct | No Ct | 82.7 | - |
| Spa_10 | NIT | No Ct | No Ct | 82.9 | - |
| Spa_11 | NIT | No Ct | No Ct | 82.8 | - |
| Spa_12 | NIT | No Ct | No Ct | 82.8 | - |
| Spa_13 | NIT | No Ct | No Ct | 82.8 | - |
| Spa_14 | NIT | No Ct | No Ct | 82.7 | - |
| Spa_15 | NINT | No Ct | No Ct | 82.9 | - |
| Spa_16 | NINT | No Ct | No Ct | 83.0 | - |
| Spa_17 | NIT | No Ct | No Ct | 82.8 | - |
| Spa_18 | NIT | No Ct | No Ct | 82.7 | - |
| Spa_19 | NIT | No Ct | No Ct | 82.7 | - |
| Spa_20 | NIT | No Ct | No Ct | 82.7 | - |
| Spa_21 | NINT | No Ct | No Ct | 82.8 | - |
| Spa_22 | NIT | No Ct | No Ct | 82.9 | - |
| (+) control | N/A | 23.44 | 21.75 | 77.6 | + |
| (-) control | N/A | No Ct | No Ct | 55.34 | - |
| Rate (°C/min) | T (°C) | Hold Time (min) | Run Time (min) | |
|---|---|---|---|---|
| Initial | 40 | 2 | 2 | |
| Ramp 1 | 5 | 150 | 1 | 25 |
| Ramp 2 | 10 | 250 | 5 | 40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira, J.A.; Ramos, J.A.; Dutra, D.R.C.S.; Di Lella, B.; Helmick, E.E.; Queiroz, S.C.N.; Bahder, B.W. Identification of Green-Leaf Volatiles Released from Cabbage Palms (Sabal palmetto) Infected with the Lethal Bronzing Phytoplasma. Plants 2023, 12, 2164. https://doi.org/10.3390/plants12112164
Ferreira JA, Ramos JA, Dutra DRCS, Di Lella B, Helmick EE, Queiroz SCN, Bahder BW. Identification of Green-Leaf Volatiles Released from Cabbage Palms (Sabal palmetto) Infected with the Lethal Bronzing Phytoplasma. Plants. 2023; 12(11):2164. https://doi.org/10.3390/plants12112164
Chicago/Turabian StyleFerreira, Jordana A., José A. Ramos, Debora R. C. S. Dutra, Brandon Di Lella, Ericka E. Helmick, Sonia C. N. Queiroz, and Brian W. Bahder. 2023. "Identification of Green-Leaf Volatiles Released from Cabbage Palms (Sabal palmetto) Infected with the Lethal Bronzing Phytoplasma" Plants 12, no. 11: 2164. https://doi.org/10.3390/plants12112164
APA StyleFerreira, J. A., Ramos, J. A., Dutra, D. R. C. S., Di Lella, B., Helmick, E. E., Queiroz, S. C. N., & Bahder, B. W. (2023). Identification of Green-Leaf Volatiles Released from Cabbage Palms (Sabal palmetto) Infected with the Lethal Bronzing Phytoplasma. Plants, 12(11), 2164. https://doi.org/10.3390/plants12112164