
What Is Wrong with Frankenia nodiflora Lam. (Frankeniaceae)? New Insights into the South African Sea-Heaths

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological and Habitat Studies
2.2. Geographic Coding of the Vouchers and Observations
2.3. Molecular Analyses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Provenance (Herbarium Voucher) | Source | GenBank Code |
|---|---|---|---|
| Tamarix gallica L. | France: Saintes Maries de la Mer (ABH57865) | Villar et al. [30] | MH626294 |
| Myricaria germanica L. | Kazakhstan: Zajsanskaya depression (LE) | Zhang et al. [31] | KJ808607 |
| Reaumuria alternifolia (Labill.) Britten 1 | Azerbaijan: Caucasus (MW) | Zhang et al. [32] | KJ729627 |
| Frankenia anneliseae M.B.Crespo & al. | South Africa: Klipfontein (ABH76891) | This paper | OR183455 |
| South Africa: Skoverfontein (ABH83196) | This paper | OR183456 | |
| F. boissieri Reut. ex Boiss. | Spain: Huelva, Ayamonte, Is. Canela (ABH83543) | This paper | OR183457 |
| F. capitata Webb & Berthel. | Spain: Gran Canaria, Isleta (ABH83612) | This paper | OR183458 |
| F. composita Pau & Font Quer | Morocco: Al Hoceïma, Cala Iris (ABH81590) | This paper | OR183459 |
| F. corymbosa Desf. | Morocco: Al-Hoceïma (ABH54256) | This paper | OR183460 |
| Morocco: Nador, Punta Charrana (ABH54294) | This paper | OR183461 | |
| Spain: Alicante, Santa Pola (ABH79956) | This paper | OR183462 | |
| Spain: Murcia, Cabo Cope (ABH83531) | This paper | OR183463 | |
| F. ericifolia C.Sm. ex DC., nom. cons. prop. | Spain: Tenerife, Punta de Teno (ABH79975) | This paper | OR183464 |
| Spain: Tenerife, Güímar (ABH83613) | This paper | OR183465 | |
| F. fruticosa J.C.Manning & Helme. | South Africa: Moedverloren (ABH76898) | This paper | OR183466 |
| F. hirsuta L. | Türkiye: Tuz Gölii, salty lagoon (ABH45933) | This paper | OR183467 |
| F. ifniensis Caball. | Morocco: Sidi Ifni to Oued Noun (MA758515) | This paper | OR183468 |
| F. laevis L. | France: Aude, Étang de La Palme (ABH70584) | This paper | OR183469 |
| Italy: Sardinia, Cagliari (ABH70072) | This paper | OR183470 | |
| F. nummularia M.B.Crespo & al. | South Africa: Kookfontein River (ABH83290) | This paper | OR183471 |
| South Africa: Tankwa Karoo (ABH83295) | This paper | OR183472 | |
| F. pseudoericifolia Rivas Mart. & al. | Portugal: Cape Verde, São Antão (MA0906845) | This paper | OR183473 |
| F. pulverulenta L. | South Africa: Redelinghuys (ABH77205) | This paper | OR183474 |
| Spain: Teruel, Alcañiz (ABH73564) | This paper | OR183475 | |
| Spain: Alicante, Cabo de las Huertas (ABH41853) | This paper | OR183476 | |
| Spain: Tenerife, Puerto de la Cruz (ABH79974) | This paper | OR183477 | |
| F. repens (P.J.Bergius) Fourc. | South Africa: S of Groenrivier (ABH76868) | This paper | OR183478 |
| South Africa: S of Hondeklipbaai (ABH76862) | This paper | OR183479 | |
| F. thymifolia Desf. | Spain: Zaragoza: Bujaraloz (ABH75454) | This paper | OR183480 |
| Algeria: Bougtob, Chott Cherguí (ABH59344) | This paper | OR183481 | |
| F. velutina Brouss. ex DC. | Morocco: Essaouira (ABH79929) | This paper | OR183482 |
3. Results
3.1. A Brief Story and Typification of Frankenia nodiflora
3.2. Phylogenetic Relationships
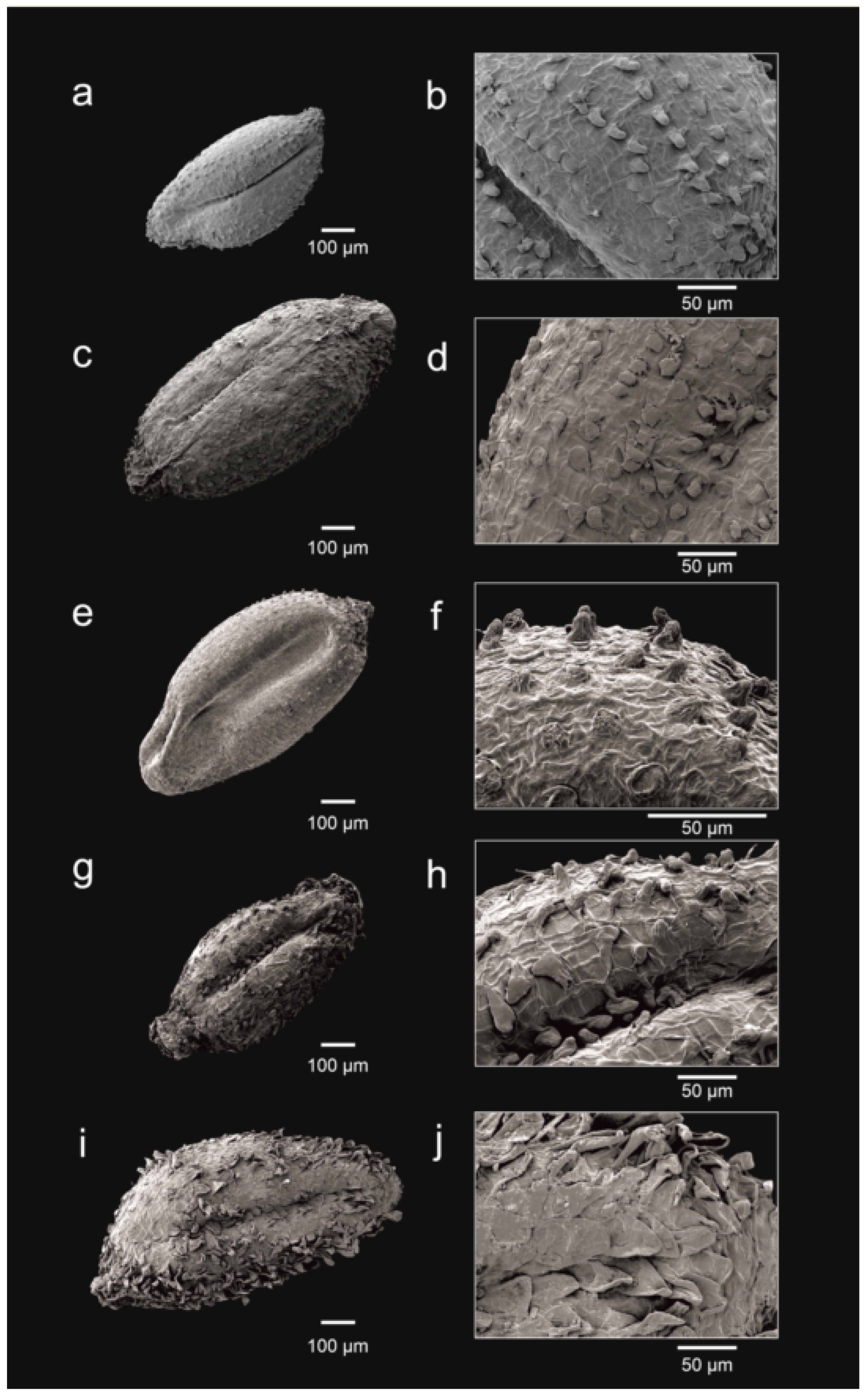
3.3. Number of Ovules and Seed Morphology
3.4. Taxonomic Treatment of Frankenia nodiflora and Description of New Related Species
3.4.1. Frankenia nodiflora Lam., Encycl. 2(2): 543. 1788 ≡ Franca nodiflora (Lam.) Vis. in Mem. Reale Ist. Veneto Sci. 16: 158. 1871.
- -
- F. pulverulenta auct. pl. atque F. krebsii auct. pl.
3.4.2. Frankenia nummularia M.B.Crespo, M.Á.Alonso, Mart.-Azorín, J.L.Villar & Mucina, sp. nov.
- -
- F. pulverulenta auct. pl. atque F. repens auct. pl.
3.4.3. Frankenia anneliseae M.B.Crespo, M.Á.Alonso, Mart.-Azorín, J.L.Villar & Mucina, sp. nov.
- -
- F. capitata auct. pl. atque F. repens auct. pl.
4. Discussion
4.1. Morphological and Evolutionary Relationships
- (i)
- ascendent to erect stems (vs. prostrate, usually rooting stems);
- (ii)
- a smaller (3–6 mm vs. 6–8 mm) calyx;
- (iii)
- a distinct petal length (5.5–8 mm vs. 9–11 mm); and
- (iv)
- a seed testa with smaller papillae (10–43 μm vs. 55–110 μm long), that are heterogeneous in shape, globose to conical-obtuse (vs. subhomogeneous in shape, cylindrical-conical obtuse), and that are sparsely disposed but denser on the distal part (vs. more densely disposed all over the testa surface; Figure 3g,h).
- (i)
- concolorous leaves, which are bright green on both sides (or slightly paler beneath);
- (ii)
- inflorescences that are mostly axillary and glomerular and borne on short lateral opposite branches;
- (iii)
- calyx teeth that are conspicuously acuminate with acumen up to 0.7 mm long;
- (iv)
- an ovary with 4–6 ovules per placenta; and
- (v)
- a seed testa that is sparsely covered with medium-sized papillae that are 12–33 μm long, heterogeneous, and globose to conical-obtuse (Figure 3c,d).
- (i)
- a slender annual habit (vs. suffruticose perennial) with stems and branches that are pubescent on only one side with minute curled hairs (vs. glabrous or covered with minute curled and long flexuous hairs up to 1.2 mm long);
- (ii)
- leaf blade that is long and hairy on the abaxial side (vs. glabrous on both sides);
- (iii)
- flowers that are solitary and scattered along the branches (vs. crowded in loose or dense terminal and/or axillary dichasia);
- (iv)
- petals that are 3.5–5 mm long (vs. 5–9 mm);
- (v)
- a calyx that is 2.5–4(5) mm, puberulous on groves or glabrous (vs. entirely glabrous to sparsely puberulous to hirtellous at base and on ribs), with two bracteoles oblong-linear, as long as or longer than the calyx tube (vs. suborbicular to broadly ovate-oblong, much shorter than the calyx tube); and
- (vi)
- seeds that are very numerous, up to 45 per capsule, ca. 0.5–0.7 mm long (vs. less numerous, up to 22 per capsule, ca. 0.7–1.1 mm long), with testa very sparsely covered with small papillae that are 4–17 μm long, homogeneous, and conical-obtuse (Figure 3a,b).
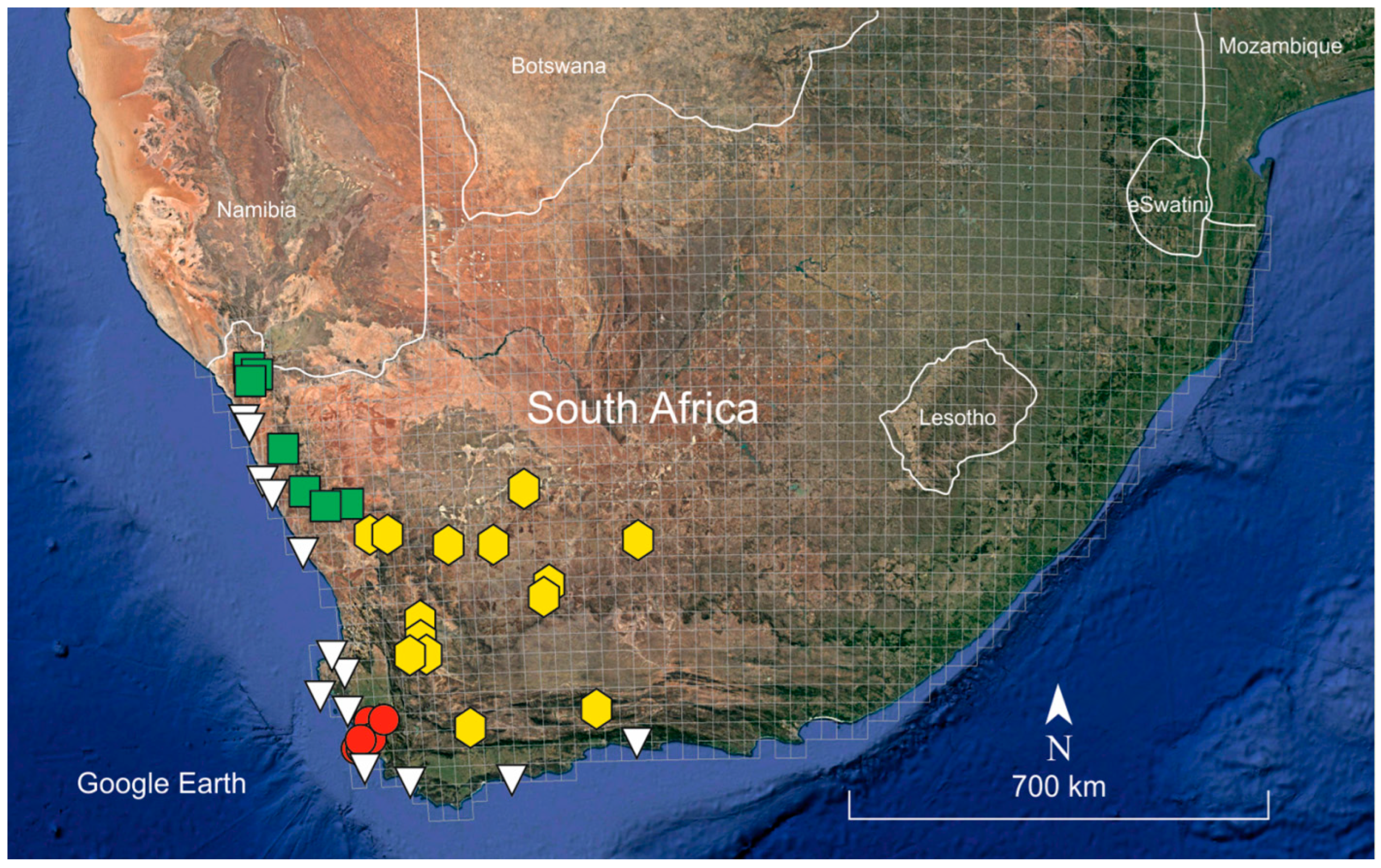
4.2. Phytogeographic and Ecological Patterns
5. Identification Key for Southern African Species of Frankenia
- 1.
- Leaf blade broad, suborbicular to broadly oblong or obovate, flat or longitudinally folded, and sometimes slightly recurved on margins and on the distal half; petiole distinct and 0.5–1.5 mm long......................................................................................................................................................................................2
- -
- Leaf blade linear to linear-lanceolate, mostly strongly recurved on margins, allantoid, or sometimes somewhat flattened; petiole inconspicuous and up to 0.5 mm long ......................................................................................................................................................................................4
- 2.
- Plants annual, delicate. Leaves and bracts hairy beneath. Flowers scattered and solitary in the forks of the branches. Bracteoles as long as or longer than the calyx. Petals 3.5–5 mm long. Seeds up to 45 per capsule, small, 0.5–0.7 mm long................................................................................................................................F. pulverulenta
- -
- Plants suffruticose perennial and shrubby. Leaves and bracts glabrous on both sides. Flowers in terminal dichasia. Bracteoles about half the length of the calyx. Petals 6–9 mm long. Seeds up to 22 per capsule, large, 0.7–1.3 mm long.................................................................................................................................................................3
- 3.
- Leaves of flowering stems broadly ovate and subacute. Inflorescence mostly glomerular and condensed at stem nodes. Calyx 4–4.5 mm long; teeth with a long, conspicuous acumen up to 0.7 mm long. Seed testa sparsely covered with medium-sized papillae 12–33 μm long, heterogeneous, globose to conical-obtuse....................................................................................................................................................................................F. nodiflora
- -
- Leaves of flowering stems suborbicular to elliptic and obtuse. Inflorescence loosely dichasial and usually widely branched. Calyx 4.5–6 mm long; teeth acute or with an inconspicuous mucro up to 0.2 mm long. Seed testa almost smooth with small papillae 3.5–9 μm long only at the base, homogeneous, conical-obtuse....................................................................................................................................................................................F. nummularia
- 4.
- Stems stout, tortuous, and strongly woody at the base. Leaves 1.5–2.5 mm long. Calyx ribs densely puberulous. Petal blade scarcely wider than the claw. Ovules 6, subbasal-parietal....................................................................................................................................................................................F. fruticosa
- -
- Stems thin and suffruticose. Leaves 3–7 mm long. Calyx ribs glabrous or sparsely hairy. Petal blade distinctly wider than the claw. Ovules 15–40, parietal....................................................................................................................................................................................5
- 5.
- Plant sprawling, up to 1 m tall. Stems glabrous. Calyx glabrous. Stamens much shorter than the petals and scarcely protruding from the corolla throat....................................................................................................................................................................................F. pomonensis
- -
- Plant compact, erect to prostrate perennial, and up to 45 cm tall. Stems variously indumented. Calyx puberulent in grooves and often with coarse whitish hairs on the ribs, rarely glabrous. Stamens protruding far from the corolla throat.................................................................................................................................................................6
- 6.
- Stems erect or ascendant, non-rooting, and covered with minute hooked trichomes and longer complanate trichomes. Petals 5.5–8 mm long. Calyx 3–5(–6) mm long. Seeds sparsely covered with medium-sized papillae 10–43 μm long, heterogeneous in shape, globose to conical-obtuse...................................................................................F. anneliseae
- -
- Stems prostrate, often rooting at nodes, and covered with short curly hairs. Petals 9–11 mm long. Calyx 6–8 mm long. Seeds densely covered with large papillae 55–110 μm long, subhomogeneous in shape, cylindrical-conical obtuse............................................................................................................................................................F. repens
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Revised Material of Frankenia repens (s.l.) for Figure 4
References
- Niedenzu, F. Frankeniaceae. In Die Natürlichen Pflanzenfamilien, 2nd ed.; Engler, H.G., Prantl, K.A., Eds.; Wilhelm Engelmann: Leipzig, Germany, 1925; Volume 21, pp. 276–281. [Google Scholar]
- Correa, M.N. Las Frankeniaceae argentinas. Darwiniana 1966, 14, 68–94. [Google Scholar]
- Kubitzki, K. Frankeniaceae. In The Families and Genera of Vascular Plants 5. Malvales, Capparales and Non-Betalain Caryophyllales; Kubitzki, K., Bayer, C., Eds.; Springer: Berlin, Germany, 2003; Volume 5, pp. 209–212. [Google Scholar]
- Obermeyer, A.A. Frankeniaceae. In Flora of Southern Africa; Ross, J.H., Ed.; Botanical Research Institute: Pretoria, South Africa, 1976; Volume 22, pp. 32–36. [Google Scholar]
- Cuénoud, P.; Savolainen, V.; Chatrou, L.W.; Powell, M.; Grayer, R.J.; Chase, M.W. Molecular phylogenetics of Caryophyllales based on nuclear 18S rDNA and plastid rbcL, atpB, and matK DNA sequences. Amer. J. Bot. 2002, 89, 132–144. [Google Scholar] [CrossRef]
- Olson, M.E.; Gaskin, J.F.; Ghahremani-Nejad, F. Stem anatomy is congruent with molecular phylogenies placing Hypericopsis persica in Frankenia (Frankeniaceae): Comments on vasicentric tracheids. Taxon 2003, 52, 525–532. [Google Scholar] [CrossRef] [Green Version]
- Gaskin, J.F.; Ghahremani-Nejad, F.; Zhang, D.-Y.; Londo, J.P. A systematic overview of Frankeniaceae and Tamaricaceae from nuclear rDNA and plastid sequence data. Ann. Missouri. Bot. Gard. 2004, 91, 401–409. [Google Scholar]
- Summerhayes, V.S. A revision of the Australian species of Frankenia. Bot. J. Linn. Soc. 1930, 48, 337–387. [Google Scholar] [CrossRef]
- Heywood, V.H.; Brummitt, R.K.; Culham, A.; Seberg, O. Flowering Plant Families of the World; Royal Botanic Gardens: Kew, UK, 2007. [Google Scholar]
- Manning, J.C.; Helme, N.A. Frankenia fruticosa (Frankeniaceae), a new dwarf shrub from the Knersvlakte, Western Cape. S. Afr. J. Bot. 2014, 91, 84–87. [Google Scholar] [CrossRef]
- Bredenkamp, C.L. Frankeniaceae. In Plants of Southern Africa: An Annotated Checklist; Geminshuizen, G., Meyer, N.L., Eds.; National Botanic Institute: Pretoria, South Africa, 2003; Strelitzia 14; p. 561. [Google Scholar]
- POWO. Plants of the World Online (Continuously Updated). Available online: https://powo.science.kew.org/ (accessed on 3 April 2023).
- de Lamarck, J.-B.P.A.M. Encyclopédie méthodique. Botanique; Panckoucke: Paris, France, 1788; Volume 2, pp. 543–544. Available online: https://www.biodiversitylibrary.org/page/33447031 (accessed on 3 April 2023).
- Mucina, L.; Rutherford, M.C. (Eds.) The Vegetation of South Africa, Lesotho and Swaziland; South African National Biodiversity Institute: Pretoria, South Africa, 2006; Strelitzia 19. [Google Scholar]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff (Continuously Updated). Available online: http://sweetgum.nybg.org/ih/ (accessed on 3 April 2023).
- Rasband, W.S. ImageJ. U.S. National Institutes of Health, Bethesda, Maryland. Available online: https://imagej.nih.gov/ij/ (accessed on 19 March 2020).
- IPNI. The International Plant Names Index. Available online: http://www.ipni.org (accessed on 3 April 2023).
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; et al. (Eds.) International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code) Adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017; Koeltz Scientific Books: Königstein, Germany, 2018. [Google Scholar] [CrossRef]
- Leistner, O.A.; Morris, J.W. Southern African place names. Ann. Cape Prov. Mus. Nat. Hist. 1976, 12, 1–565. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucl. Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbour-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control. 1874, 19, 716–723. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Villar, J.L.; Alonso, M.Á.; Juan, A.; Gaskin, J.; Crespo, M.B. Out of the Middle East: New phylogenetic insights in the genus Tamarix (Tamaricaceae). J. Syst. Evol. 2019, 57, 488–507. [Google Scholar] [CrossRef]
- Zhang, M.-L.; Meng, H.-H.; Zhang, H.-X.; Vyacheslav, B.V.; Sanderson, S.C. Himalayan origin and evolution of Myricaria (Tamaricaeae) in the Neogene. PLoS ONE 2014, 9, e97582. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.-L.; Hao, X.-L.; Sanderson, S.C.; Vyacheslav, B.V.; Sukhorukov, A.P.; Zhang, H.-X. Spatiotemporal evolution of Reaumuria (Tamaricaceae) in Central Asia: Insights from molecular biogeography. Phytotaxa 2014, 167, 89–103. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Sonnerat, P. Voyage aux Indes Orientales et a la Chine, Fait par ordre du Roi, depuis 1774 jusqu’en 1781, 2 vols; l’auteur: Paris, France, 1782. [Google Scholar]
- de Lamarck, J.-B.P.A.M. Tableau Encyclopédique et Méthodique de trois Règnes de la Nature. Botanique. Premiere Livraison; Panckoucke: Paris, France, 1792; Volume 2, pp. 395–396. [Google Scholar]
- de Candolle, A.P. Prodromus Systematis Naturalis Regni Vegetabilis; Treuttel et Würtz: Paris, France, 1824; Volume 1, pp. 349–350. Available online: https://www.biodiversitylibrary.org/page/153953 (accessed on 3 April 2023).
- Harvey, W.H. Order XIV Frankeniaceae St. Hil. In Flora Capensis; Being a Systematic Description of the Plants of the Cape Colony, Caffraria, & Port Natal; Harvey, W.H., Sonder, O.W., Eds.; Hodges, Smith & Co.: Dublin, Ireland, 1860; Volume 1, pp. 114–115. [Google Scholar]
- Roessler, H.; Schreiber, A. Frankeniaceae. In Prodromus einer Flora von Südwestafrika; Merxmüller, H., Ed.; J. Cramer: Lehre, Germany, 1968; Volume 91, pp. 1–2. [Google Scholar]
- Manning, J.C.; Goldblatt, P. (Eds.) Plants of the Greater Cape Floristic Region 1: The Core Cape Flora; South African National Biodiversity Institute: Pretoria, South Africa, 2012; Strelitzia 29; Volume 1, p. 583. [Google Scholar]
- Snijman, D.A. Frankeniaceae. In Plants of the Greater Cape Floristic Region 2: The Extra Cape Flora; Snijman, D.A., Ed.; South African National Biodiversity Institute: Pretoria, South Africa, 2013; Strelitzia 30; Volume 2, pp. 399–400. [Google Scholar]
- IUCN. IUCN Red List Categories and Criteria: Version 3.1, 2nd ed.; UCN Species Survival Commission: Gland, Switzerland; Cambridge, UK, 2012. [Google Scholar]
- Grigore, M.N.; Flowers, T.J. Evolution in gngiosperm halophytes. In Handbook of Halophytes; Grigore, M.N., Ed.; Springer: Cham, Switzerland, 2021; pp. 2117–2146. [Google Scholar] [CrossRef]
- Nègre, R. Les Frankenia du Maroc. Trav. Inst. Sci. Chérifien, Sér. Bot. 1957, 12, 5–56. [Google Scholar]
- Chater, A.O. Frankenia. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1968; Volume 2, pp. 294–295. [Google Scholar]
- Santos, A. Frankenia. In Flora Iberica 3, Plumbaginaceae (Partim)-Capparaceae; Castroviejo, S., Aedo, C., Cirujano, S., Laínz, M., Montserrat, P., Morales, R., Muñoz Garmendia, F., Navarro, C., Paiva, J., Soriano, C., Eds.; Real Jardín Botánico, CSIC: Madrid, Spain, 1993; Volume 3, pp. 446–453. [Google Scholar]
- Drège, J.F. Zwei planzengeographische Documente. Flora 1843, 26, 1–230. [Google Scholar]
- Le Roux, A. Wild Flowers of Namaqualand, 4th rev. ed.; Struik Nature: Cape Town, South Africa, 2015. [Google Scholar]
- Alonso, M.Á.; Crespo, M.B.; Martínez-Azorín, M.; Mucina, L. Morphological and molecular data support recognition of Spergularia quartzicola (Caryophyllaceae) as a new species endemic to South Africa. Plant Biosyst. 2022, 156, 506–514. [Google Scholar] [CrossRef]
- Pohnert, H. Neue Arten aus Südwest-Afrika. Frankeniaceae. Mitt. Bot. Staatssamml. München 1954, 1, 446–448. [Google Scholar]
- Mucina, L.; Jürgens, N.; Le Roux, A.; Rutherford, M.C.; Schmiedel, U.; Esler, K.J.; Powrie, L.W.; Desmet, P.G.; Milton, S.J.; Boucher, C.; et al. Succulent Karoo Biome. In The Vegetation of South Africa, Lesotho and Swaziland; Mucina, L., Rutherford, M.C., Eds.; SANBI: Pretoria, South Africa, 2006; pp. 220–299. [Google Scholar]
- Mucina, L.; Rutherford, M.C.; Palmer, A.R.; Milton, S.J.; Scott, L.; Lloyd, J.W.; van der Merwe, B.; Hoare, D.B.; Bezuidenhout, H.; Vlok, J.H.J.; et al. Nama-Karoo Biome. In The Vegetation of South Africa, Lesotho and Swaziland; Mucina, L., Rutherford, M.C., Eds.; SANBI: Pretoria, South Africa, 2006; pp. 324–347. [Google Scholar]
- Mucina, L. Biomes of the Southern Hemisphere; Springer Nature: Cham, Switzerland, 2023. [Google Scholar] [CrossRef]
- Marais, W. Cruciferae. In Flora of Southern Africa; Codd, L.E., De Winter, B., Killick, D.J.B., Rycroft, H.B., Eds.; Department of Agricultural Technical Services: Pretoria, South Africa, 1970; Volume 13, pp. 1–118. [Google Scholar]
- Mucina, L. Caroxylon (Chenopodiaceae s.str.) in continental southern Africa and Madagascar: A preliminary nomenclatural synopsis and biogeographic considerations. Phytotaxa 2017, 312, 151–178. [Google Scholar] [CrossRef]
- Martínez-Azorín, M.; Crespo, M.B.; Alonso-Vargas, M.Á.; Dold, A.P.; Pinter, M.; Wetschnig, W. Austronea (Asparagaceae, Scilloideae), a new genus from southern Africa, including the description of seven new species. Phytotaxa 2018, 365, 101–129. [Google Scholar] [CrossRef] [Green Version]
- Alonso, M.Á.; Crespo, M.B.; Martínez-Azorín, M.; Mucina, L. Spergularia hanoverensis (Caryophyllaceae): Validation and recircumscription of a misinterpreted species from South Africa. Plants 2023, 12, 2418. [Google Scholar] [CrossRef]
- Köcke, A.V.; von Mering, S.; Mucina, L.; Kadereit, J.W. Revision of the Mediterranean and southern African Triglochin bulbosa complex (Juncaginaceae). Edinb. J. Bot. 2010, 67, 353–398. [Google Scholar] [CrossRef] [Green Version]
- Magee, A.R.; Boatwright, J.S.; Mucina, L. Four new species of Ursinia (Asteraceae, Anthemideae) from South Africa, with an updated key to the genus in Namaqualand. Phytotaxa 2014, 177, 137–145. [Google Scholar] [CrossRef]
- Steffen, S.; Ball, P.; Mucina, L.; Kadereit, G. Phylogeny, biogeography and ecological diversification of Sarcocornia (Salicornioideae, Amaranthaceae). Ann. Bot. 2015, 115, 353–368. [Google Scholar] [CrossRef] [Green Version]
- Mucina, L.; Hammer, T.A. Limonium dagmarae (Plumbaginaceae), a new species from Namaqualand coast, South Africa. Phytotaxa 2019, 403, 71–85. [Google Scholar] [CrossRef]
- Jakoet, A.; Boatwright, J.S.; Mucina, L.; Magee, A.R. A taxonomic revision of the Cotula barbata group (Anthemideae, Cotulinae, Asteraceae). S. Afr. J. Bot. 2022, 151, 495–513. [Google Scholar] [CrossRef]
- Linder, H.P. Plant species radiations: Where, when, why? Philos. Trans. Royal Soc. B Biol. Sci. 2008, 363, 3097–3105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mucina, L.; Kadereit, G.; Steffen, S.; van Niekerk, A. Evolution of flora and vegetation in saline habitats of South Africa. In Proceedings of the MEDECOS XI 2007 Conference, 2–5 September 2007; Rokich, D., Wardell-Johnson, G., Yates, C., Stevens, J., Dixon, K., McLellan, R., Moss, G., Eds.; Kings Park and Botanic Garden: Perth, Australia, 2007; pp. 173–174. [Google Scholar]







| Taxon | Locality | Herbarium Voucher |
|---|---|---|
| F. anneliseae | South Africa: Steinkopf | P05038802 |
| South Africa: Klipfontein | ABH76891 | |
| South Africa: Skoverfontein | ABH83198 | |
| F. nodiflora | South Africa: Cape Town | P05038804 |
| South Africa: Uitkamp | ABH83529 | |
| South Africa: C.T., Salt River | K s.n. | |
| F. nummularia | South Africa: Beukesfontein | P05038725 |
| South Africa: Kookfontein River | ABH83290 | |
| South Africa: Gansfontein | K s.n. | |
| F. pulverulenta | South Africa: Redelinghuys | ABH77205 |
| Spain: Alicante, Jávea | ABH41888 | |
| Spain: Tenerife, Puerto de la Cruz | ABH79974 | |
| F. repens | South Africa: Port Nolloth | ABH76882 |
| South Africa: S of Groenrivier | ABH76868 | |
| South Africa: S of Hondeklipbaai | ABH76862 |
| F. nodiflora | F. nummularia | F. anneliseae | F. repens s.l. | F. pulverulenta s.l. | |
|---|---|---|---|---|---|
| General habit | shrublet, densely branched | shrublet, densely branched | shrub, densely branched | shrublets, lignified at base | annual or rarely short-lived perennial |
| Stem features | rooting only at base, diffuse, creeping prostrate | rooting only at base, procumbent to ascending | erect, densely disposed | mostly rooting at nodes | prostrate to ascending, non-rooting |
| Branchlet indumentum | glabrous to shortly and loosely puberulous all around | glabrous to ± densely hairy mostly on one side | ±densely pubescent all around (rarely glabrescent), | ± densely pubescent all around | ±densely pubescent on one side |
| Branch trichomes: types and length | minute and scattered, up to 0.05 mm, or absent | curled, ca. 0.2 mm, plus flexuous, 0.7–1 mm | claviform or hooked, ca. 0.1–0.2 mm, plus complanate, up to 0.4 mm | minute, curled or hooked, ca. 0.1–0.2 mm | minute, curled, ca. 0.1–0.2 mm |
| Petiole length × width (mm) | 0.6–1.2 × 0.2–0.3 | 0.7–1.5 × 0.3–0.5 | 0.3–0.5 × 0.2–0.5 | 0.5–1.5 × 0.2–0.3 | 0.5–1.5 × 0.2–0.3 |
| Petiole sheath | 2–5 pairs of cilia to subglabrous | 2–5 pairs of cilia | 6–10 pairs of cilia | 3–4 pairs of cilia | 2–6 pairs of cilia |
| Sheath cilia length (mm) and colour | 0.2–0.8, unequal, whitish | 0.3–0.8, unequal, whitish | 0.4–1.2, unequal, whitish | 0.8–2, unequal, whitish | 0.3–0.6, unequal, whitish |
| Leaf blade length × width (mm) | 2.5–3.5 × 1–2 | 2–7 × 1.5–6 | 2–4 × 1–2 | 4–7 × 1–1.5 | 2–5 × 1.5–3 |
| Leaf blade outline and colour | broadly ovate-elliptic to oblong, concolorous (green) | suborbicular or broadly elliptic, often discolorous | broadly oblong to elliptic, often discolorous | narrowly linear, often discolorous | broadly obovate-cuneate, subconcolorous (greyish-green) |
| Leaf blade apex and margins | subacute to mucronulate; gradually revolute | rounded to slightly pointed at apex, loosely and gradually revolute | rounded to subacute at apex, often strongly revolute | subacute at apex, strongly revolute, allantoid | rounded, slightly emarginate, flattened or slightly revolute |
| Leaf blade indumentum | absent (glabrous) on both sides | absent (glabrous) on both sides | absent (glabrous) on both sides | absent (glabrous) or pubescent above, ± densely pubescent beneath | absent (glabrous) above, ± densely pubescent beneath |
| Inflorescence | mostly glomerular, condensed at stem nodes | loosely dichasial, usually widely branched | dense dichasial, usually branched, but compact | dense dichasial, usually branched, but compact | flowers solitary and scattered along branch dichotomies |
| Bracteole length (mm) | 1–2, about half to 2/3 of calyx | 1–2, up to half of calyx | 1–2, about half of calyx | 2–4, about half to 2/3 of calyx | 2.5–4, as long as or longer than calyx |
| Calyx length × width (mm), shape and torsion | 4–4.5 × 1–1.5, tubular, untwisted | 4.5–6 × 1–1.5, fusiform-tubular to fusiform, untwisted | 3–5(–6) × 0.8–1.5, fusiform-tubular to fusiform, untwisted | 6–8 × 1–2, fusiform-tubular to fusiform, untwisted | 2.5–4(–5) × 0.8–1.5, fusiform-tubular to fusiform, untwisted |
| Sepal indumentum (appearance and length) | absent (glabrous) | absent (glabrous) or sparsely puberulous to hirtellous (trichomes up to 0.5 mm) | minutely papillate between ribs, sparsely hirtellous on ribs (trichomes up to 0.2 mm) | absent (glabrous) or puberulous to hirtellous (trichomes up to 2 mm) | absent (glabrous) or minutely papillate between ribs, (trichomes up to 0.2 mm) |
| Calyx teeth length (mm) | 0.9–1.2, with subapical acumen ca. 0.7 mm | 0.7–0.9, acute to mucronate (mucro up to 0.2 mm) | 0.8–1.2, acute to mucronate (mucro ca. 0.2 mm) | 1.2–2, acute to mucronate (mucro ca. 0.2 mm) | 0.4–0.8, acute to mucronate (mucro ca. 0.1 mm) |
| Petal size (mm) and colour | 5–7 × 1–2, pinkish-mauve but whitish below | 6–9 × 1–2, pinkish-mauve | 5.5–8 × 0.9–1.5, whitish to pinkish-mauve or purplish | 9–11 × 2–3.5, pinkish-mauve to purplish | 3.5–5 × 0.6–0.9, whitish-pink to pinkish-mauve |
| Petal blade size (mm), and shape | 2.3–3 × 1.5–2, broadly obovate, rounded to truncate, slightly emarginate, ±sinuate apex | 2.3–4 × 1.5–2, broadly obovate to suborbicular, rounded to truncate, erose-denticulate apex | 2.3–3.5 × 1.5–2, broadly obovate to suborbicular with rounded to truncate, erose-denticulate apex | 4–5 × 2.5–3.5, broadly obovate to suborbicular with rounded, erose-denticulate apex | 2–2.5 × 0.5–0,8, narrowly cuneate to obovate, with truncate, erose-denticulate apex |
| Petal claw (mm) | 2–3 × 0.5–0.7, narrowly cuneate | 4–5 × 0.8–1, narrowly cuneate | 2–3 × 0.6–0.7, narrowly cuneate | 5–6 × 1.5–2, narrowly cuneate | 2–2.5 × 0.3–0.5, cuneate |
| Petal ligule (mm) | 1.5–2.5 × 0.3–0.4, free apex ca. 0.5 mm, oblong-obtuse, entire | 2–3 × 0.5–1, free apex ca. 1 mm, triangular-acute, entire | 1.5–2.5 × 0.3–0.4, free apex ca. 0.5 mm, ovate-acute to acuminate, entire to eroded | 3–4 × 1–1.5, free apex ca. 1.5 mm, ovate-acute to acuminate, entire | 1–2 × 0.2–0.3, free apex ca. 0.4 mm, triangular-acute, entire |
| Stamen filament length (mm) and morphology | 3–6, expanded ca. 0.5 mm in the lower half | 6–8, expanded ca. 0.5 mm in the lower half | 6–8, expanded ca. 0.5 mm in the lower half | 8–11, expanded ca. 0.7 mm in the lower half | 4–6, expanded ca. 0.2 mm in the lower half |
| Anther length (mm), shape and colour | 0.5–0.7, oblong-ellipsoid, yellowish | 0.8–1.2, ellipsoid, yellowish to reddish | 0.4–0.6, ellipsoid, yellow | 0.8–1.4, ellipsoid, yellowish | 0.2–0.4, oblong-ellipsoid, yellowish |
| Ovules per placenta | 4–6 | 6–10 | 5–6 | 7–12 | 12–20 |
| Capsule size (mm) | 2.5–3.5 × 0.6–2 | 2.5–3.5 × 1–2 | 2.5–3.5 × 1–2 | 3.5–5 × 1–2 | 2–3 × 0.5–1.0 |
| Seed number and size (mm) | 9–12, 0.7–1 × 0.3–0.5 | 12–22, 0.7–0.9 × 0.3–0.5 | 1–8, 0.7–1.1 × 0.3–0.4 | 12–22, 0.9–1.3 × 0.4–0.6 | up to 45, 0.5–0.7× 0.2–0.3 |
| Testa papillae length (μm) | 12–33 | 3.5–9 | 10–43 | 55–110 | 4–17 |
| Papillae morphology and distribution | heterogeneous, globose to conical-obtuse, denser on the distal part | subhomogeneous, conical-obtuse, sparse on the funicular part | heterogeneous, globose to conical-obtuse, denser on the distal part | subhomogeneous in shape, cylindrical-conical obtuse, dense all over the testa | homogeneous, conical-obtuse, denser on the distal part |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crespo, M.B.; Alonso, M.Á.; Martínez-Azorín, M.; Villar, J.L.; Mucina, L. What Is Wrong with Frankenia nodiflora Lam. (Frankeniaceae)? New Insights into the South African Sea-Heaths. Plants 2023, 12, 2630. https://doi.org/10.3390/plants12142630
Crespo MB, Alonso MÁ, Martínez-Azorín M, Villar JL, Mucina L. What Is Wrong with Frankenia nodiflora Lam. (Frankeniaceae)? New Insights into the South African Sea-Heaths. Plants. 2023; 12(14):2630. https://doi.org/10.3390/plants12142630
Chicago/Turabian StyleCrespo, Manuel B., María Ángeles Alonso, Mario Martínez-Azorín, José Luis Villar, and Ladislav Mucina. 2023. "What Is Wrong with Frankenia nodiflora Lam. (Frankeniaceae)? New Insights into the South African Sea-Heaths" Plants 12, no. 14: 2630. https://doi.org/10.3390/plants12142630
APA StyleCrespo, M. B., Alonso, M. Á., Martínez-Azorín, M., Villar, J. L., & Mucina, L. (2023). What Is Wrong with Frankenia nodiflora Lam. (Frankeniaceae)? New Insights into the South African Sea-Heaths. Plants, 12(14), 2630. https://doi.org/10.3390/plants12142630