

Phylogenetic Partitioning of Gansu Flora: Unveiling the Core Transitional Zone of Chinese Flora


Abstract
:1. Introduction
2. Results
2.1. Beta Diversity
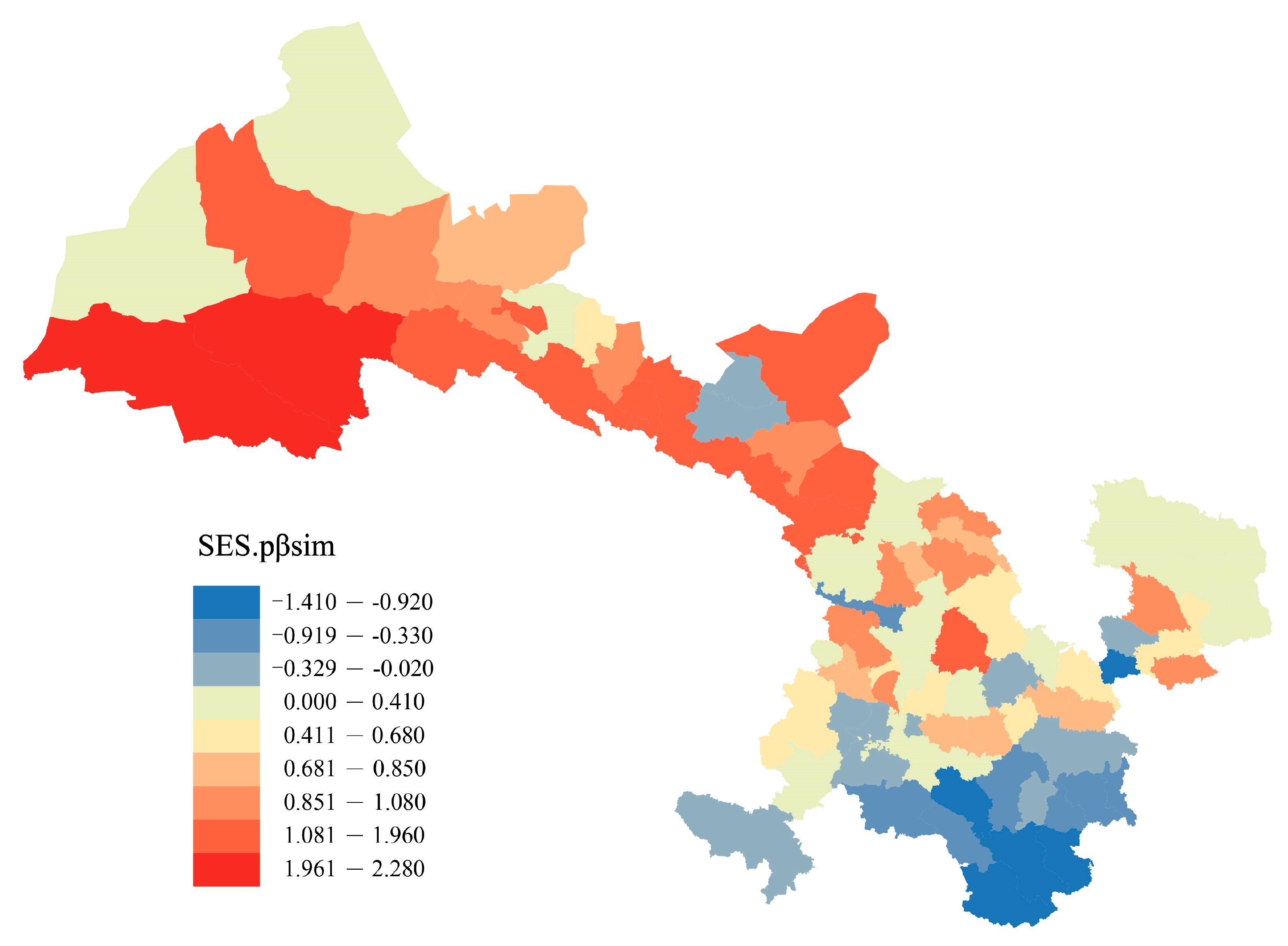
2.2. Floristic Regions
3. Discussion
3.1. Correlation between Phylogenetic and Taxonomic Patterns
3.2. Phylogenetic Delimitation of Floristic Regions
3.3. Ranges and Intersection Boundaries of China’s Four Major Phytogeographic Regions
4. Materials and Methods
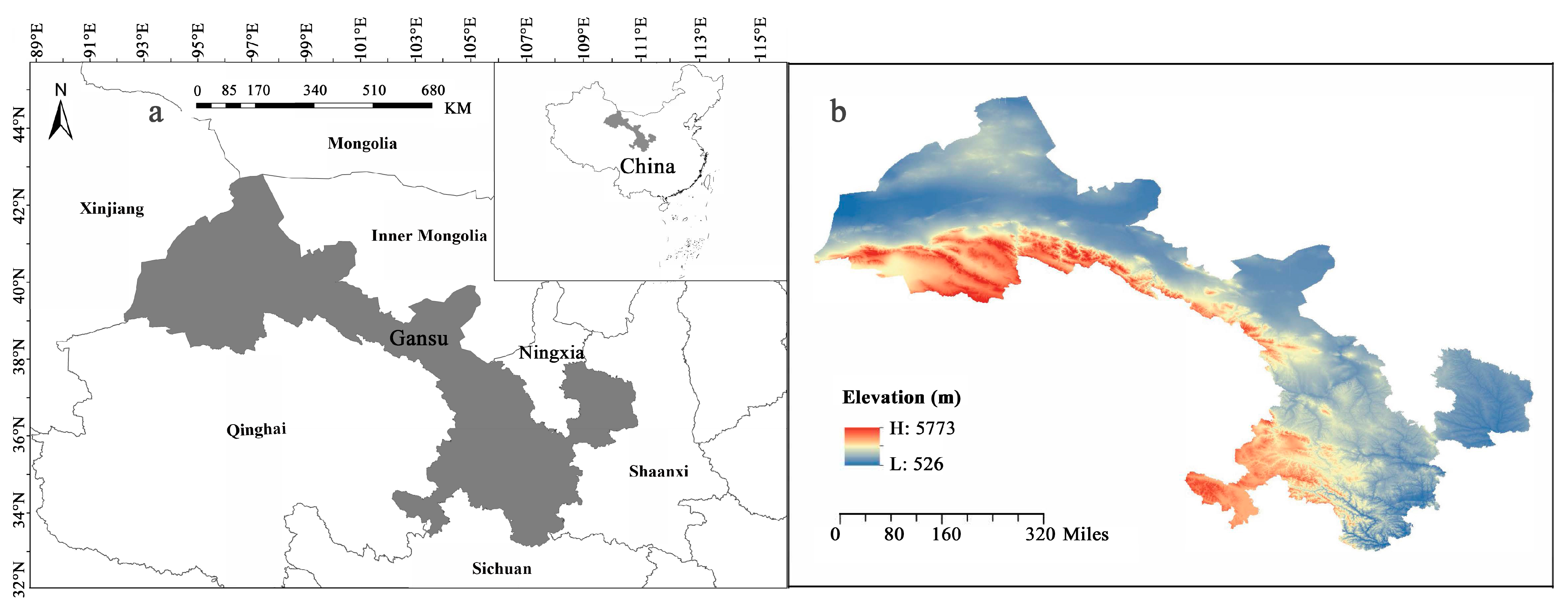
4.1. Study Area
4.2. Species Distribution Data
4.3. Phylogeny Construction
4.4. Phylogeny Diversity Analyses
4.5. Cluster Analysis
4.6. Ordination Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, M.-G.; Slik, J.W.F.; Ma, K.-P. Using species distribution modeling to delineate the botanical richness patterns and phytogeographical regions of China. Sci. Rep. 2016, 6, 22400. [Google Scholar] [CrossRef]
- He, J.; Kreft, H.; Gao, E.; Wang, Z.; Jiang, H. Patterns and drivers of zoogeographical regions of terrestrial vertebrates in China. J. Biogeogr. 2017, 44, 1172–1184. [Google Scholar] [CrossRef]
- Freitas, C.; Brum, F.T.; Cássia-Silva, C.; Maracahipes, L.; Carlucci, M.B.; Collevatti, R.G.; Bacon, C.D. Incongruent Spatial Distribution of Taxonomic, Phylogenetic, and Functional Diversity in Neotropical Cocosoid Palms. Front. For. Glob. Chang. 2021, 4, 739468. [Google Scholar] [CrossRef]
- Li, R.; Kraft, N.J.B.; Yang, J.; Wang, Y. A phylogenetically informed delineation of floristic regions within a biodiversity hotspot in Yunnan, China. Sci. Rep. 2015, 5, srep09396. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Lu, L.; Liu, B.; Yang, T.; Zhang, J.; Hu, H.; Li, R.; Lu, A.; Liu, H.; Mao, L.; et al. Phylogenetic delineation of regional biota: A case study of the Chinese flora. Mol. Phylogenet. Evol. 2019, 135, 222–229. [Google Scholar] [CrossRef] [PubMed]
- González-Orozco, C.E.; Ebach, M.C.; Laffan, S.; Thornhill, A.H.; Knerr, N.J.; Schmidt-Lebuhn, A.N.; Cargill, C.C.; Clements, M.; Nagalingum, N.S.; Mishler, B.D.; et al. Quantifying Phytogeographical Regions of Australia Using Geospatial Turnover in Species Composition. PLoS ONE 2014, 9, e92558. [Google Scholar] [CrossRef]
- Zhang, D.-C.; Ye, J.-X.; Sun, H. Quantitative approaches to identify floristic units and centres of species endemism in the Qinghai-Tibetan Plateau, south-western China. J. Biogeogr. 2016, 43, 2465–2476. [Google Scholar] [CrossRef]
- Thorne, R.F. Phytogeography: Floristic Regions of the World. Armen Takhtajan. Science 1987, 236, 100. [Google Scholar] [CrossRef]
- Takhtajan, A. Floristic Regions of the World; University of California Press: Berkeley, CA, USA, 1986. [Google Scholar]
- Ornduff, R.; Takhtajan, A. The Floristic Regions of the World. Syst. Bot. 1988, 3, 350. [Google Scholar] [CrossRef]
- Wu, Z.Y.; Wu, S.G. Floristic Characteristics and Diversity of East Asian Plants; Zhang, A.L., Wu, S.G., Eds.; China Higher Education Press: Beijing, China, 1996; pp. 3–42. [Google Scholar]
- Wu, Z.Y. The regionalization of Chinese flora. Plant Divers. 1979, 1, 1–20. [Google Scholar]
- Wu, Z.; Sun, H.; Zhou, Z.; Li, D.; Peng, H. Floristics of Seed Plants from China; Science Press: Beijing, China, 2010. [Google Scholar]
- Wu, X.; Chen, X. Analysis of the flora of the family of the seed plants in Kongtong Mountain of Gansu Province. Guihaia 2003, 23, 203–210. [Google Scholar]
- Wang, G.H.; Feng, Z.C. An analysis on the Endemic Elements Occurred to the Woody Plants’ Flora from Gansu Province. J. Gansu Agric. Univ. 1996, 03, 56–61. [Google Scholar]
- Crisp, M.D.; Arroyo, M.T.K.; Cook, L.G.; Gandolfo, M.A.; Jordan, G.J.; McGlone, M.S.; Weston, P.H.; Westoby, M.; Wilf, P.; Linder, H.P. Phylogenetic biome conservatism on a global scale. Nature 2009, 458, 754–756. [Google Scholar] [CrossRef] [PubMed]
- Philippopoulos, K.; Pantavou, K.; Cartalis, C.; Agathangelidis, I.; Mavrakou, T.; Polydoros, A.; Nikolopoulos, G. A novel artificial neural network methodology to produce high-resolution bioclimatic maps using Earth Observation data: A case study for Cyprus. Sci. Total Environ. 2023, 893, 164734. [Google Scholar] [CrossRef]
- Darroch, S.A.F.; Fraser, D.; Casey, M.M. The preservation potential of terrestrial biogeographic patterns. Proc. R. Soc. B Biol. Sci. 2021, 288, 20202927. [Google Scholar] [CrossRef] [PubMed]
- Elliott, T.L.; Davies, T.J. A comparison of phylogenetic and species beta diversity measures describing vegetation assemblages along an elevation gradient. J. Veg. Sci. 2019, 30, 98–107. [Google Scholar] [CrossRef]
- Li, Q.; Sun, H.; Boufford, D.E.; Bartholomew, B.; Fritsch, P.W.; Chen, J.; Deng, T.; Ree, R.H. Grade of Membership models reveal geographical and environmental correlates of floristic structure in a temperate biodiversity hotspot. New Phytol. 2021, 232, 1424–1435. [Google Scholar] [CrossRef]
- White, A.E.; Dey, K.K.; Mohan, D.; Stephens, M.; Price, T.D. Regional influences on community structure across the tropical-temperate divide. Nat. Commun. 2019, 10, 2646. [Google Scholar] [CrossRef]
- Yue, J.; Li, R. Phylogenetic relatedness of woody angiosperm assemblages and its environmental determinants along a subtropical elevational gradient in China. Plant Divers. 2021, 43, 111–116. [Google Scholar] [CrossRef]
- Zhang, X.; Ye, J.; Laffan, S.W.; Mishler, B.D.; Thornhill, A.H.; Lu, L.; Mao, L.; Liu, B.; Chen, Y.; Lu, A.; et al. Spatial phylogenetics of the Chinese angiosperm flora provides insights into endemism and conservation. J. Integr. Plant Biol. 2022, 64, 105–117. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, S.; Hu, G.; Mwachala, G.; Yan, X.; Wang, Q. Species richness and phylogenetic diversity of seed plants across vegetation zones of Mount Kenya, East Africa. Ecol. Evol. 2018, 8, 8930–8939. [Google Scholar] [CrossRef]
- Li, R.; Qian, L.; Sun, H. Current progress and future prospects in phylofloristics. Plant Divers. 2018, 40, 141–146. [Google Scholar] [CrossRef]
- Qian, H.; Zhang, Y.; Zhang, J.; Wang, X. Latitudinal gradients in phylogenetic relatedness of angiosperm trees in North America. Glob. Ecol. Biogeogr. 2013, 22, 1183–1191. [Google Scholar] [CrossRef]
- Li, X.-H.; Zhu, X.-X.; Niu, Y.; Sun, H. Phylogenetic clustering and overdispersion for alpine plants along elevational gradient in the Hengduan Mountains Region, southwest China. J. Syst. Evol. 2014, 52, 280–288. [Google Scholar] [CrossRef]
- Swenson, N.G.; Umaña, M.N. Phylofloristics: An example from the Lesser Antilles. J. Plant Ecol. 2014, 7, 166–175. [Google Scholar] [CrossRef]
- Mishler, B.D.; Knerr, N.; González-Orozco, C.E.; Thornhill, A.H.; Laffan, S.W.; Miller, J.T. Phylogenetic measures of biodiversity and neo- and paleo-endemism in Australian Acacia. Nat. Commun. 2014, 5, 4473. [Google Scholar] [CrossRef] [PubMed]
- Thornhill, A.H.; Mishler, B.D.; Knerr, N.J.; González-Orozco, C.E.; Costion, C.M.; Crayn, D.M.; Laffan, S.W.; Miller, J.T. Continental-scale spatial phylogenetics of Australian angiosperms provides insights into ecology, evolution and conservation. J. Biogeogr. 2016, 43, 2085–2098. [Google Scholar] [CrossRef]
- Scherson, R.A.; Thornhill, A.H.; Urbina-Casanova, R.; Freyman, W.A.; Pliscoff, P.A.; Mishler, B.D. Spatial phylogenetics of the vascular flora of Chile. Mol. Phylogenet. Evol. 2017, 112, 88–95. [Google Scholar] [CrossRef]
- Verboom, G.A.; Archibald, J.K.; Bakker, F.T.; Bellstedt, D.U.; Conrad, F.; Dreyer, L.L.; Forest, F.; Galley, C.; Goldblatt, P.; Henning, J.F.; et al. Origin and diversification of the Greater Cape flora: Ancient species repository, hot-bed of recent radiation, or both? Mol. Phylogenet. Evol. 2009, 51, 44–53. [Google Scholar] [CrossRef]
- Linder, H.P. The evolution of African plant diversity. Front. Ecol. Evol. 2014, 2, 38. [Google Scholar] [CrossRef]
- Chen, Y.S.; Deng, T.; Zhou, Z.; Sun, H. Is the East Asian flora ancient or not? Natl. Sci. Rev. 2018, 5, 920–932. [Google Scholar] [CrossRef]
- Du, W.-B.; Jia, P.; Du, G.-Z. Current patterns of plant diversity and phylogenetic structure on the Kunlun Mountains. Plant Divers. 2022, 44, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Daru, B.H.; Elliott, T.L.; Park, D.S.; Davies, T.J. Understanding the Processes Underpinning Patterns of Phylogenetic Regionalization. Trends Ecol. Evol. 2017, 32, 845–860. [Google Scholar] [CrossRef] [PubMed]
- Moritz, C. Strategies to Protect Biological Diversity and the Evolutionary Processes That Sustain It. Syst. Biol. 2002, 51, 238–254. [Google Scholar] [CrossRef]
- Mace, G.M.; Purvis, A. Evolutionary biology and practical conservation: Bridging a widening gap. Mol. Ecol. 2008, 17, 9–19. [Google Scholar] [CrossRef]
- Winter, M.; Devictor, V.; Schweiger, O. Phylogenetic diversity and nature conservation: Where are we? Trends Ecol. Evol. 2013, 28, 199–204. [Google Scholar] [CrossRef]
- Lyashevska, O.; Farnsworth, K.D. How many dimensions of biodiversity do we need? Ecol. Indic. 2012, 18, 485–492. [Google Scholar] [CrossRef]
- Diniz-Filho, J.A.F.; Loyola, R.D.; Raia, P.; Mooers, A.O.; Bini, L.M. Darwinian shortfalls in biodiversity conservation. Trends Ecol. Evol. 2013, 28, 689–695. [Google Scholar] [CrossRef]
- Costion, C.M.; Edwards, W.; Ford, A.J.; Metcalfe, D.J.; Cross, H.B.; Harrington, M.G.; Richardson, J.E.; Hilbert, D.W.; Lowe, A.J.; Crayn, D.M. Using phylogenetic diversity to identify ancient rain forest refugia and diversification zones in a biodiversity hotspot. Divers. Distrib. 2015, 21, 279–289. [Google Scholar] [CrossRef]
- Lu, L.-M.; Mao, L.-F.; Yang, T.; Ye, J.-F.; Liu, B.; Li, H.-L.; Sun, M.; Miller, J.T.; Mathews, S.; Hu, H.-H.; et al. Evolutionary history of the angiosperm flora of China. Nature 2018, 554, 234–238. [Google Scholar] [CrossRef]
- Graham, C.H.; Fine, P.V.A. Phylogenetic beta diversity: Linking ecological and evolutionary processes across space in time. Ecol. Lett. 2008, 11, 1265–1277. [Google Scholar] [CrossRef] [PubMed]
- Gaston, K.J.; Davies, R.G.; Orme, C.D.L.; Olson, V.A.; Thomas, G.H.; Ding, T.-S.; Rasmussen, P.C.; Lennon, J.J.; Bennett, P.M.; Owens, I.P.; et al. Spatial turnover in the global avifauna. Proc. R. Soc. B Biol. Sci. 2007, 274, 1567–1574. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.J. Survey on medicinal plant resources of Ranunculaceae in Yuzhong county of Gansu province. J. Gansu Univ. Chin. Med. 2018, 35, 5–19. [Google Scholar]
- Zhang, Y.L.; Zhu, Y.H. Geomorphologic zone of Gansu Province. Gansu Geol. 2020, 29, 7–11. [Google Scholar]
- Cui, Z.J.; Jin, L. Research progress of the relationship between compositions and drug properties of Chinese medicine based on group classification. J. Gansu Univ. Chin. Med. 2018, 35, 93–97. [Google Scholar]
- Wu, C.P.; Qu, Y.F.; Chen, X.L.; Cui, Z.J.; Lian, Y.S. The Analysis of Gymnospermous Flora in Gansu Province. Acta Bot. Boreali-Occiden. Sin. 2009, 29, 2084–2089. [Google Scholar]
- Zhang, Z.D.; Yu, Y.W.; Hua, L.M.; Pu, X.; Wang, H.C.; Liang, T.G. Analysis of the distribution pattern of wild vascular plant diversity in Gansu Province, China. Acta Prataculturae Sin. 2014, 23, 22–30. [Google Scholar]
- Shi, J.; Lan, X.; Yu, H.; Wu, H.; Yao, N.; Liang, Y.; Yi, X.; Teng, S. Study on the fern flora in grassland of Gansu Province. Pratacult. Sci. 2016, 33, 1696–1701. [Google Scholar]
- Jalil, N.; Amir, T.; Moslem, D.; Sara, M.; Zahra, A.; Gerald, M.S. Endemic Diversity And Distribution Of The Iranian Vascular Flora Across Phytogeographical Regions, Biodiversity Hotspots And Areas Of Endemism. Sci. Rep. 2019, 9, 12991. [Google Scholar]
- Daniel, J.M.; Malte, C.E.; Joseph, T.M.; Shawn, W.L.; Gerasimos, C.; Visotheary, U.; Andrew, H.T.; Nunzio, K.; Melinda, L.T. Do phytogeographic patterns reveal biomes or biotic regions. Cladistics 2019, 35, 654–670. [Google Scholar]
- Marcusso, G.M.; Kamimura, V.d.A.; Borgiani, R.; Neto, L.M.; Lombardi, J.A. Phytogeographic Meta-Analysis of the Vascular Epiphytes in the Neotropical Region. Bot. Rev. 2022, 88, 388–412. [Google Scholar] [CrossRef]
- Marta, S.; Brunetti, M.; Manenti, R.; Provenzale, A.; Ficetola, G.F. Climate and land-use changes drive biodiversity turnover in arthropod assemblages over 150 years. Nat. Ecol. Evol. 2021, 5, 1291–1300. [Google Scholar] [CrossRef]
- González-Orozco, C.E.; Pollock, L.J.; Thornhill, A.H.; Mishler, B.D.; Knerr, N.; Laffan, S.W.; Miller, J.T.; Rosauer, D.F.; Faith, D.P.; Nipperess, D.A.; et al. Phylogenetic approaches reveal biodiversity threats under climate change. Nat. Clim. Chang. 2016, 6, 1110–1114. [Google Scholar] [CrossRef]
- Feng, J.M. Spatial patterns of species diversity of seed plants in China and their climatic explanation. Biodivers. Sci. 2008, 16, 470–476. [Google Scholar]
- Chen, Z.R.; Zhang, X.X.; Hu, H.H.; Niu, Y.Y.; Ye, J.F.; Zhang, Q.; Liu, B.; Zhao, L.N.; Lu, S.; Lu, L.M. Plant geography in China: History, progress and prospect. Acta Geogr. Sin. 2022, 77, 120–132. [Google Scholar]
- Mao, Z.M.; Chen, J.S.; Peng, E.R.; Liu, Q. Study on temporal and spatial variation trend of vegetation coverage in Gansu Province. J. Jiangxi Agric. 2020, 32, 125–130. [Google Scholar]
- Feng, J.M.; Zhu, Y. Geographical Patterns and Flora Differentiation of Seed Plants in Northwest Yunnan, China. Acta Bot. Boreali-Occiden. Sin. 2009, 29, 2312–2317. [Google Scholar]
- Liu, J.Q.; Zhou, Y.X.; Zhou, D.L.; Wu, H.; Wang, T.H. Flora analysis of the vascular plant families in Gansu Annanba Wild Camel National Nature Reserve. J. Arid. Land Resour. Environ. 2020, 34, 162–167. [Google Scholar]
- Wu, Y.H. Characteristics of Vascular Plant Flora of Altun Mountains. Master’s Thesis, Inner Mongolia University, Hohhot, China, 2021. [Google Scholar]
- Chen, P.; Pan, X.L. The floristic characteristics in the area of the Hexi Corridor. Bull. Bot. Res. 2001, 1, 24–30. [Google Scholar]
- Li, K.P.; Hu, H.; Luo, C.; Qiang, P.; Deng, Y. Study on vegetation composition and distribution characteristics of Wushaoling area in Qilian Mountains. Environ. Ecol. 2020, 2, 79–84+88. [Google Scholar]
- Leprieur, F.; Oikonomou, A. The need for richness-independent measures of turnover when delineating biogeographical regions. J. Biogeogr. 2014, 41, 417–420. [Google Scholar] [CrossRef]
- Teng, S.C.; Zhang, M.; Teng, J.; Qiao, Q. Climatic Change Characteristics in Wushaoling Region of Gansu Province During 1951–2016. J. Arid. Meteorol. 2018, 36, 75–81. [Google Scholar]
- Zhang, Y.; Li, P.; Li, C.X.; Jia, H. Study of halophytes in Hexi area of Gansu. Acta Bot. Boreali-Occiden. Sin. 2003, 23, 115–119. [Google Scholar]
- Hao, S.L.; Pu, X.; Ou, K.W.; Mao, L.Y.; Bin, Z.J.; Long, L.Y.; Liang, C. Comparison of vegetation type changes in Gansu Anxi Extreme Arid Desert National Nature Reserve. Agric. Res. Appl. 2016, 4, 12–15. [Google Scholar]
- Mao, X.W.; Zhang, H.L.; Kong, H. A Study on composition and characterisics of floristic in Xiaolong Mountains. Bull. Bot. Res. 2003, 4, 485–491. [Google Scholar]
- Qu, M.J.; Nueryila, A.; Zhou, X.G.; Zhao, H.; Zhu, W.L.; Wang, J.M.; Li, J.W. Influence of geographic distance and environmental factors on plant β diversity in Alxa Gobi region of northern China. Biodivers. Sci. 2022, 30, 22029. [Google Scholar] [CrossRef]
- Dong, X.R.; Zhang, H.; Zhang, M.G. Explaining the diversity and endemic patterns based on phylogenetic approach for woody plants of the Loess Plateau. Biodivers. Sci. 2019, 27, 1269–1278. [Google Scholar]
- Fujita, A.; Takahashi, D.Y.; Patriota, A.G. A non-parametric method to estimate the number of clusters. Comput. Stat. Data Anal. 2014, 73, 27–39. [Google Scholar] [CrossRef]
- Wolfe, J.A. Some Aspects of Plant Geography of the Northern Hemisphere During the Late Cretaceous and Tertiary. Ann. Mo. Bot. Gard. 1975, 62, 264–279. [Google Scholar] [CrossRef]
- Wen, J. Evolution of Eastern Asian and Eastern North American Disjunct Distributions in Flowering Plants. Annu. Rev. Ecol. Syst. 1999, 30, 421–455. [Google Scholar] [CrossRef]
- López-Pujol, J.; Ren, M.X. China: A hot spot of relict plant taxa. In Biodiversity Hotspots; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2011; pp. 123–137. [Google Scholar]
- Liu, L.; Liang, Y.; Hashimoto, S. Integrated assessment of land-use/coverage changes and their impacts on ecosystem services in Gansu Province, northwest China: Implications for sustainable development goals. Sustain. Sci. 2020, 15, 297–314. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, J.; Su, J.; Sun, W.; Meng, H. Research on a Biodiversity Conservation Value Assessment Method Based on Habitat Suitability of Species: A Case Study in Gansu Province, China. Sustainability 2021, 13, 3007. [Google Scholar] [CrossRef]
- Zhao, P.; He, Z. Temperature Change Characteristics in Gansu Province of China. Atmosphere 2022, 13, 728. [Google Scholar] [CrossRef]
- Rivas-Martínez, S.; Rivas-Saenz, S.; Penas, A. Worldwide Bioclimatic Classification System; Backhuys Pub.: Kerkwerve, The Netherlands, 2002. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 20 May 2022).
- Zhang, J.L.; Liu, B.; Liu, S.; Feng, Z.H.; Jiang, K.W. Plantlist: Looking Up the Status of Plant Scientific Names based on the Plant List Database, Searching the Chinese Names and Making Checklists of Plants. R package Version (0.8.0). 2019. Available online: https://github.com/helixcn/plantlist/ (accessed on 11 May 2022).
- The Angiosperm Phylogeny Group. An Update of the Angiosperm Phylogeny Group Classification for the Orders and Families of Flowering Plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef]
- Jin, Y.; Qian, H.V. PhyloMaker: An R package that can generate very large phylogenies for vascular plants. Ecography 2019, 42, 1353–1359. [Google Scholar] [CrossRef]
- Hardy, O.J.; Couteron, P.; Munoz, F.; Ramesh, B.R.; Pélissier, R. Phylogenetic turnover in tropical tree communities: Impact of environmental filtering, biogeography and mesoclimatic niche conservatism. Glob. Ecol. Biogeogr. 2012, 21, 1007–1016. [Google Scholar] [CrossRef]
- Baselga, A. The relationship between species replacement, dissimilarity derived from nestedness, and nestedness. Glob. Ecol. Biogeogr. 2012, 21, 1223–1232. [Google Scholar] [CrossRef]
- Baselga, A.; Orme, C.D.L. betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Legendre, P.; Fortin, M.-J. Comparison of the Mantel test and alternative approaches for detecting complex multivariate relationships in the spatial analysis of genetic data. Mol. Ecol. Resour. 2010, 10, 831–844. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.; Wagner, H. Vegan: Community Ecology Package; R Package Version. 2.0-10; R Core Team: Helsinki, Finland, 2013. [Google Scholar]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. The comparison of dendrograms by objective methods. Taxon 1962, 11, 33–40. [Google Scholar] [CrossRef]
- Gower, J.C. Comparing Classifications. In Numerical Taxonomy; Springer: Berlin/Heidelberg, Germany, 1983; pp. 137–155. [Google Scholar]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: New York, NY, USA, 2011. [Google Scholar]
- Rousseeuw, P.J. Silhouettes: A graphical aid to the interpretation and validation of cluster analysis. J. Comput. Appl. Math. 1987, 20, 53–65. [Google Scholar] [CrossRef]
- Kruskal, J.B. Nonmetric multidimensional scaling: A numerical method. Psychometrika 1964, 29, 115–129. [Google Scholar] [CrossRef]
- Editorial Committee of Gansu Flora. Flora of Gansu. Lanzhou; Gansu Science Press: Lanzhou, China, 2009; Volumes 1 and 2. [Google Scholar]
- Editorial Committee of Chinese Flora. Flora of China; Beijing Science Press: Beijing, China, 2004. [Google Scholar]
- Lanzhou Institute of Desert Research, China Academy of Sciences. China Desert Flora; Science Publishing: Beijing, China, 1987. [Google Scholar]
- Fu, K.J. Flora of Loess Plateau; Science Press: Beijing, China, 2000. [Google Scholar]
- Liu, L.P.; Ding, G.K. Woody flora of Ziwuling; Lanzhou University Press: Lanzhou, China, 1998. [Google Scholar]
- Gao, W.H. Flora of Kongtong Mountain; Gansu Culture Press: Lanzhou, China, 1998. [Google Scholar]
- An, D.G. Higher Plants in Xiaolongshan; Gansu Ethnic Publishing House: Lanzhou, China, 2002. [Google Scholar]
- Feng, Z.C.; Xu, M.L.; Sun, X.G. Tree Atlas of Gannan; Gansu Science Press: Lanzhou, China, 1994. [Google Scholar]
- Zhao, R.E. Records of Chinese Herbal Medicine Resources in Gansu (Part I); Gansu Science Press: Lanzhou, China, 1994. [Google Scholar]
- Huang, D.S. Vegetation in Gansu; Gansu Science and Technology Press: Lanzhou, China, 1998. [Google Scholar]
- Bai, C.C.; Ma, Z.G. Investigation and study on the ornamental plant resources of Viburnum in Gansu. J. Hortic. 2005, 1, 155–158. [Google Scholar] [CrossRef]
- Cao, J.H. Systematic Taxonomy and Phytogeography of Saussurea in Gansu. Master’s Thesis, Northwest Normal University, Lanzhou, China, 2016. [Google Scholar]
- Chen, X.C. National key protected wild medicinal plant resources in Gansu. China For. Prod. Spec. 2007, 2, 73–76. [Google Scholar]
- Chen, X.C. Distribution and species of Thymelaeaceae woody plants in Gansu. Gansu For. Sci. Technol. 2011, 36, 11–13. [Google Scholar]
- Chen, X.C.; Pei, J.F. Development and utilization of Syringa resources in Gansu. Spec. Econ. Anim. Plant 2017, 20, 25–26. [Google Scholar]
- Chen, X.L. A new record of Syringa plant distribution in Gansu. Gansu For. Sci. Technol. 1988, 2, 33–34. [Google Scholar]
- Chen, X.Q.; Yu, G.D.; Zhou, Y.S. A new plant of Fritillaria in Gansu Province. Yunnan Plant Res. 1985, 2, 146–150. [Google Scholar]
- Cui, Z.J.; Lian, Y.S. A new variety of Meconopsis. Guangxi Bot. 2005, 2, 106–198. [Google Scholar]
- Cui, Z.J.; Lu, M.Z.; Chen, X.L.; Lian, Y.S. Research on endemic genera of angiosperms in China in Gansu Province. Pratacultural Sci. 2009, 26, 30–38. [Google Scholar]
- Cui, Z.J.; Lu, P.L.; Li, S.J.; Jin, L.; Lian, Y.S. Lycium qingshuiheense—A new distribution species of Solanaceae in Gansu Province. China Mod. Chin. Med. 2013, 15, 590+629. [Google Scholar] [CrossRef]
- Dong, Y.; Liu, X.J.; Sun, X.G.; Tian, Q. Four new records of Corydalis plants distribution in Gansu Province. J. Gansu Agric. Univ. 2020, 55, 159–162. [Google Scholar] [CrossRef]
- Duan, L.D.; Lin, Q.; Yang, Z.R.; Shao, Q. New records of Urticaceae plants distribution in twelve provinces of China. Northwest J. Bot. 2011, 31, 1050. [Google Scholar]
- Fang, Z.F.; Wang, J.Q. A new plant of Salix from Gansu. Bull. Bot. Res. 1981, 4, 123–127. [Google Scholar]
- Feng, Z.C. Several new records of woody plants in Gansu. J. Gansu For. Sci. Technol. 1985, 1, 35–36. [Google Scholar]
- Feng, Z.C.; Sun, X.G. A new species of Rhododendron in Gansu. Plant Res. 1992, 12, 145–146. [Google Scholar]
- Gao, J.F. A preliminary report on the investigation of Taxus wallichiana var. chinensis resources in Gannan Prefecture. Spec. Econ. Anim. Plant 2015, 18, 41–42. [Google Scholar]
- Guo, Y.X.; Jin, L.; Cui, Z.J.; Wang, Z.H.; Ma, Y.; Pu, X. Five new records species of angiosperms distribution in Gansu Province. Chin. Mod. Tradit. Chin. Med. 2020, 22, 21–25. [Google Scholar] [CrossRef]
- Guo, Y.X.; Pu, X.; Zheng, G.S.; Cui, Z.J.; Wang, Z.H.; Ma, Y.; Jin, L. Two new records species of angiosperms distribution in Gansu. Chin. Wild Plant Resour. 2021, 40, 69–70. [Google Scholar]
- Li, B.; Zhang, K.N.; Gong, H.D. Investigation on Tibetan medicinal plant resources of Liliaceae in Gan Nan area of Qinghai-Tibet Plateau. J. Ningxia Norm. Univ. 2017, 38, 57–61+65. [Google Scholar]
- Li, B.; Gong, H.D.; Dong, G.; Zhang, K.N. Investigation on Tibetan medicinal plant resources of Scrophulariaceae in Gan Nan area of Qinghai-Tibet Plateau. Heilongjiang Anim. Husb. Vet. Med. 2017, 7, 192–194. [Google Scholar] [CrossRef]
- Li, B.; Dong, G.; Gong, H.D. Investigation and study on the resources of Papaveraceae Tibetan medicinal plants in Gannan area of Qinghai-Tibet Plateau. J. Ningxia Norm. Univ. 2018, 39, 66–70. [Google Scholar]
- Li, B.; Dong, G.; Gong, Y.Q.; Lu, X.S. Investigation and study on the resources of Apiaceae Tibetan medicinal plants in Gannan area of Qinghai-Tibet Plateau. J. Gansu High. Teach. Coll. 2016, 21, 43–46. [Google Scholar]
- Li, B.; Dong, G.; Ma, G.H.; Li, Z.J. Investigation and study on the resources of Caryophyllaceae Tibetan medicinal plants in Gannan area of Qinghai-Tibet Plateau. J. Chuxiong Norm. Univ. 2017, 32, 42–44+50. [Google Scholar]
- Li, B.; Dong, G.; Yu, C.C. Investigation and study on the resources of Fabaceae Tibetan medicinal plants in Gannan area. J. Mudanjiang Norm. Univ. 2018, 3, 62–65. [Google Scholar]
- Li, B.; Gong, H.D.; Dong, G. Investigation on Tibetan medicinal plant resources of Geraniaceae in Gan Nan area of Qinghai-Tibet Plateau. J. Mudanjiang Norm. Univ. 2017, 4, 52–54. [Google Scholar] [CrossRef]
- Li, B.; Li, Z.L.; Luo, Y.L.; Gong, H.D. Investigation and study on the resources of Compositae Tibetan medicinal plants in Gannan area of Qinghai-Tibet Plateau. J. Ningxia Norm. Univ. 2018, 39, 78–87. [Google Scholar]
- Li, B.; Li, Z.J.; Gong, H.D. Investigation and study on the resources of Gentianaceae Tibetan medicinal plants in Gannan area of Qinghai-Tibet Plateau. J. Qujing Norm. Univ. 2017, 36, 40–43. [Google Scholar]
- Li, B.; Lu, X.S.; Gong, H.D.; Ma, G.H. Investigation on Tibetan medicinal plant resources of Labiatae in Gan Nan area of Qinghai-Tibet Plateau. J. Mudanjiang Norm. Univ. 2016, 4, 43–46. [Google Scholar] [CrossRef]
- Li, B.; Luo, Y.L.; Li, Z.L. Investigation and study on the resources of Rosaceae Tibetan medicinal plants in Gannan area. Plateau Sci. Res. 2018, 2, 28–33. [Google Scholar] [CrossRef]
- Li, B.; Zhou, T.L.; Yu, C.C. Investigation on Tibetan medicinal plant resources of Solanaceae in Gan Nan area of Qinghai-Tibet Plateau. J. Ningde Norm. Univ. 2018, 30, 139–142. [Google Scholar] [CrossRef]
- Li, J.J.; Li, Y.W.; Hu, X.F.; Zhao, C.X.; Zhang, J.H.; Yang, J. Eleven new records and utilization of wild plants in Yunping Forest Farm of Xiaolongshan Mountain in Gansu Province. Spec. Econ. Anim. Plant J. 2019, 22, 25–27+30. [Google Scholar]
- Liu, L.P.; Lian, Y.S. A new species of Wikstroemia in Gansu. Acta Phytotaxon. Sin. 1996, 4, 440–442. [Google Scholar]
- Liu, P.L.; Du, C.; Lu, Y.; Jiang, Z.M.; Wen, J.L. New plant distribution records in Gansu Province. Acta Bot. Boreali-Occident. Sin. 2013, 33, 194–196. [Google Scholar]
- Liu, X.J.; Wang, J.H.; Sun, X.G.; Zhang, H.Q.; Chen, X.D.; Li, W.X. Three new recorded species of Orchidaceae in Gansu Province. J. Gansu Agric. Univ. 2016, 51, 85–87+94. [Google Scholar] [CrossRef]
- Liu, X.J.; Wang, J.H.; Sun, X.G.; Zhang, H.Q.; Chen, X.D.; Li, W.X. Three new recorded genera of Asteraceae in Gansu Province. Acta Bot. Boreali-Occident. Sin. 2015, 35, 1273. [Google Scholar]
- Liu, X.L.; Mao, W.X.; Tian, Q.; Sun, X.G.; Liu, X.J. Investigation of Cypripedium resources and two new recorded species in Gansu Province. Acta Bot. Boreali-Occident. Sin. 2021, 41, 533–538. [Google Scholar]
- Lu, Y.; Sun, Y.F.; Liu, P.L.; Du, C.; Jiang, Z.M. Newly recorded plants to Qinling Mountains, Gansu. Shaanxi For. Sci. Technol. 2017, 5, 64–66. [Google Scholar]
- Ma, C.S.; Huang, H.L.; Zhang, G.Z.; Zhang, L.H.; Jin, Q.Y.; Yang, Y.X. New records of Scrophulariaceae in Gansu Province and their geographical distribution. Gansu For. Sci. Technol. 2017, 42, 7–8. [Google Scholar]
- Ma, H.S.; Chen, X.L.; Wang, Y.B. A new record plant of Xanthium in Gansu and its taxonomic key. Anhui Agric. Sci. 2012, 40, 11–12. [Google Scholar] [CrossRef]
- Ma, W.B. Study on Medicinal Plant Resources and Diversity in Maqu County. Master’s Thesis, Northwest Normal University, Lanzhou, China, 2017. [Google Scholar]
- Ma, B.; Chen, X.L.; Liu, L.; Da Q j Zhang, H.X.; Liu, Z.H. New records of angiosperm distribution in Gansu Province. Guangxi Bot. 2017, 37, 220–224. [Google Scholar] [CrossRef]
- Ma, X.H.; Jin, L.; Lv, P.L.; Huang, D.D.; Zhu, T.T. Sorting out the species and varieties of Tibetan medicine in Gentianaceae in Gannan area of Gansu Province. Chin. Vet. Med. J. 2018, 37, 86–88. [Google Scholar] [CrossRef]
- Ma, X. Investigation on medicinal fern resources in Gannan Plateau. J. Xinyang Norm. Univ. 2012, 25, 486–489. [Google Scholar]
- Ma, Y.; Guo, Y.X.; Lin, L.; Cui, Z.J.; Jin, L. Investigation on medicinal plant resources in Linxia County of Gansu Province. Chin. J. Tradit. Chin. Med. Inf. 2021, 28, 1–4. [Google Scholar] [CrossRef]
- Ma, Z.L.; Zhou, X.C. Regional distribution characteristics and protection and utilization of medicinal plant resources in Gansu. China For. By-Prod. Spec. 2009, 2, 72–74. [Google Scholar]
- Mao, W.X.; Sun, X.G.; Liu, X.J. New records of Gentianaceae medicinal plants distribution in Gansu Province. For. Sci. Technol. Commun. 2018, 9, 77–79. [Google Scholar] [CrossRef]
- Pan, S.C.; Zou, T.F.; Zhang, X.Y.; Wang, R.X.; Sun, S.; Liu, R.; Wang, C.L.; Wang, C.Y.; Chen, L.; Qi, J. Investigation on the resources of medicinal plant Sinopodophyllum hexandrum in Xinglong Mountain Nature Reserve in Gansu Province. For. Sci. Technol. Commun. 2017, 12, 37–39. [Google Scholar] [CrossRef]
- Peng, W.D.; Peng, Z.X. A new species of Ligularia from Gansu. Acta Phytotaxon. Sin. 1995, 6, 612–613. [Google Scholar]
- Pu, M.; Wang, J.H. A new record species of Orchidaceae in Gansu Province-Oreorchis foliosa var. indica. Gansu For. Sci. Technol. 2019, 44, 51–52+2. [Google Scholar]
- Ren, J.W.; Lei, Y.; Zhang, G.L. New records of Clematoclethra in Gansu. J. Gansu For. Sci. Technol. 1993, 1, 48. [Google Scholar]
- Shi, C.K.; Liu, X.J.; Sun, X.G. Four new records of Orchidaceae plants in Gansu Province. J. Gansu Agric. Univ. 2008, 1, 130–132. [Google Scholar]
- Su, J.J.; Chen, X.L.; Hou, Q.Z.; Yan, M.S.; Ma, Z.X. New data of seed plants in Gansu Province. Northwest J. Bot. 2012, 32, 622–623. [Google Scholar]
- Sun, X.G. Species and distribution of Rhododendron simsii in Gansu. Gansu For. Sci. Technol. 1996, 1, 76–78. [Google Scholar]
- Sun, X.G.; Chen, Y.P.; Wang, Z.T.; Zhang, X.C.; Zhang, S.R. New data on vascular plants in Gannan forest area. Gansu For. Sci. Technol. 2001, 4, 7–8. [Google Scholar]
- Sun, X.G.; Huang, H.L.; Shi, D.Z. New records of seed plants in Gansu. Gansu For. Sci. Technol. 1999, 1, 24–25. [Google Scholar]
- Tian, F.; Chen, X.L. Resources and conservation of rare and endangered medicinal plants in Gansu. Chin. J. Vet. Med. 2009, 28, 77–79. [Google Scholar] [CrossRef]
- Wang, L.Z. Quantitative analysis of Rosa plants in Gansu. J. Northwest Norm. Univ. Nat. Sci. Ed. 1990, 1, 78–84. [Google Scholar]
- Wang, Y.F.; Lian, Y.S.; Wang, G.H. A new species of Dipelta in Gansu Province. Northwest J. Bot. 1994, 6, 91–92. [Google Scholar]
- Wang, Y.F.; Wang, J.L. Study on Chenopodiaceae plants in Gansu. Plant Res. 2006, 6, 742–749. [Google Scholar]
- Wu, T. Germplasm Resources of Spermatophyte in Dunhuang Westlake National Nature Reserve of Gansu. Shaanxi For. Sci. Technol. 2020, 48, 17–21. [Google Scholar]
- Wu, X.J. Analysis of Seed Plant Flora of Kongtong Mountain in Gansu. Master’s Thesis, Northwest Normal University, Lanzhou, China, 2003. [Google Scholar]
- Xu, W.B. New records of plant distribution in southeastern Gansu Province. J. Nanjing For. Univ. Nat. Sci. Ed. 2014, 38, 65–66. [Google Scholar]
- Xu, R.; Zhao, Y.; Zhu, Y.J.; Chen, L.; Zang, C.X.; Zhao, L.Q. New records of vascular plants in the northern mountain regions of Hexi Corridor and Gansu. Arid Zone Resour. Environ. 2018, 32, 181–186. [Google Scholar] [CrossRef]
- Yan, M.S.; Yang, X.L. A new plant of Brassicaceae from Gansu. J. Northwest Norm. Univ. 1992, 4, 84–86. [Google Scholar]
- Yang, C.J.; Liu, X.J.; Wang, J.H.; Li, W.X.; Zhang, H.Q.; Wang, F.; Sun, X.G. Four new records genera of angiosperms in Gansu Province. J. Gansu Agric. Univ. 2017, 52, 133–137. [Google Scholar] [CrossRef]
- Yang, F.X. Investigation and Analysis of Tibetan Medicinal plant Resources in Xiahe County. Master’s Thesis, Gansu Agricultural University, Lanzhou, China, 2018. [Google Scholar]
- Yang, T.; Ning, Y.M.; Jin, L. A new record genus of Orchidaceae in Gansu Province—Frigidorchis. Northwest J. Bot. 2017, 37, 812–814. [Google Scholar]
- Yao, D.S.; Chen, X.C.; Ren, J.W. A new species of Periploca in Gansu. Plant Res. 2002, 3, 259–260. [Google Scholar]
- Yao, D.S.; Yao, Y. Species and garden application of Rosa in Gansu. For. Sci. Technol. Newsl. 2015, 6, 54–55. [Google Scholar] [CrossRef]
- Ye, W.B.; Huang, Z.H.; Zhang, F.L.; Liu, F.S.; Pang, S.W. Preliminary investigation and habitat analysis of the new Gansu record—Balanophora henryi and related medicinal plants. J. Cap. Norm. Univ. Nat. Sci. Ed. 2016, 37, 50–54. [Google Scholar] [CrossRef]
- Zhan, T.J.; Zhao, D.M.; Zhang, J.; Li, F.H.; Xu, M.; Zhao, B.; Jia, W.H. Investigation on the resources of medicinal gymnosperms and pteridophytes in Minqin Liangucheng National Nature Reserve in Gansu Province. Gansu Sci. Technol. 2017, 33, 129–130+126. [Google Scholar]
- Zhang, J.; Zhao, D.M.; Hu, S.X.; Gao, W.L.; Zhao, Q.Y.; Xie, R.D.; Pan, C.H. Medicinal plant resources of Zygophyllaceae and Convolvulaceae in Lian Gu Cheng National Nature Reserve of Minqin County, Gansu Province. Gansu Sci. Technol. 2017, 33, 138–140. [Google Scholar]
- Zhang, Y.J.; Cheng, L. Taxonomic study on Fritillaria in Gansu. J. Lanzhou Univ. 1998, 2, 88–95. [Google Scholar]
- Zhang, Y.J.; Peng, Z.X.; Zhang, G. A new species of Fagaceae from Gansu. Acta Bot. Yunnanica 1994, 2, 121–122. [Google Scholar]
- Zhang, Y.B.; Dou, Z.G.; Yang, H.R.; Wang, H.; Li, C.H.; Sun, X.G.; Zhang, Z.Y.; Liu, X.J. Nine new records of angiosperms in Gansu Province. J. Gansu Agric. Univ. 2012, 47, 80–84. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Sun, X.G.; Shi, C.K. Five new records of Ericaceae in Gansu Province. J. Gansu Agric. Univ. 2006, 6, 86–88. [Google Scholar]
- Zhang, Z.Y.; Zhang, Y.B.; Liu, X.J.; Yan, C.C.; Wang, Y.M.; Dong, W.T.; Fu, H.Y.; Ma, Y.X.; Yu, H.Z.; Sun, X.G. Eight new records of angiosperms in Gansu Province. J. Gansu Agric. Univ. 2011, 46, 77–80. [Google Scholar] [CrossRef]
- Zhao, D.M.; Zhao, Q.Y.; Hu, S.X.; Gao, W.L.; Wang, R.Y.; Yang, Y.T.; Zhang, J. Study on the resources of medicinal plants of Tamaricaceae and Brassicaceae in Minqin Liangucheng National Nature Reserve in Gansu Province. Gansu Sci. Technol. 2017, 33, 131–133. [Google Scholar]
- Zhao, J.L.; Sun, X.G.; Chen, X.D.; Zhang, H.Q.; Li, W.X. Seven new records species of angiosperms distribution in Gansu Province. J. Gansu Agric. Univ. 2014, 49, 118–121+126. [Google Scholar] [CrossRef]
- Zhu, J.N. Systematic Taxonomy and Phytogeography of Primulaceae in Gansu Province. Master’s Thesis, Gansu Agricultural University, Lanzhou, China, 2013. [Google Scholar]






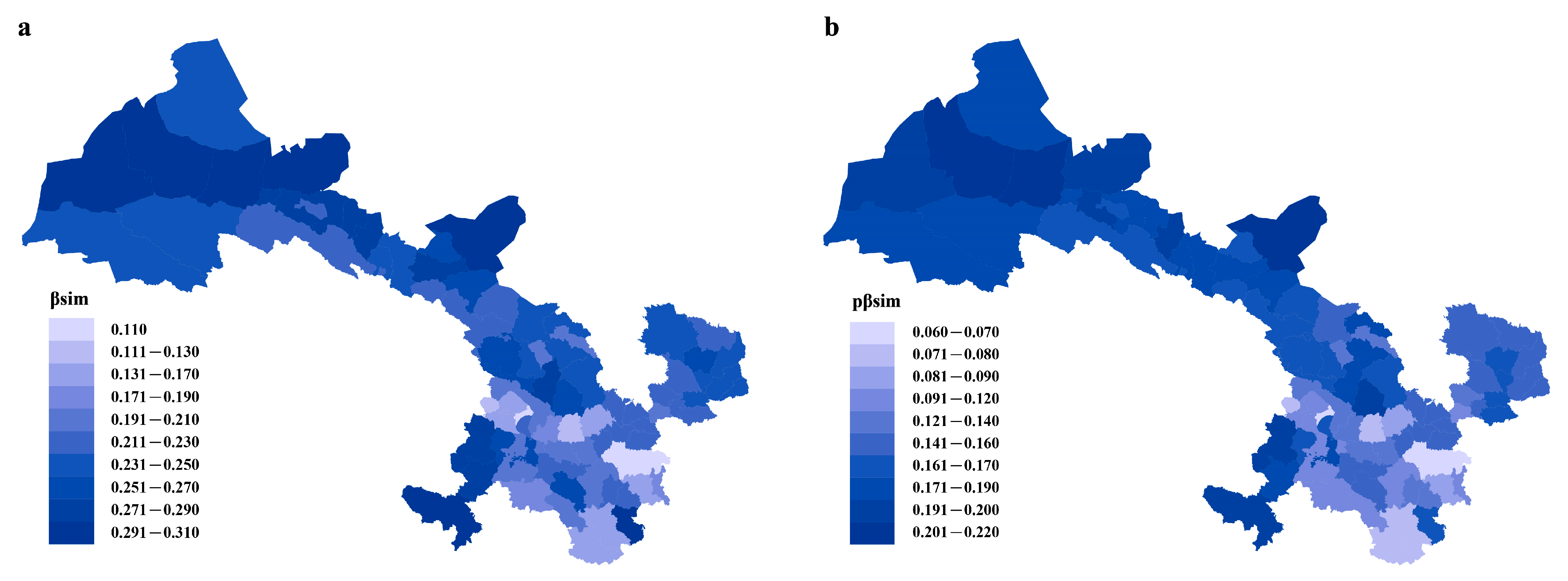{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clustering Algorithms | CCC (pβsim) | GD (pβsim) | CCC (βsim) | GD (βsim) |
|---|---|---|---|---|
| ward.D2 | 0.840 | 13,784.600 | 0.860 | 13,779.820 |
| ward.D | 0.814 | 213,347.500 | 0.830 | 212,874.300 |
| single | 0.856 | 298.473 | 0.838 | 335.790 |
| Complete | 0.859 | 266.409 | 0.869 | 230.959 |
| UPGMA | 0.872 | 38.090 | 0.876 | 37.130 |
| WPGMA | 0.867 | 44.907 | 0.873 | 38.218 |
| WPGMC | 0.813 | 243.142 | 0.830 | 409.498 |
| UPGMC | 0.813 | 213,347.500 | 0.830 | 212,874.300 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Tian, Q.; Chong, P.; Du, W.; Wei, J.; Huang, R. Phylogenetic Partitioning of Gansu Flora: Unveiling the Core Transitional Zone of Chinese Flora. Plants 2023, 12, 3060. https://doi.org/10.3390/plants12173060
Li Z, Tian Q, Chong P, Du W, Wei J, Huang R. Phylogenetic Partitioning of Gansu Flora: Unveiling the Core Transitional Zone of Chinese Flora. Plants. 2023; 12(17):3060. https://doi.org/10.3390/plants12173060
Chicago/Turabian StyleLi, Zizhen, Qing Tian, Peifang Chong, Weibo Du, Jia Wei, and Rong Huang. 2023. "Phylogenetic Partitioning of Gansu Flora: Unveiling the Core Transitional Zone of Chinese Flora" Plants 12, no. 17: 3060. https://doi.org/10.3390/plants12173060
APA StyleLi, Z., Tian, Q., Chong, P., Du, W., Wei, J., & Huang, R. (2023). Phylogenetic Partitioning of Gansu Flora: Unveiling the Core Transitional Zone of Chinese Flora. Plants, 12(17), 3060. https://doi.org/10.3390/plants12173060