

Exogenous Salicylic Acid Alleviates NO2 Damage by Maintaining Cell Stability and Physiological Metabolism in Bougainvillea × buttiana ‘Miss Manila’ Seedlings

Abstract
:1. Introduction
2. Results
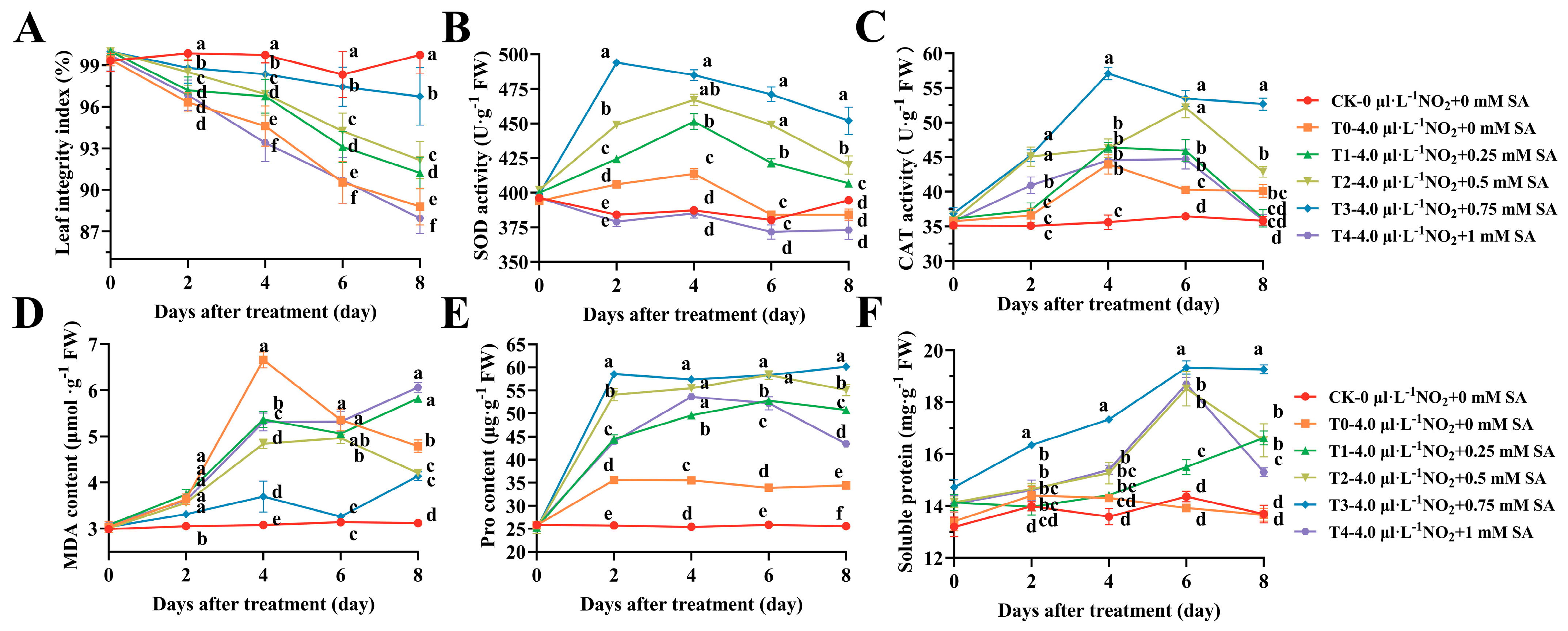
2.1. Changes in Morphology, Marker for Oxidative Stress, and Antioxidant Enzyme Activity
- (1).
- CK: Clean air + no SA;
- (2).
- T0: 4.0 μL·L−1 NO2 + 0 mM SA (pure water);
- (3).
- T1: 4.0 μL·L−1 NO2 + 0.25 mM SA;
- (4).
- T2: 4.0 μL·L−1 NO2 + 0.5 mM SA;
- (5).
- T3: 4.0 μL·L−1 NO2 + 0.75 mM SA;
- (6).
- T4: 4.0 μL·L−1 NO2 + 1.0 mM SA.
2.2. Changes in Photosynthetic Pigments and Gas Exchange Parameters
2.3. Changes in Nitrogen-Metabolizing Enzymes and Abundance of Different Forms of Nitrogen
2.4. Changes in Leaf Microstructure and Organelles
2.5. Analysis of Main Effects of Influencing Factors and Evaluation of Mitigation Effects at Different Concentrations
3. Discussion
3.1. Morphological Repair and Photosynthetic Physiological Alleviation of SA
3.2. Regulation of SA on the Microstructure and Nitrogen Metabolism Pathway
3.3. Systematic Evaluation and Analysis of the Mechanism of SA Regulating NO2 Injury
- (1).
- SA, as an important signaling molecule, interacts with other hormones, such as gibberellin, ethylene, and abscisic acid, to regulate the growth and response of B. × buttiana ‘Miss Manila’ under stress. The interactions between these hormones may include common signaling pathways, gene expression regulation, and metabolic regulators.
- (2).
- SA can activate the expression of defense response-related genes in plants under NO2 stress, produce antioxidant substances, and enhance cell wall stability.
- (3).
- SA can participate in the regulation of transcription factors, thereby altering the transcription activity of specific genes, regulating biochemical reactions and physiological processes.
- (4).
- SA can regulate stomatal opening and closing through the interaction of stomatal signaling pathways, thereby affecting plant water regulation and gas exchange.
4. Materials and Methods
4.1. Test Plants Materials
4.2. NO2 Fumigation Treatment and Sample Collection
4.3. Measurement Indicators and Methods
4.3.1. Morphological and In Vivo Osmotic Substances and Antioxidant Enzymes
4.3.2. Photosynthetic Pigments and Gas Exchange Parameters
4.3.3. Nitrogen Metabolism Enzymes and Different Forms of Nitrogen Abundance
4.3.4. Leaf Microstructure and Organelle Changes
4.3.5. Statistical Data Analysis and Processing
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Sheng, Q.Q.; Zhu, Z.L. Effects of Nitrogen Dioxide on Biochemical Responses in 41 Garden Plants. Plants 2019, 8, 45. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Higaki, A.; Nohno, M.; Kamada, M.; Okamura, Y.; Matsui, K.; Kitani, S.; Morikawa, H. Differential assimilation of nitrogen dioxide by 70 taxa of roadside trees at an urban pollution level. Chemosphere 2005, 61, 633–639. [Google Scholar] [CrossRef]
- Takahashi, M.; Furuhashi, T.; Ishikawa, N.; Horiguchi, G.; Sakamoto, A.; Tsukaya, H.; Morikawa, H. Nitrogen dioxide regulates organ growth by controlling cell proliferation and enlargement in Arabidopsis. New Phytol. 2014, 201, 1304–1305. [Google Scholar] [CrossRef]
- Cheng, X.J.; Frank, U.; Zhao, F.; Capella, J.R.; Winkler, J.B.; Schnitzler, J.P.; Ghirardo, A.; Bertic, M.; Estrella, N.; Durner, J.; et al. Plant growth traits and allergenic potential of Ambrosia artemisiifolia pollen as modified by temperature and NO2. Environ. Exp. Bot. 2023, 206, 105193. [Google Scholar] [CrossRef]
- Liu, X.F.; Fen, H.; Li, G.K.; Sang, N. Effects of nitrogen dioxide and its acid mist on reactive oxygen species production and antioxidant enzyme activity in Arabidopsis plants. J. Environ. Sci. 2015, 34, 93–99. [Google Scholar] [CrossRef]
- Zhao, S.; Liu, S.L.; Hou, X.Y.; Cheng, F.Y.; Wu, X.; Dong, S.K.; Beazley, R. Temporal dynamics of SO2 and NOX pollution and contributions of driving forces in urban areas in China. Environ. Pollut. 2018, 242 Pt. A, 239–248. [Google Scholar] [CrossRef]
- Shao, M.; Zhang, Y.H.; Zeng, L.M.; Tang, X.Y.; Zhang, J.; Zhong, L.J.; Wang, B.G. Ground-level ozone in the Pearl River Delta and the roles of VOC and NOx in its production. J. Environ. Manag. 2007, 90, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.M.; Wang, Y.F.; Wang, S.; Jiao, M.Y.; Huang, G.H.; Xia, B.C. Spatial-temporal heterogeneity of air pollution and its relationship with meteorological factors in the Pearl River Delta, China. Atmos. Environ. 2021, 254, 118415. [Google Scholar] [CrossRef]
- Khan, M.; Ali, S.; Al Azzawi, T.N.I.; Saqib, S.; Ullah, F.; Ayaz, A.; Zaman, W. The Key Roles of ROS and RNS as a Signaling Molecule in Plant–Microbe Interactions. Antioxidants 2023, 12, 268. [Google Scholar] [CrossRef] [PubMed]
- Campbell, S.A.; Vallano, D.M. Plant defences mediate interactions between herbivory and the direct foliar uptake of atmospheric reactive nitrogen. Nat. Commun. 2018, 9, 4743. [Google Scholar] [CrossRef]
- Okano, K.; Machida, T.; Totsuka, T. Differences in ability of NO2 absorption in various broad-leaved tree species. Environ. Pollut. 1989, 58, 1–17. [Google Scholar] [CrossRef]
- Sheng, Q.Q.; Zhu, Z.L. Photosynthetic Capacity, Stomatal Behavior and Chloroplast Ultrastructure in Leaves of the Endangered Plant Carpinus putoensis W.C. Cheng during Gaseous NO2 Exposure and after Recovery. Forests 2018, 9, 561. [Google Scholar] [CrossRef]
- Takahashi, M.; Nakagawa, M.; Sakamoto, A.; Morikawa, H. Atmospheric nitrogen dioxide gas is a plant vitalization signal to increase plant size and the contents of cell constituents. New Phytol. 2005, 168, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Papazian, S.; Blande, J.D. Dynamics of plant responses to combinations of air pollutants. Plant Biol. 2020, 22, 68–83. [Google Scholar] [CrossRef] [PubMed]
- Sumit, M.; Navdeep, G.; Kavita, D. NMR-based investigation of the altered metabolic response of Bougainvillea spectabilis leaves exposed to air pollution stress during the circadian cycle. Environ. Exp. Bot. 2019, 164, 58–70. [Google Scholar] [CrossRef]
- Sheng, Q.Q.; Song, M.; Zhu, Z.L.; Cao, F.L. Physiological and biochemical responses of two precious Carpinus species to high-concentration NO2 stress and their natural recovery. Sci. Rep. 2021, 11, 9500. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, W.W.; Che, Y.H.; Huang, D.; Wang, J.C.; Zhao, M.C.; Sun, G.Y. Atmospheric Nitrogen Dioxide Improves Photosynthesis in Mulberry Leaves via Effective Utilization of Excess Absorbed Light Energy. Forests 2019, 10, 312. [Google Scholar] [CrossRef]
- Sheng, Q.Q.; Zhou, C.Y.; Liang, Y.X.; Zhang, H.H.; Song, M.; Zhu, Z.L. Elevated NO2 induces leaf defensive mechanisms in Bougainvillea spectabilis seedlings. Ecotox. Environ. Safe 2022, 248, 114292. [Google Scholar] [CrossRef]
- Dempsey, A.D.; Amick, C.V.; Mary, C.W.; Daniel, F.K. Salicylic Acid Biosynthesis and Metabolism. Arab. Book 2011, 9, e0156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Y.; Zhu, Z.; Gao, J.; Zhou, X.; Zhu, S.; Wang, X.Y.; Kuai, B.K. The NPR1-WRKY46-WRKY6 signaling cascade mediates probenazole/salicylic acid-elicited leaf senescence in Arabidopsis thaliana. J. Integr. Plant Biol. 2020, 63, 924–936. [Google Scholar] [CrossRef] [PubMed]
- Rai, K.K.; Pandey, N.; Rai, S.P. Salicylic acid and nitric oxide signaling in plant heat stress. Physiol. Plantarum. 2020, 168, 241–255. [Google Scholar] [CrossRef]
- Jayakannan, M.; Bose, J.; Babourina, O. Salicylic acid in plant salinity stress signalling and tolerance. Plant Growth Regul. 2015, 76, 25–40. [Google Scholar] [CrossRef]
- Sharma, A.; Sidhu, G.P.S.; Araniti, F.; Bali, A.S.; Shahzad, B.; Tripathi, D.K.; Brestic, M.; Skalicky, M.; Landi, M.; Giampieri, F. The Role of Salicylic Acid in Plants Exposed to Heavy Metals. Molecules 2020, 25, 540. [Google Scholar] [CrossRef] [PubMed]
- Fateme, S.; Morteza, A.; Hossein, S.; Ali, K. Physiological and molecular mechanisms underlying salicylic acid-mitigated mercury toxicity in lemon balm (Melissa officinalis L.). Ecotox. Environ. Safe 2019, 183, 109542. [Google Scholar] [CrossRef]
- Dong, Q.L.; Duan, D.Y.; Zheng, W.Q.; Huang, D.; Wang, Q.; Li, X.R.; Ma, F.W. MdVQ37 overexpression reduces basal thermotolerance in transgenic apple by affecting transcription factor activity and salicylic acid homeostasis. Hortic. Res. 2021, 8, 220. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.T.; Li, T.T.; Zhang, L.Y.; Li, H.L.; Liu, S.K.; Yang, S.; Zou, N. Exogenous salicylic acid alleviates the accumulation of pesticides and mitigates pesticide-induced oxidative stress in cucumber plants (Cucumis sativus L.). Ecotox. Environ. Safe 2021, 208, 111654. [Google Scholar] [CrossRef]
- Khan, M.; Ali, S.; Manghwar, H.; Saqib, S.; Ullah, F.; Ayaz, A.; Zaman, W. Melatonin Function and Crosstalk with Other Phytohormones under Normal and Stressful Conditions. Genes 2022, 13, 1699. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, J.C.; Zhang, H.H.; Guo, D.D.; He, G.Q.; Sun, G.Y. A intermediate concentration of atmospheric nitrogen dioxide enhances PSII activity and inhibits PSI activity in expanded leaves of tobacco seedlings. Ecotox. Environ. Safe 2021, 209, 111844. [Google Scholar] [CrossRef]
- Ma, C.Y.; Feng, S.D.; Huang, L.L.; Wang, Y.; Li, N.; Xu, X.; Zhou, B.; Jia, K.; Xu, Q.; Li, R.G.; et al. Exogenous salicylic acid prevents nitrogen dioxide-induced oxidative injury and nitrate accumulation in Brassica campestris L. ssp. chinensis seedlings. J. Hortic. Sci. Biotech. 2010, 85, 241–247. [Google Scholar] [CrossRef]
- Cassaniti, C.; Leonardi, C.; Flowers, T.J. The effects of sodium chloride on ornamental shrubs. Sci. Hortic. 2009, 122, 586–593. [Google Scholar] [CrossRef]
- Nazir, H.; Salman, M.; Athar, M.; Farooq, U.; Wahab, A.; Akram, M. Citric acid functionalized bougainvillea spectabilis: A novel, sustainable, and cost-effective biosorbent for removal of heavy metal (Pb2+) from waste water. Water Air Soil Pollut. Int. J. Environ. Pollut. 2019, 230, 303. [Google Scholar] [CrossRef]
- Pal, A.; Kulshreshtha, K.; Ahmad, K.J.; Behl, H.M. Do leaf surface characters play a role in plant resistance to auto-exhaust pollution? Flora 2002, 197, 47–55. [Google Scholar] [CrossRef]
- Zhang, M.Z.; Liu, J.V.; Wang, W.J.; Bao, Y.Y. Responses of Bougainvillea spectabilis to elevated atmospheric CO2 under galaxolide (HHCB) pollution and the mechanisms of its rhizosphere metabolism. J. Soil Sediment. 2019, 19, 159–170. [Google Scholar] [CrossRef]
- Ullah, C.; Chen, Y.H.; Ortega, M.A.; Tsai, C.J. The diversity of salicylic acid biosynthesis and defense signaling in plants: Knowledge gaps and future opportunities. Curr. Opin. Plant Biol. 2023, 72, 102349. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Zhang, H.H.; Zhao, R.N.; Zhu, Z.L. Establishing an Efficient Regeneration System for Tissue Culture in Bougainvillea buttiana ‘Miss Manila’. Plants 2022, 11, 2372. [Google Scholar] [CrossRef] [PubMed]
- Ahanger, M.A.; Aziz, U.; Alsahli, A.A.; Alyemeni, M.N.; Ahmad, P. Influence of exogenous salicylic acid and nitric oxide on growth, photosynthesis, and ascorbate-glutathione cycle in salt stressed vigna angularis. Biomol. Ther. 2019, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Zaid, A.; Mohammad, F.; Wani, S.H.; Siddique, K.M.H. Salicylic acid enhances nickel stress tolerance by up-regulating antioxidant defense and glyoxalase systems in mustard plants. Ecotox. Environ. Safe 2019, 180, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Li, X.F.; Muhammad, R.; Song, B.Q.; Liang, X.L.; Liu, H.J. Exogenous salicylic acid alleviates fomesafen toxicity by improving photosynthetic characteristics and antioxidant defense system in sugar beet. Ecotox. Environ. Safe 2022, 238, 113587. [Google Scholar] [CrossRef]
- Hu, Y.L.; Yue, J.; Nie, J.Z.; Luo, D.J.; Cao, S.; Wang, C.J.; Chen, P. Salicylic acid alleviates the salt toxicity in kenaf by activating antioxidant system and regulating crucial pathways and genes. Ind. Crop. Prod. 2023, 199, 116691. [Google Scholar] [CrossRef]
- Moharekar, S.T.; Lokhande, S.D.; Hara, T.; Tanaka, R.; Tanaka, A.; Chavan, P.D. Effect of salicylic acid on chlorophyll and carotenoid contents of wheat and moong seedlings. Photosynthetica 2003, 41, 315–317. [Google Scholar] [CrossRef]
- Ababaf, M.; Omidi, H.; Bakhshandeh, A. Changes in antioxidant enzymes activities and alkaloid amount of Catharanthus roseus in response to plant growth regulators under drought condition. Ind. Crop. Prod. 2021, 167, 113505. [Google Scholar] [CrossRef]
- Ma, X.H.; Qian, R.J.; Zheng, J. Salicylic Acid Alleviates the Adverse Effects of Salt Stress on Dianthus superbus (Caryophyllaceae) by Activating Photosynthesis, Protecting Morphological Structure, and Enhancing the Antioxidant System. Front. Plant Sci. 2017, 8, 600. [Google Scholar] [CrossRef]
- Wassie, M.; Zhang, W.H.; Zhang, Q.; Ji, K.; Cao, L.W.; Chen, L. Exogenous salicylic acid ameliorates heat stress-induced damages and improves growth and photosynthetic efficiency in alfalfa (Medicago sativa L.). Ecotox. Environ. Safe 2020, 191, 110206. [Google Scholar] [CrossRef]
- Voznesenskaya, E.V.; Franceschi, V.R.; Kiirats, O.; Freitag, H.; Edwards, G.E. Kranz anatomy is not essential for terrestrial C4 plant photosynthesis. Nature 2001, 414, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Uzunova, A.N.; Popova, L.P. Effect of salicylic acid on leaf anatomy and chloroplast ultrastructure of barley plants. Photosynthetica 2000, 38, 243–250. [Google Scholar] [CrossRef]
- Chen, H.M.; Jin, J.J.; Hu, S.H.; Shen, L.Q.; Zhang, P.; Li, Z.H.; Fang, Z.G.; Liu, H.J. Metabolomics and proteomics reveal the toxicological mechanisms of florfenicol stress on wheat (Triticum aestivum L.) seedlings. J. Hazard. Mater. 2022, 443 Pt. B, 130264. [Google Scholar] [CrossRef]
- Jia, H.L.; Wang, X.H.; Wei, T.; Wang, M.; Liu, X.; Hua, L.; Ren, X.H.; Guo, J.K.; Li, J.S. Exogenous salicylic acid regulates cell wall polysaccharides synthesis and pectin methylation to reduce Cd accumulation of tomato. Ecotox. Environ. Safe 2021, 207, 111550. [Google Scholar] [CrossRef]
- Naeem, M.; Basit, A.; Ahmad, I.; Mohamed, H.I.; Wasila, H. Effect of Salicylic Acid and Salinity Stress on the Performance of Tomato Plants. Gesunde Pflanz. 2020, 72. [Google Scholar] [CrossRef]
- Sinha, A.; Gill, P.P.S.; Jawandha, S.K.; Kaur, P.; Grewal, S.K. Salicylic acid enriched beeswax coatings suppress fruit softening in pears by modulation of cell wall degrading enzymes under different storage conditions. Food Packag. Shelf. 2022, 32, 100821. [Google Scholar] [CrossRef]
- Cui, G.C.; Zhang, Y.; Zhang, W.J.; Lang, D.Y.; Zhang, X.J.; Li, Z.X.; Zhang, X.H. Response of Carbon and Nitrogen Metabolism and Secondary Metabolites to Drought Stress and Salt Stress in Plants. J. Plant Biol. 2019, 62, 387–399. [Google Scholar] [CrossRef]
- Hu, Y.B.; Bellaloui, N.; Tigabu, M.; Wang, J.H.; Diao, J.; Wang, K.; Yang, R.; Sun, G.Y. Gaseous NO2 effects on stomatal behavior, photosynthesis and respiration of hybrid poplar leaves. Acta Physiol. Plant. 2015, 37, 1–8. [Google Scholar] [CrossRef]
- Chen, Z.M.; Chen, Y.X.; Du, G.J.; Wu, X.L.; Li, F. Effects of 60-day NO2 fumigation on growth, oxidative stress and antioxidative response in Cinnamomum camphora seedlings. J. Zhejiang Univ. Sci. B 2010, 11, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Hayat, Q.; Hayat, S.; Alyemeni, M.N.; Ahmad, A. Salicylic acid mediated changes in growth, photosynthesis, nitrogen metabolism and antioxidant defense system in Cicer arietinum L. Plant Soil Environ. 2022, 32, 417–423. [Google Scholar] [CrossRef]
- Gaffney, T.; Friedrich, L.; Vernooij, B.; Negrotto, D.; Nye, G.; Uknes, S.; Ward, E.; Kessmann, H.; Ryals, J. Requirement of salicylic acid for the induction of systemic acquired resistance. Science 1993, 261, 754–756. [Google Scholar] [CrossRef]
- Li, W.; Shi, Y.F.; Zhu, D.D.; Wang, W.Q.; Liu, H.W.; Li, J.Y.; Shi, N.N.; Ma, L.; Fu, S.L. Fine root biomass and morphology in a temperate forest are influenced more by the nitrogen treatment approach than the rate. Ecol. Indic. 2021, 130, 108031. [Google Scholar] [CrossRef]
- Xu, Q.; Zhou, B.; Ma, C.Y.; Xu, X.; Xu, J.; Jiang, Y.G.; Liu, C.; Li, G.Z.; Herbert, S.J.; Hao, L. Salicylic acid-altering arabidopsis mutants response to NO(2) exposure. B. Environ. Contam. Tox. 2010, 84, 106–111. [Google Scholar] [CrossRef]
- Daqiu, Z.; Ziwen, F.; Yuhan, T.; Tao, J. Graphene oxide as an effective soil water retention agent can confer drought stress tolerance to Paeonia ostii without toxicity. Environ. Sci. Technol. 2020, 54, 8269–8279. [Google Scholar] [CrossRef]
- Pietryczuk, A.; Czerpak, R. Effect of traumatic acid on antioxidant activity in Chlorella vulgaris (Chlorophyceae). Plant Growth Regul. 2011, 65, 279–286. [Google Scholar] [CrossRef]
- Zhou, Q. Plant Physiology; China Agricultural University Press: Beijing, China, 2000; pp. 163–166. [Google Scholar]
- Gao, J.F. Experimental Guidance of Plant Physiology; Higher Education Press: Beijing, China, 2003; pp. 61–65. [Google Scholar]
- Oyahama, T.; Ito, M.; Kobayashi, K.; So, A. Analytical procedures of N, P, K contents and manure materials using H2SO4–H2O2 Kjeldahl digestion method. Bull. Fac. Agric. Niigata Univ. 1991, 43, 111–120. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | p Value between Treatments | |||||
|---|---|---|---|---|---|---|
| CK | T0 | T1 | T2 | T3 | T4 | |
| MDA | 0.002 ** | 0.021 | 0.001 ** | 0.007 ** | 0.001 ** | 0.001 ** |
| SOD | 0.286 | 0.035 | 0.614 | 0.458 | 1.000 | 0.003 ** |
| CAT | 0.085 | 0.018 | 0.411 | 0.148 | 0.016 | 0.534 |
| Pro | 0.772 | 0.642 | 0.001 ** | 0.005 ** | 0.003 | 0.160 |
| Chla | 0.001 ** | 0.000 ** | 0.002 ** | 0.001 ** | 0.001 ** | 0.001 ** |
| Chlb | 0.173 | 0.005 | 0.001 ** | 0.969 | 0.001 ** | 0.001 ** |
| Chl | 0.001 ** | 0.001 ** | 0.001 ** | 0.001 ** | 0.001 ** | 0.001 ** |
| Car | 0.013 | 0.001 ** | 0.011 | 0.001 ** | 0.001 ** | 0.001 ** |
| Pn | 0.001 ** | 0.001 ** | 0.001 ** | 0.001 ** | 0.001 ** | 0.001 ** |
| Tr | 0.001 ** | 0.001 ** | 0.001 ** | 0.002 | 0.001 ** | 0.001 ** |
| SC | 0.001 ** | 0.001 ** | 0.001 ** | 0.001 ** | 0.001 ** | 0.001 ** |
| Ci | 0.021 | 0.001 ** | 0.013 | 0.013 | 0.001 ** | 0.011 |
| SL | 0.005 ** | 0.001 ** | 0.007 ** | 0.001 ** | 0.641 | 0.001 ** |
| SW | 0.259 | 0.015 | 0.233 | 0.194 | 0.482 | 0.001 ** |
| Sa | 0.114 | 0.001 ** | 0.063 | 0.003 ** | 0.969 | 0.001 ** |
| Stp | 0.010 | 0.001 ** | 0.180 | 0.002 ** | 0.728 | 0.787 |
| SO | 0.938 | 0.006 ** | 0.001 ** | 0.001 ** | 0.187 | 0.001 ** |
| SD | 0.969 | 0.007 ** | 0.532 | 0.029 | 0.187 | 0.335 |
| NR | 0.892 | 0.250 | 0.268 | 0.160 | 0.586 | 0.035 |
| NiR | 0.786 | 0.011 | 0.005 ** | 0.001 ** | 0.001 ** | 0.021 |
| GS | 0.016 | 0.095 | 0.001 ** | 0.001 ** | 0.001 ** | 0.001 ** |
| GOGAT | 0.172 | 0.728 | 0.008 ** | 0.021 | 0.005 ** | 0.021 |
| GDH | 0.420 | 0.022 | 0.001 ** | 0.001 ** | 0.001 ** | 0.001 ** |
| SP | 0.187 | 0.787 | 0.001 ** | 0.001 ** | 0.001 ** | 0.001 ** |
| TN | 0.367 | 0.124 | 0.001 ** | 0.002 | 0.001 ** | 0.001 ** |
| NO3 | 0.642 | 0.021 | 0.001 ** | 0.728 | 0.001 ** | 0.001 ** |
| NH4 | 0.268 | 0.021 | 0.021 | 0.005 | 0.013 | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, Y.; Li, D.; Sheng, Q.; Zhu, Z. Exogenous Salicylic Acid Alleviates NO2 Damage by Maintaining Cell Stability and Physiological Metabolism in Bougainvillea × buttiana ‘Miss Manila’ Seedlings. Plants 2023, 12, 3283. https://doi.org/10.3390/plants12183283
Liang Y, Li D, Sheng Q, Zhu Z. Exogenous Salicylic Acid Alleviates NO2 Damage by Maintaining Cell Stability and Physiological Metabolism in Bougainvillea × buttiana ‘Miss Manila’ Seedlings. Plants. 2023; 12(18):3283. https://doi.org/10.3390/plants12183283
Chicago/Turabian StyleLiang, Yuxiang, Dalu Li, Qianqian Sheng, and Zunling Zhu. 2023. "Exogenous Salicylic Acid Alleviates NO2 Damage by Maintaining Cell Stability and Physiological Metabolism in Bougainvillea × buttiana ‘Miss Manila’ Seedlings" Plants 12, no. 18: 3283. https://doi.org/10.3390/plants12183283
APA StyleLiang, Y., Li, D., Sheng, Q., & Zhu, Z. (2023). Exogenous Salicylic Acid Alleviates NO2 Damage by Maintaining Cell Stability and Physiological Metabolism in Bougainvillea × buttiana ‘Miss Manila’ Seedlings. Plants, 12(18), 3283. https://doi.org/10.3390/plants12183283