
Phylogeny and Flow Cytometry of the Genus Kalidium Moq. (Amaranthaceae s.l.) in Kazakhstan

 ,
,
Abstract
:1. Introduction
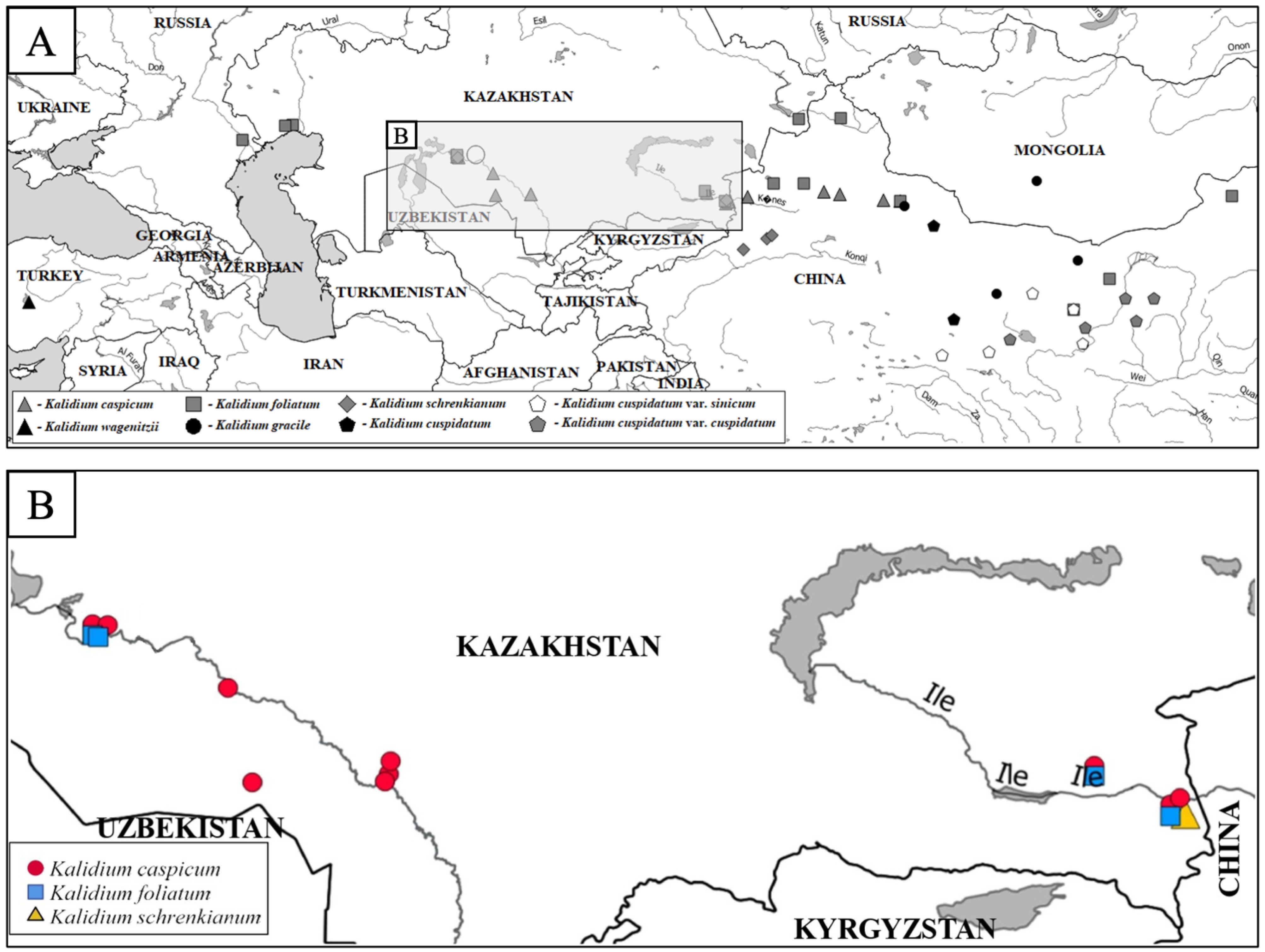
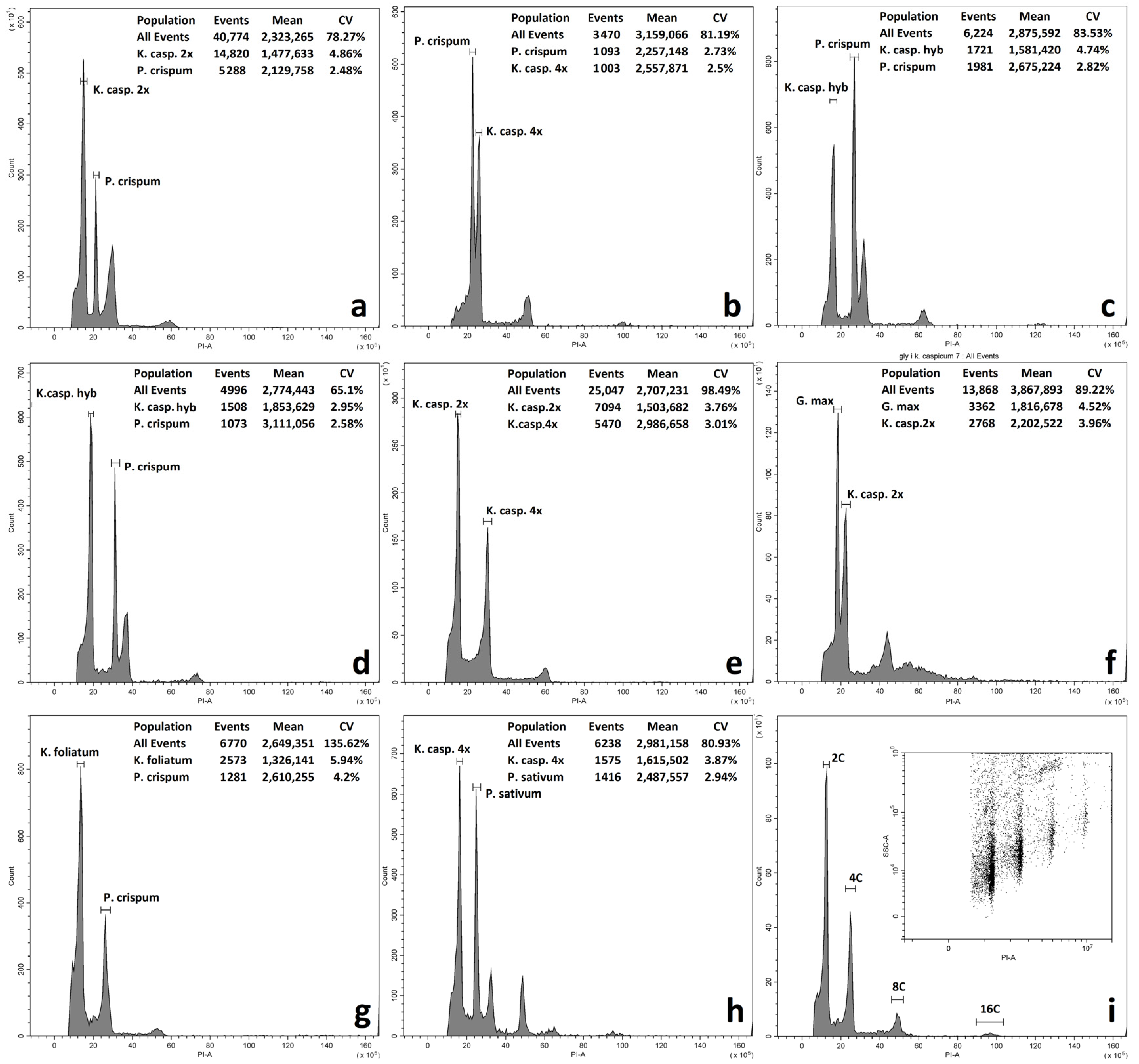
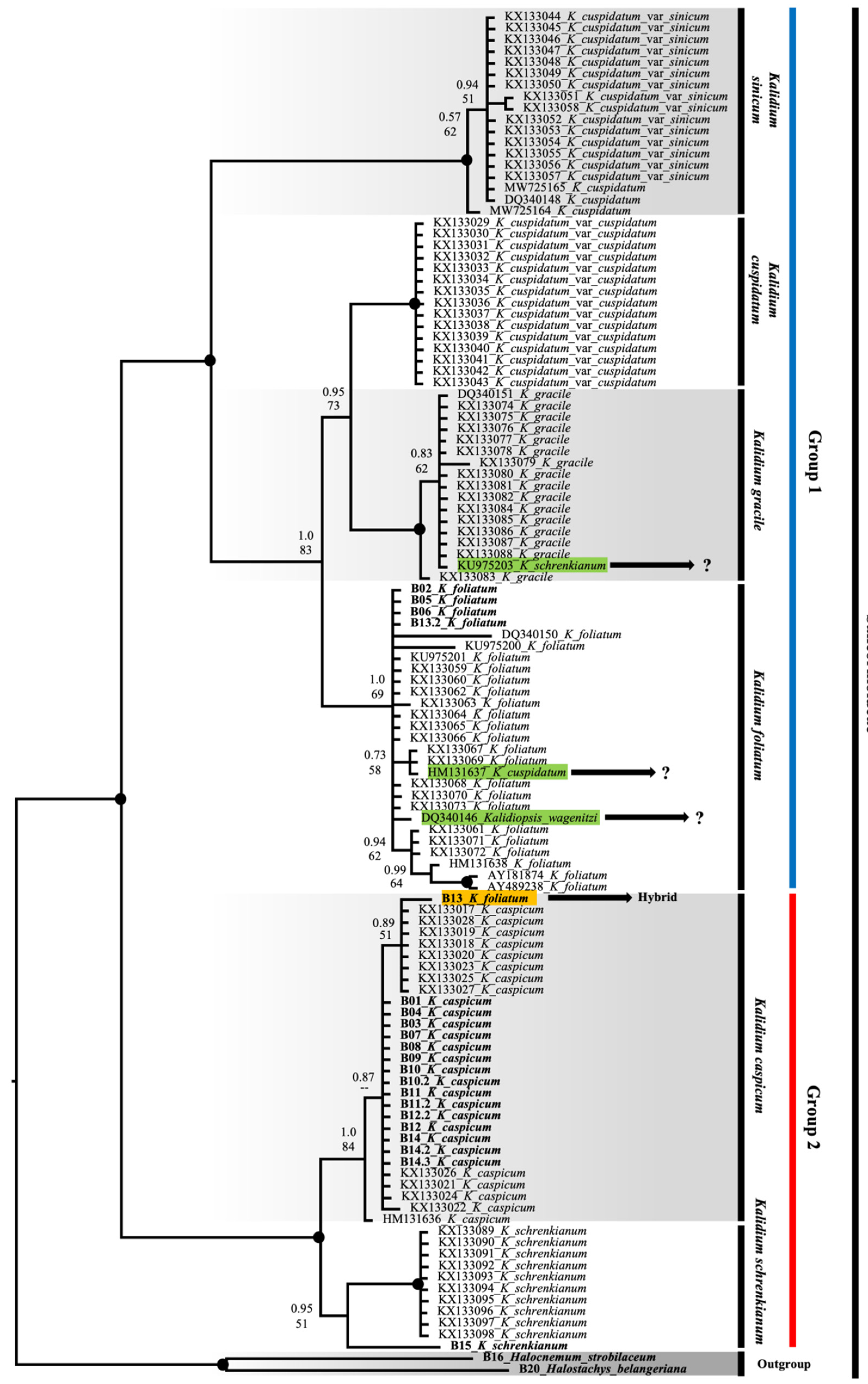
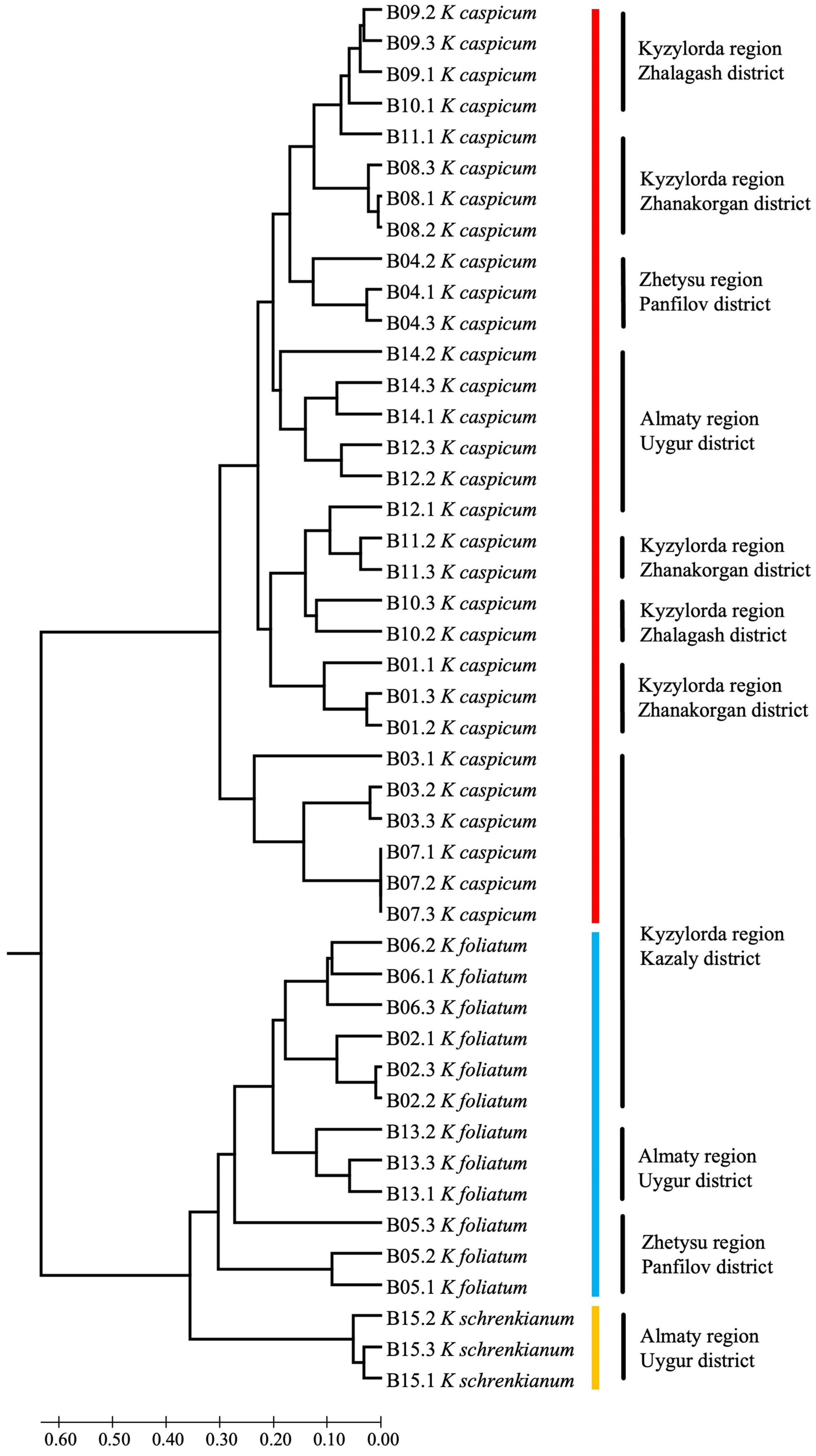
2. Results
2.1. Flow Cytometry
2.2. Molecular Phylogeny
2.3. SCoT Results
3. Discussion
4. Material and Methods
4.1. Flow Cytometry
4.2. Molecular Genetics Methods
4.3. Amplification and Sequencing
4.4. Phylogenetic Analyses
4.5. The Start Codon Targeted (SCoT) Method
5. Conclusions
- -
- The species of the genus Kalidium are divided into two large groups. The first group: K. foliatum, K. gracile, K. wagenitzii, K. sinicum, and K. cuspidatum. The second group: K. caspicum and K. schrenkianum. A promising direction for further study of the genus is additional research on the isolation of superspecific categories.
- -
- Specimens previously attributed to Kalidium cuspidatum variations (K. cuspidatum var. sinicum and K. cuspidatum var. cuspidatum) represent two independent taxa: K. cuspidatum var. sinicum, understood as K. sinicum and K. cuspidatum var. cuspidatum, as K. cuspidatum.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kadereit, G.; Mucina, L.; Freitag, H. Phylogeny of Salicornioideae (Chenopodiaceae): Diversification, biogeography, and evolutionary trends in leaf and flower morphology. Taxon 2006, 55, 6. [Google Scholar] [CrossRef]
- Piirainen, M.; Liebisch, O.; Kadereit, G. Phylogeny, biogeography, systematics and taxonomy of Salicornioideae (Amaranthaceae/Chenopodiaceae)—A cosmopolitan, highly specialized hygrohalophyte lineage dating back to the Oligocene. Taxon 2017, 66, 109–132. [Google Scholar] [CrossRef] [Green Version]
- Vesselova, P.V.; Kudabayeva, G.M. Comparative analysis of floristic lists of the desert part of the Syr Darya valley. Ukr. J. Ecol. 2018, 8, 427–432. [Google Scholar]
- Chapman, V.J. The New Perspective in the Halophytes. Q. Rev. Biol. 1942, 17, 291–311. [Google Scholar] [CrossRef]
- Shreve, F. Vegetation of the Sonoran Desert. Carnegie Inst. Wash. Publ. 1951, 591, 1–192. [Google Scholar]
- Wilder, B.T.; Felger, R.S.; Morales, H.R. Succulent Plant Diversity of the Sonoran Islands, Gulf of California, Mexico. Haseltonia 2008, 14, 127–160. [Google Scholar] [CrossRef]
- Kung, H.-W.; Chu, G.-L.; Tsien, C.-P.; Li, A.-J.; Ma, C.-G. The Chenopodiaceae in China. Acta Phytotaxon. Sin. 1978, 16, 99–123. (In Chinese) [Google Scholar]
- Zhao, Y.-Z.; Zhao, L.-Q.; Cao, R. Flora Intramongolica, 3rd ed.; Typis Intramongolicae Popularis: Huhhot, China, 2020; Volume 1, p. 779. [Google Scholar]
- Sukhorukov, P.; Kushunina, M.A.; Lomonosova, M.N. A new Kalidium species (Amaranthaceae s. l.) from northern Central Asia. Turczaninowia 2022, 25, 24–33. [Google Scholar] [CrossRef]
- Sukhorukov, A.P.; Kushunina, M.A. Taxonomic revision of Chenopodiaceae in Nepal. Phytotaxa 2014, 191, 10–44. [Google Scholar] [CrossRef]
- Zhao, K. Improvement and utiliza- tion of saline soil by planting halophytes. Chin. J. Appl. Environ. Biol. 2002, 8, 31–35. (In Chinese) [Google Scholar]
- Liu, D.; Chang, Z.; Liang, X.; Wu, Y. Soil Chemical Properties Strongly Influence Distributions of Six Kalidium Species in Northwest China. Forests 2022, 13, 2178. [Google Scholar] [CrossRef]
- Liang, X.H.; Wu, Y.X. Identification of Kalidium species (Chenopodiaceae) by DNA barcoding. Sci. Cold Arid. Reg. 2017, 9, 0089–0096. [Google Scholar] [CrossRef]
- Wang, G.; Jia, J. Effects of NaCl on physiology and leaf ultrastructure in the halophyte Kalidium foliatum. Nord. J. Bot. 2015, 33, 232–238. [Google Scholar] [CrossRef]
- Tobe, K. Seed Germination and Radicle Growth of a Halophyte, Kalidium capsicum (Chenopodiaceae). Ann. Bot. 2000, 85, 391–396. [Google Scholar] [CrossRef] [Green Version]
- Tobe, K.; Li, X.; Omasa, K. Effects of sodium, magnesium and calcium salts on seed germination and radicle survival of a halophyte, Kalidium caspicum (Chenopodiaceae). Aust. J. Bot. 2002, 50, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Feng, G.; Zhang, F. Salinity and Temperature Effects on Germination for Three Salt-resistant Euhalophytes, Halostachys caspica, Kalidium foliatum and Halocnemum strobilaceum. Plant Soil 2006, 279, 201–207. [Google Scholar] [CrossRef]
- Qu, X.; Baskin, J.M.; Wang, L.; Huang, Z. Effects of cold stratification, temperature, light and salinity on seed germination and radicle growth of the desert halophyte shrub, Kalidium caspicum (Chenopodiaceae). Plant Growth Regul. 2007, 54, 241–248. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, D.Y.; Huang, Z.Y.; Tian, C.Y. Factors influencing seed germination of Kalidium caspicum (Chenopodiaceae), a halophytic desert shrub of Xinjiang, China. Seed Sci. Technol. 2009, 37, 281–290. [Google Scholar] [CrossRef]
- Gong, D.H.; Wang, G.Z.; Si, W.T.; Zhou, Y.; Liu, Z.; Jia, J. Effects of Salt Stress on Photosynthetic Pigments and Activity of Ribulose-1,5-bisphosphate Carboxylase/Oxygenase in Kalidium foliatum. Russ. J. Plant Physiol. 2018, 65, 98–103. [Google Scholar] [CrossRef]
- Wang, Z.G.; Zhang, P.X.; Shao, Y.T.; Xu, T.T.; Jia, X.Y.; Zhang, X.Q.; Jia, J. Molecular Cloning and the Expression Pattern of a Phospholipid Hydroperoxide Glutathione Peroxidase in Kalidium foliatum under NaCl Treatment. Russ. J. Plant Physiol. 2020, 67, 750–757. [Google Scholar] [CrossRef]
- Wang, J.; Wu, S.; Wang, M. The complete chloroplast genome of halophyte Kalidiumfoliatum (Pall.) Moq., a dominant species of desert grassland. Mitochondrial DNA Part B 2022, 7, 544–545. [Google Scholar] [CrossRef]
- Schutze, P.; Freitag, H.; Weising, K. An integrated molecular and morphological study of the subfamily Suaedoideae Ulbr. (Chenopodiaceae). Plant Syst. Evol. 2003, 239, 257–286. [Google Scholar] [CrossRef]
- Wen, Z.-B.; Zhang, M.-L.; Zhu, G.-L.; Sanderson, S.C. Phylogeny of Salsoleae s.l. (Chenopodiaceae) based on DNA sequence data from ITS, psbB–psbH, and rbcL, with emphasis on taxa of northwestern China. Plant Syst. Evol. 2010, 288, 25–42. [Google Scholar] [CrossRef]
- Xue, J.J.; Zhang, M.L. Monophyly and infrageneric variation of Corispermum L. (Chenopodiaceae), evidence from sequence data psbB-psbH, rbcL and ITS. J. Arid. Land 2011, 3, 240–253. [Google Scholar] [CrossRef]
- Lu, K.-Q.; Li, M.; Wang, G.-H.; Xu, L.-S.; Ferguson, D.K.; Trivedi, A.; Wang, Y.-F. A new pollen classification of Chenopodiaceae for exploring and tracing desert vegetation evolution in the eastern central Asia. J. Syst. Evol. 2019, 57, 190–199. [Google Scholar] [CrossRef]
- Ilyin, M.M. Family Chenopodiaceae in the Flora of the USSR, 6th ed.; Schischkin, V.K., Ed.; Academiae Scientiarum URSS: Mosqua/Leningrad, Russia, 1936; pp. 2–354. (In Russian) [Google Scholar]
- Polyakov, P.P.; Goloskokov, V.P. Family Chenopodiaceae. In The Flora of Kazakhstan, 3rd, ed.; Pavlov, P.P., Ed.; Academy of Sciences of the Kazakh SSR: Alma-Ata, Kazakh Soviet Socialist Republic, 1960; pp. 179–320. (In Russian) [Google Scholar]
- Orasova, A. Family Amaranthaceae. In Illustrated Plant Identifier of Kazakhstan; Goloskokov, V.P., Ed.; Academy of Scences of the Kazakh SSR: Alma-Ata, Kazakh Soviet Socialist Republic, 1969; pp. 271–320. (In Russian) [Google Scholar]
- Pratov, U. Family Chenopodiaceae. In The Plant Identifier of Plants of Central Asia, III; Bondarenko, O.N., Nabiev, M.M., Eds.; FAN Publishing House: Tashkent, USSR, 1972; pp. 29–137. (In Russian) [Google Scholar]
- Abdulina, S.A. Checklist of Vascular Plants of Kazakhstan; Academy of Sciences of the Republic of Kazakstan: Almaty, Kazakhstan, 1998; p. 77. ISBN 9965-01-189-3. (In Russian) [Google Scholar]
- Baitenov, M.S. Flora of Kazakhstan, Genus Complex Flora; Gylym: Almaty, Kazakhstan, 2001; Volume 2, p. 72. ISBN 9965-07-036-9. (In Russian) [Google Scholar]
- Skaptsov, M.V.; Kutsev, M.G. Possibilities of flow cytometry in modern plant science. In Proceedings of the Problems of Botany of South Siberia and Mongolia”-XIII International Scientific-Practical Conference, Barnaul, Russia, 20–23 October 2014; pp. 204–207. (In Russian). [Google Scholar]
- Friesen, N. Molecular Methods Used in Plant Systematics; AzBuka: Barnaul, Russia, 2007; p. 64. (In Russian) [Google Scholar]
- Lomonosova, M.N.; Krasnikov, A.A. Chromosome numbers in some members of the Chenopodiaceae. Bot. Zhurn. 1993, 78, 158–159. [Google Scholar]
- Lomonosova, M.N.; Krasnikov, A.A. Chromosome numbers of some Chenopodiaceae representatives of the flora of Russia. Bot. Zhurn. 2006, 91, 1757–1759. [Google Scholar]
- Krasnikov, A.; Schaulo, D. Karyological study of the Tuva Republic flora: A summary. Turczaninowia 2004, 7, 82–95. [Google Scholar]
- Marhold, K. IAPT/IOPB chromosome data 13. Taxon 2012, 61, 892–893. [Google Scholar] [CrossRef]
- Marhold, K. IAPT/IOPB chromosome data 18. Taxon 2014, 63, 1387–1393. [Google Scholar] [CrossRef] [Green Version]
- An’kova, T.V.; Lomonosova, M.N.; Voronkova, M.S.; Petruk, A.A.; Osmonali, B.; Vesselova, P.V. IAPT chromosome data 32. Taxon 2020, 69, 1126–1132. [Google Scholar] [CrossRef]
- Lomonosova, M.N.; Krasnikov, A.A.; Krasnikova, S.A. Chromosome numbers of Chenopodiaceae family members of the Kazakhstan flora. Bot. Zhurn. 2003, 88, 134–135. [Google Scholar]
- Ghaffari, S.M.; Saydrasi, L.; Ebrahimzadeh, H.; Akhani, H. Chromosome numbers and karyotype analyses of species of subfamily Salicornioideae (Chenopodiaceae) from Iran. Iran. J. Bot. 2006, 12, 128–135. [Google Scholar]
- Zhu, G.; Sergei, L.M.; Stevan, E. Chenopodiaceae: Flora of China; Science Press (Beijing): Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2003; Volume 5, pp. 351–414. [Google Scholar]
- Skvortsov, A.K. Herbarium: A Manual on Methods and Techniques; Prilipko, M.L.I., Ed.; Nauka: Moscow, Russia, 1977; p. 199. (In Russian) [Google Scholar]
- Pfosser, M.; Amon, A.; Lelley, T.; Heberle-Bors, E. Evaluation of sensitivity of flow cytometry in detecting aneuploidy in wheat using disomic and ditelosomic wheat-rye addition lines. Cytometry 1995, 21, 387–393. [Google Scholar] [CrossRef]
- Doležel, J.; Doleželová, M.; Novák, F.J. Flow cytometric estimation of nuclear DNA amount in diploid bananas (Musa acuminata and M. balbisiana). Biol. Plant. 1994, 36, 351–357. [Google Scholar] [CrossRef]
- Yokoya, K.; Roberts, A.V.; Mottley, J.; Lewis, R.; Brandham, P.E. Nuclear DNA amounts in roses. Ann. Bot. 2000, 85, 557–561. [Google Scholar] [CrossRef]
- Doležel, J.; Greilhuber, J.; Lucretti, S.; Meister, A.; Lysák, M.A.; Nardi, L.; Obermayer, R. Plant genome size estimation by flow cytometry: Inter-laboratory comparison. Ann. Bot. 1998, 82, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Blattner, F.R. Direct amplification of the entire ITS region from poorly preserved plant material using recombinant PCR. Biotechniques 1999, 27, 1180–1186. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Shinsky, J.J., White, T.J., Eds.; Academic Press, Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Shaw, J.; Lickey, E.B.; Schilling, E.E.; Small, R.L. Comparison of whole chloroplast genome sequences to choose noncoding regions for phylogenetic studies in angiosperms: The tortoise and the hare III. Am. J. Bot. 2007, 94, 275–288. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The clustal X window interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swofford, D.-L. PAUP*: Phylogenetic Analysis Using Parsimony (* and Other Methods); Version 4; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Kluge, A.G.; Farris, J.S. Quantitative phyletics and the evolution of anurans. Syst. Zool. 1969, 18, 1–32. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Ronquist, R.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertrand, C.; Collard, Y.; Mackill, D.J. Start Codon Targeted (SCoT) Polymorphism: A Simple, Novel DNA Marker Technique for Generating Gene-Targeted Markers in Plants. Plant Mol. Biol. Rep. 2009, 27, 86–93. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Populations | Number of Samples Examined | DNA Content 2C ± SD, pg | Expected Ploidy Level (Literature Consensus) | Number of Chromosomes Based on Literature Data |
|---|---|---|---|---|---|
| K. foliatum | B02, B05, B06 | 9 | 2.259 ± 0.023 | 2× | 18 [35,36,37,38,39,40] |
| K. caspicum | B03, B04, B07 | 9 | 2.981 ± 0.149 | 2× | 18, 36 [41,42] |
| B01, B08, B09, B10, B11 | 11 | 5.993 ± 0.139 | 4× | ||
| K. capsicum × K. foliatum | B07 (putative hybrid) | 4 | 2.616 | 2× | - |
| Pop. | Species | Average Fluorescence Value of the Peak | Index | Expected Ploidy Level |
|---|---|---|---|---|
| B01 | K. caspicum | 2,510,351 | 2.0 | 4× |
| B02 | K. foliatum | 1,091,471 | 1.0 | 2× |
| B03 | K. caspicum | 1,234,585 | 1.0 | 2× |
| B04 | K. caspicum | 1,280,461 | 1.0 | 2× |
| B05 | K. foliatum | 1,137,314 | 1.0 | 2× |
| B06 | K. foliatum | 1,159,205 | 1.1 | 2× |
| B07 | K. caspicum | 1,257,884 | 1.0 | 2× |
| B08 | K. caspicum | 2,575,620 | 2.1 | 4× |
| B09 | K. caspicum. | 2,535,060 | 2.1 | 4× |
| B10 | K. caspicum | 2,499,938 | 2.0 | 4× |
| B11 | K. caspicum | 2,663,129 | 2.2 | 4× |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osmonali, B.B.; Vesselova, P.V.; Kudabayeva, G.M.; Skaptsov, M.V.; Shmakov, A.I.; Friesen, N. Phylogeny and Flow Cytometry of the Genus Kalidium Moq. (Amaranthaceae s.l.) in Kazakhstan. Plants 2023, 12, 2619. https://doi.org/10.3390/plants12142619
Osmonali BB, Vesselova PV, Kudabayeva GM, Skaptsov MV, Shmakov AI, Friesen N. Phylogeny and Flow Cytometry of the Genus Kalidium Moq. (Amaranthaceae s.l.) in Kazakhstan. Plants. 2023; 12(14):2619. https://doi.org/10.3390/plants12142619
Chicago/Turabian StyleOsmonali, B. B., P. V. Vesselova, G. M. Kudabayeva, M. V. Skaptsov, A. I. Shmakov, and N. Friesen. 2023. "Phylogeny and Flow Cytometry of the Genus Kalidium Moq. (Amaranthaceae s.l.) in Kazakhstan" Plants 12, no. 14: 2619. https://doi.org/10.3390/plants12142619
APA StyleOsmonali, B. B., Vesselova, P. V., Kudabayeva, G. M., Skaptsov, M. V., Shmakov, A. I., & Friesen, N. (2023). Phylogeny and Flow Cytometry of the Genus Kalidium Moq. (Amaranthaceae s.l.) in Kazakhstan. Plants, 12(14), 2619. https://doi.org/10.3390/plants12142619