

Positive Effect of Green Photo-Selective Filter on Graft Union Formation in Tomatoes

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Oxidative Stress
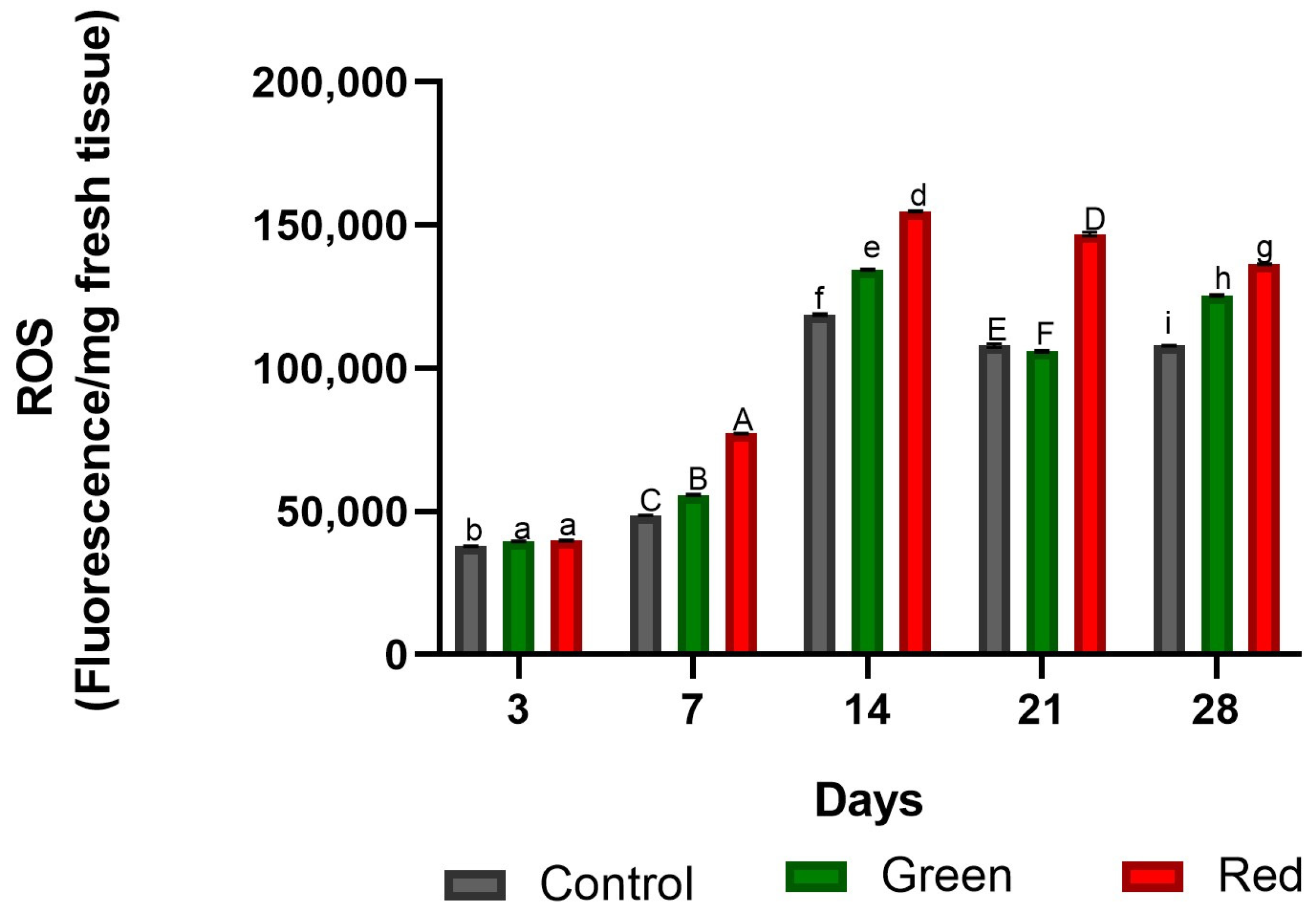
2.1.1. Determination of Reactive Oxygen Species
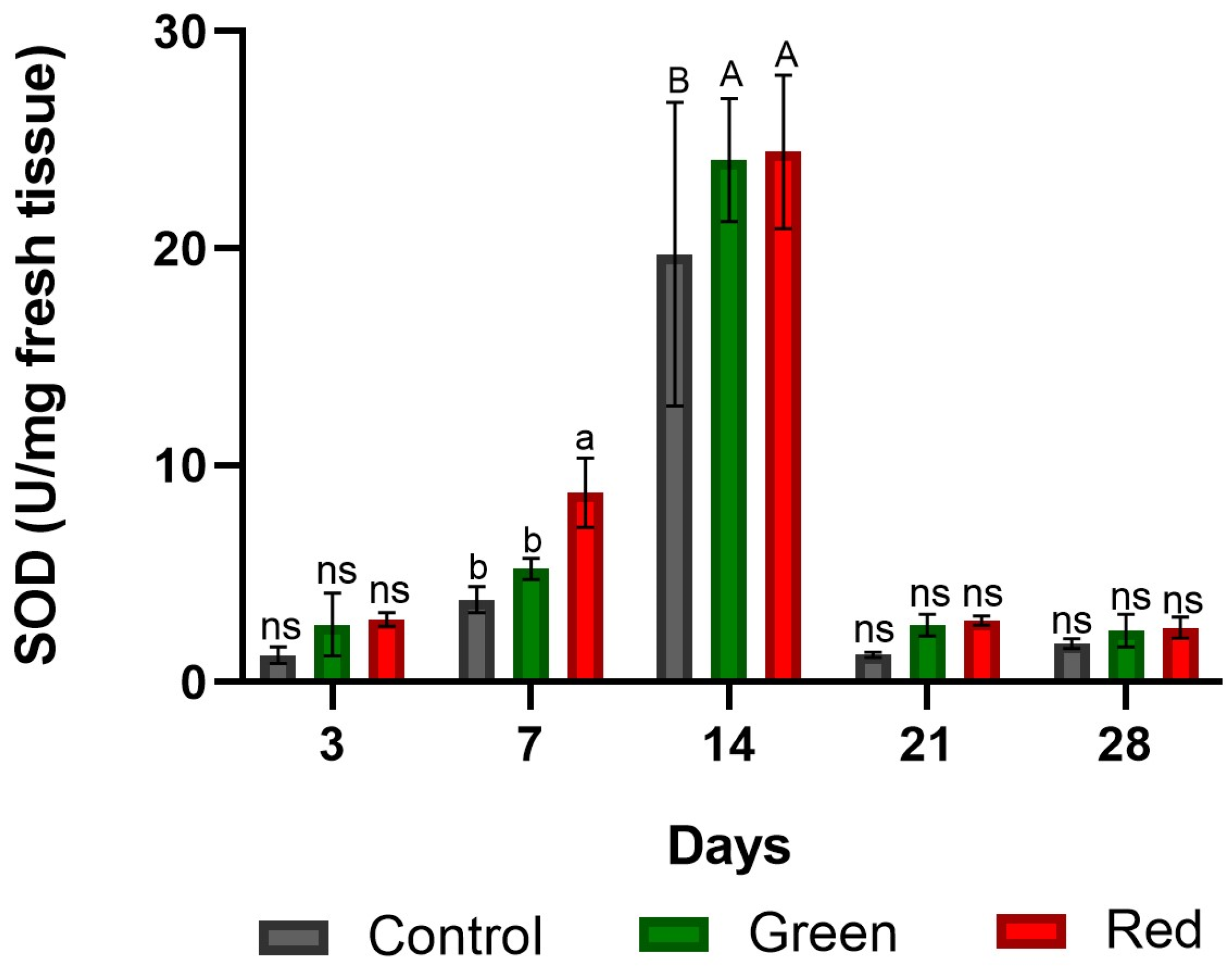
2.1.2. Superoxide Dismutase Enzyme Activity
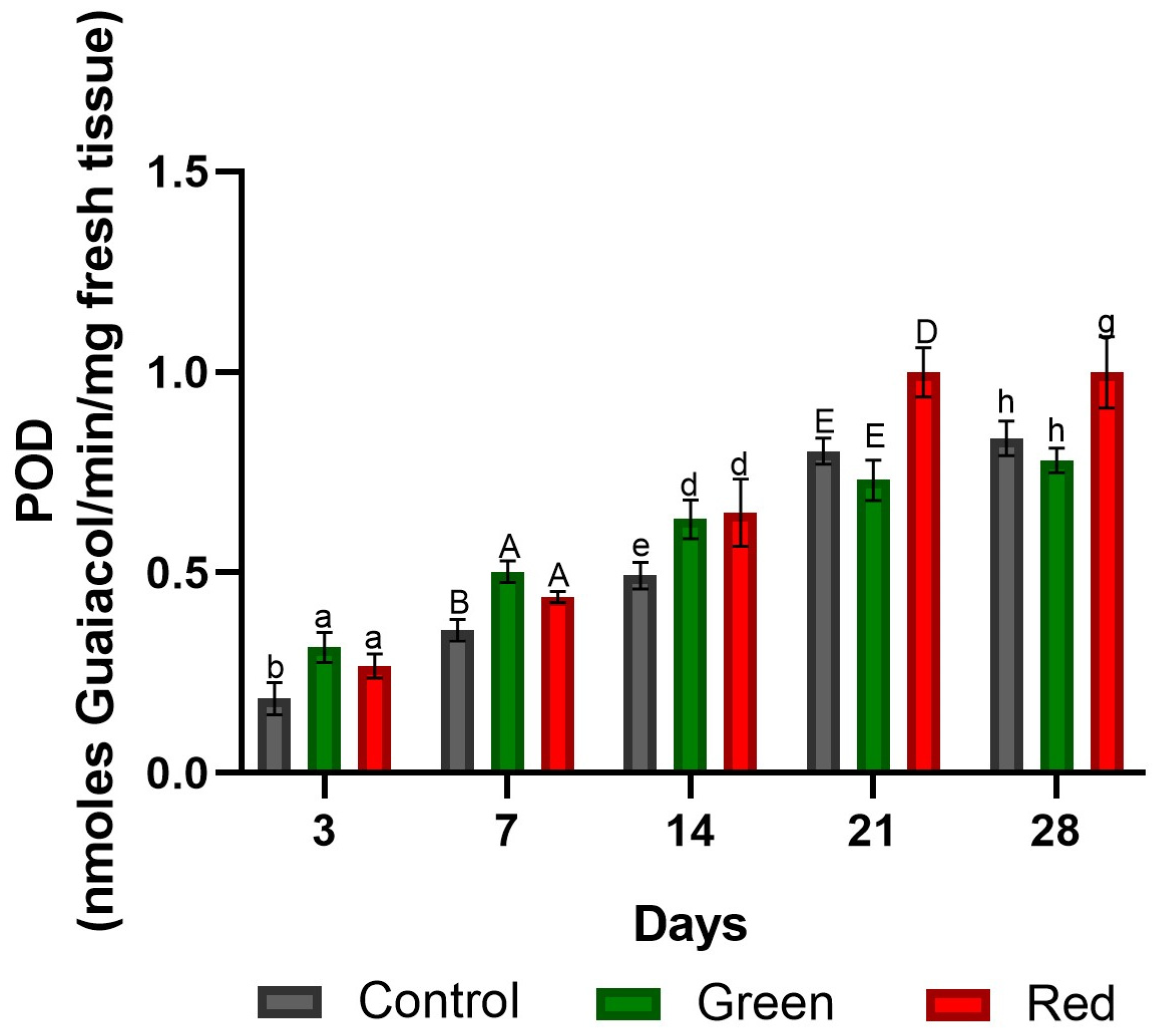
2.1.3. Peroxidase Enzyme Activity
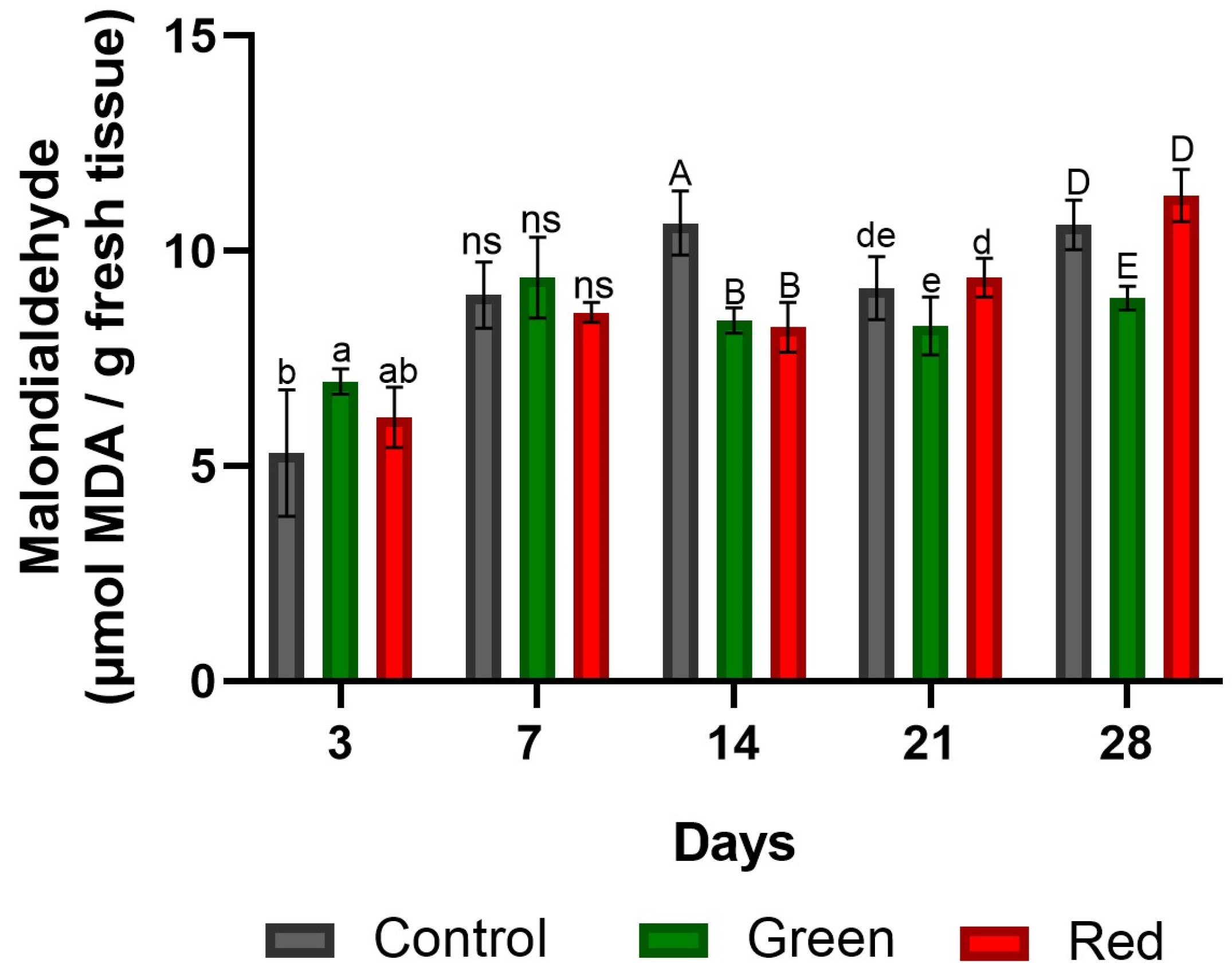
2.1.4. Determination of Malondialdehyde
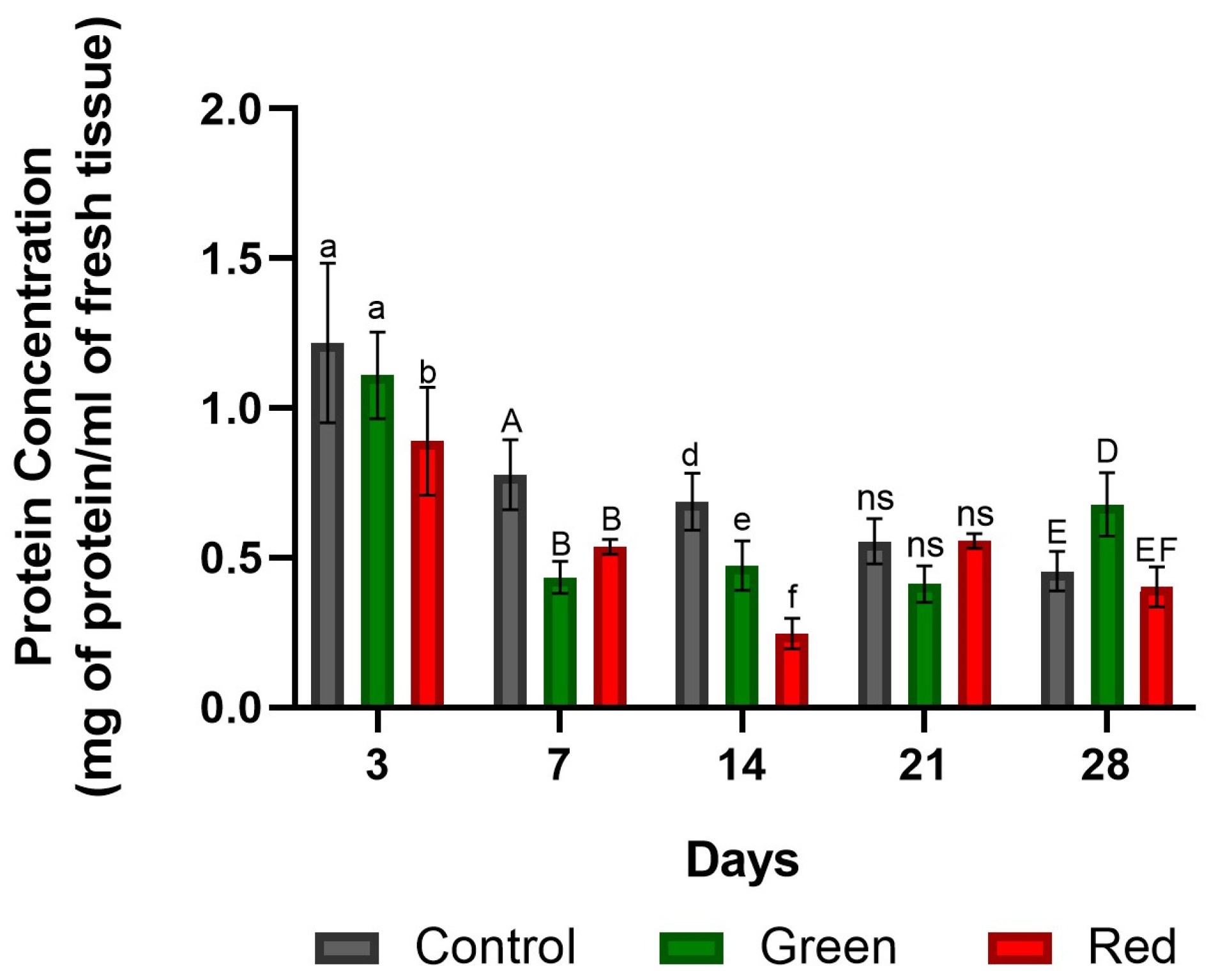
2.1.5. Protein Concentration of SOD/POD-Enriched Extracts
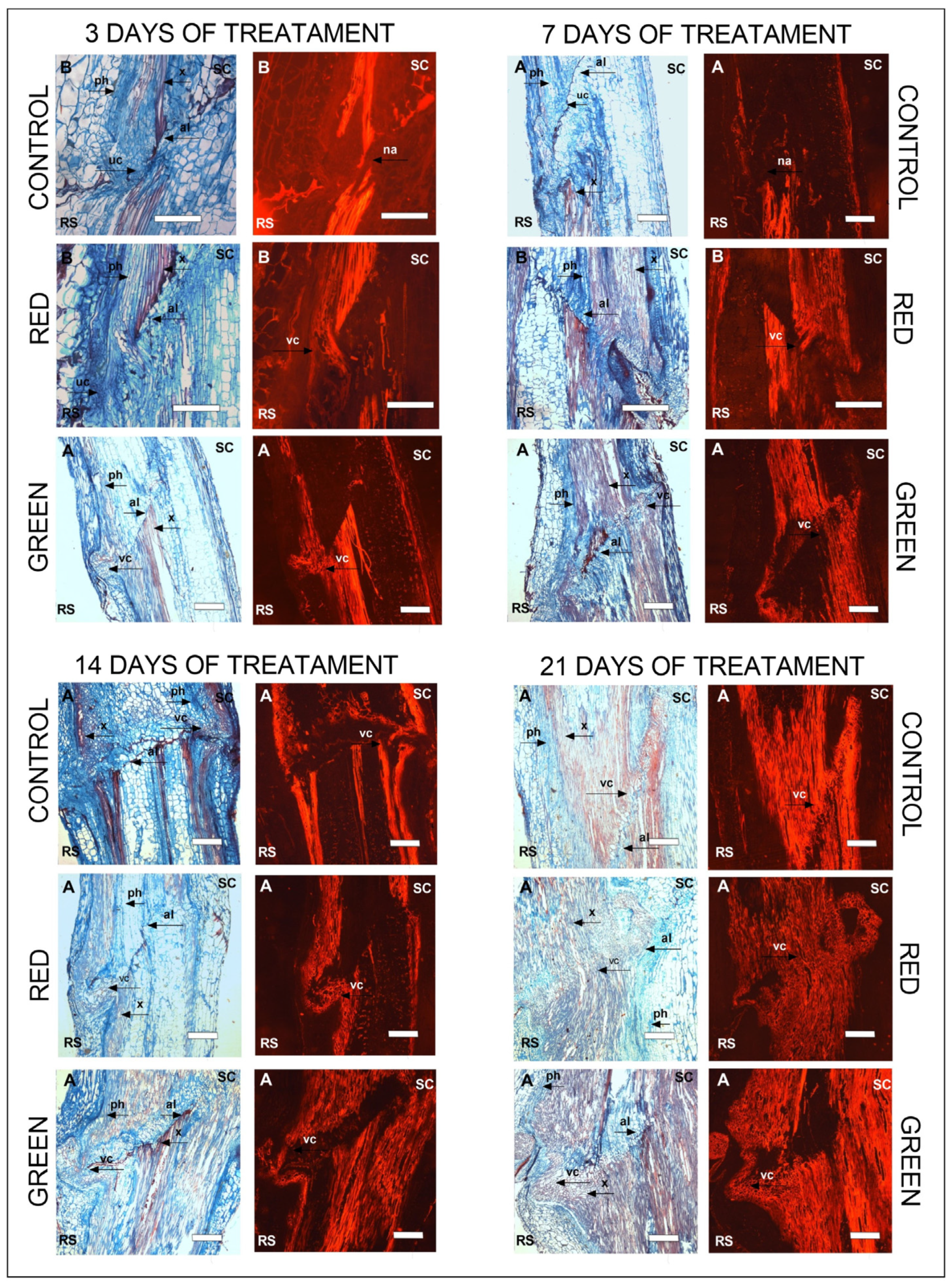
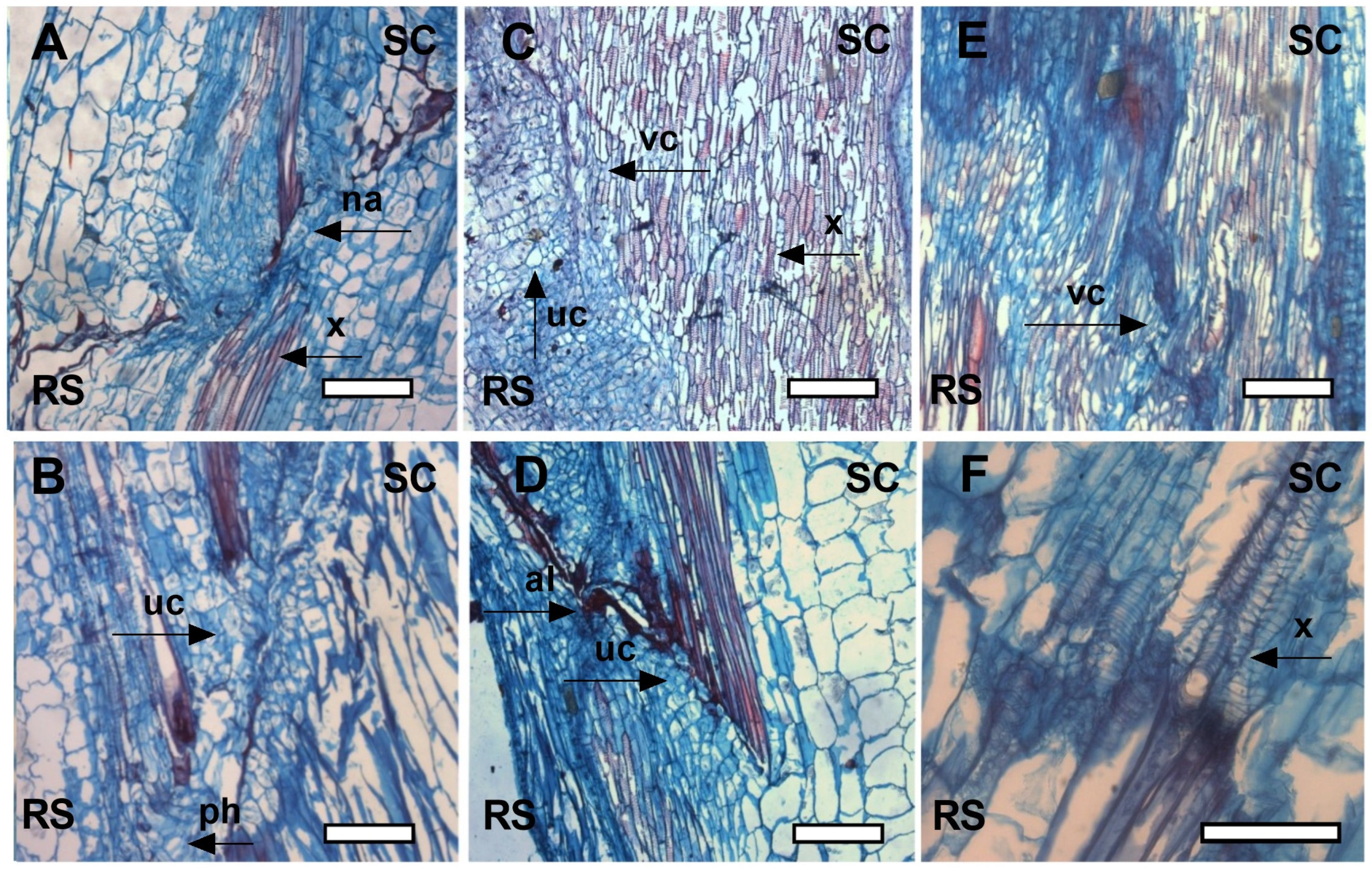
2.2. Anatomical Development
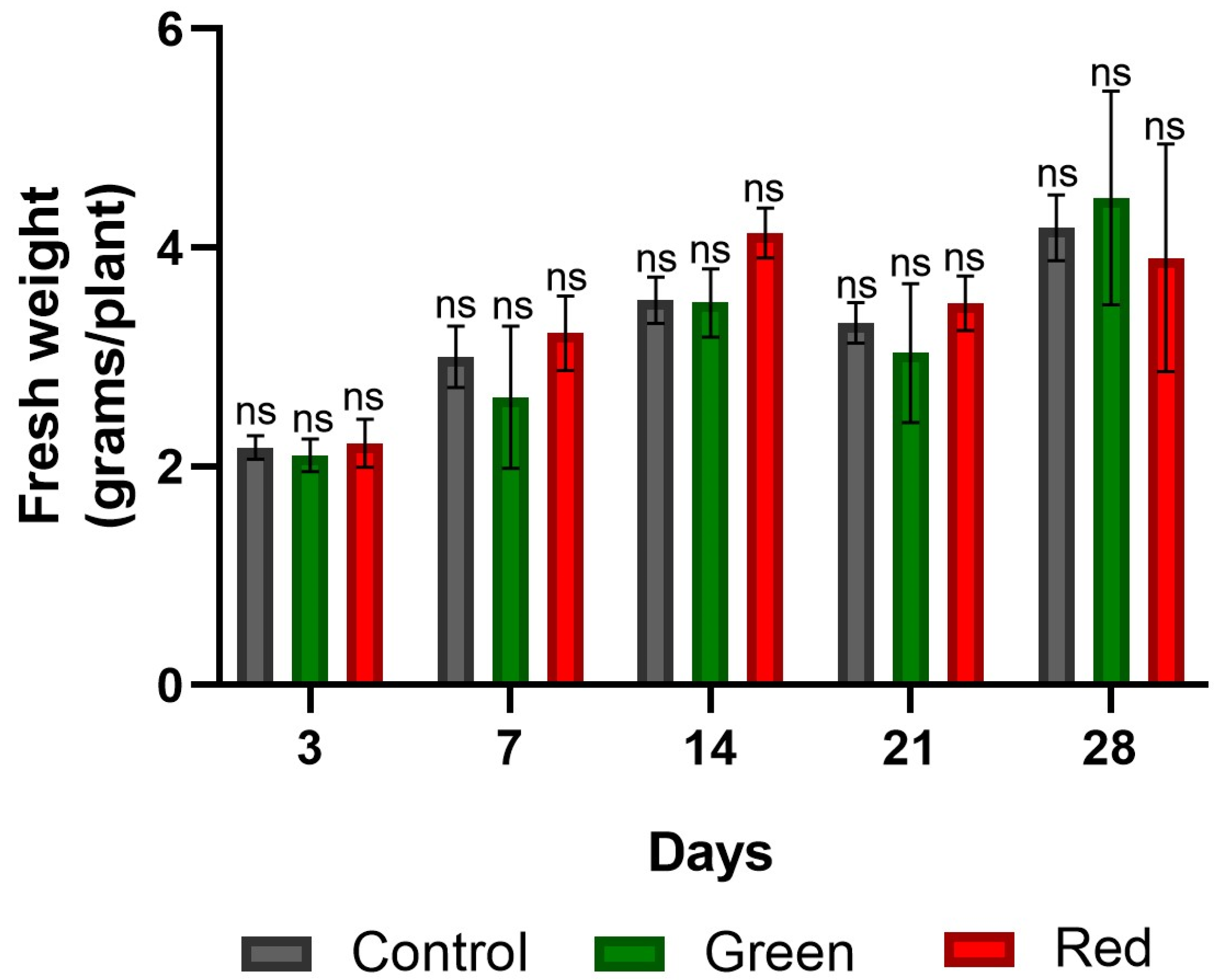
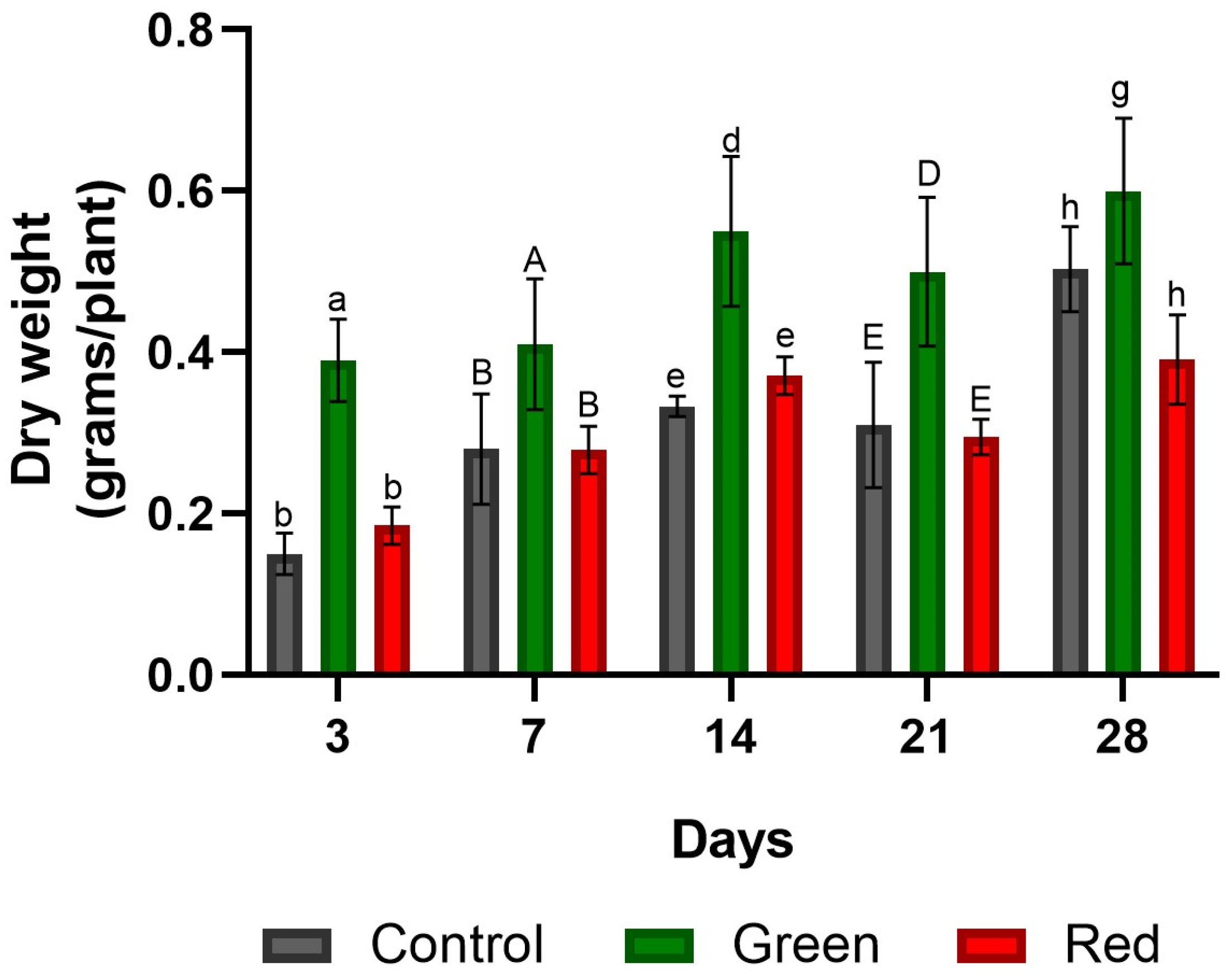
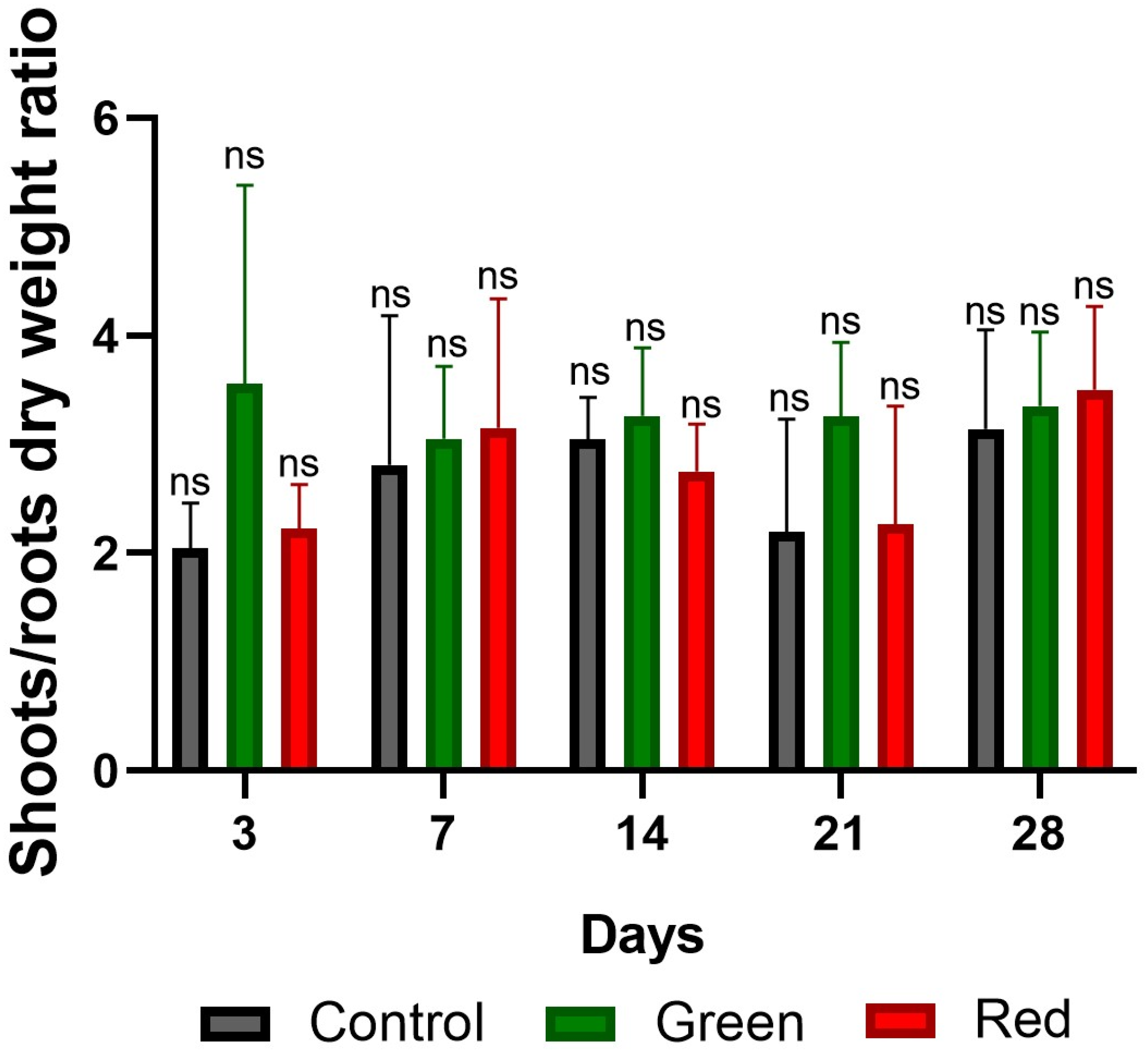
2.3. Fresh and Dry Plant Weight
3. Discussion
4. Materials and Methods
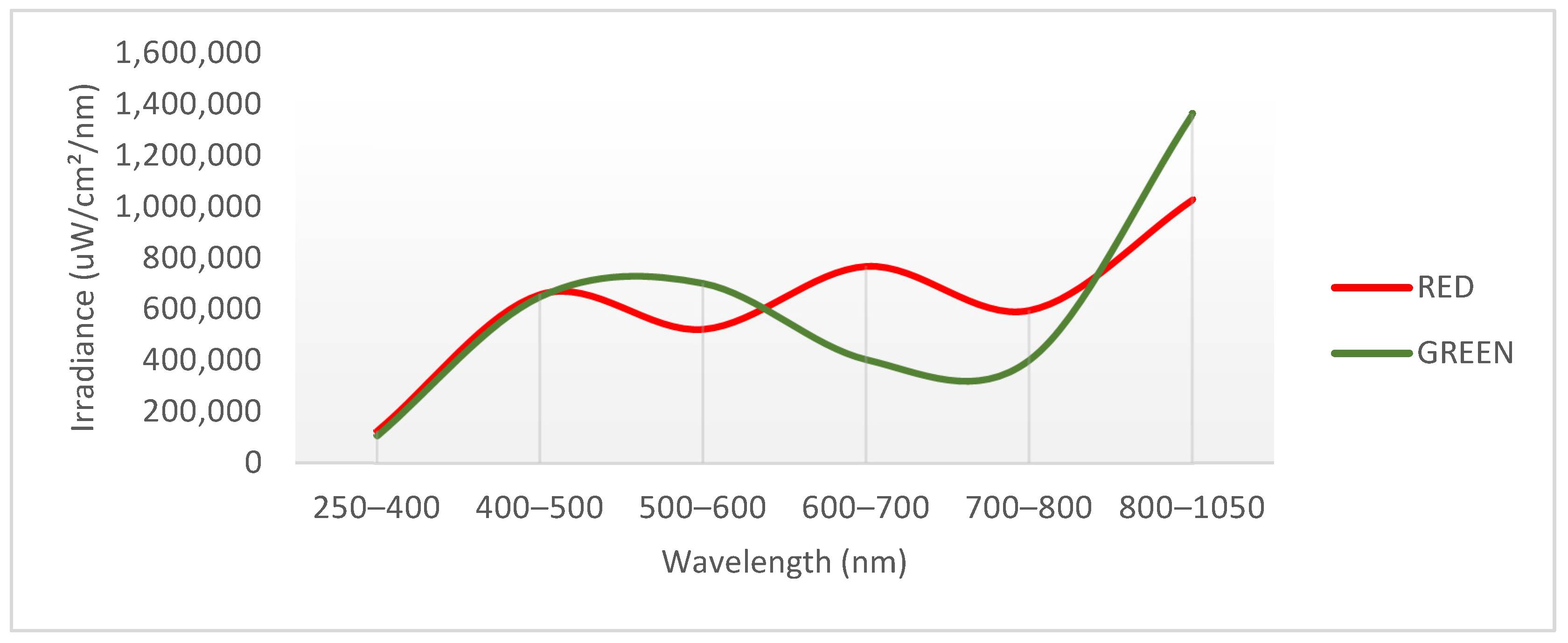
4.1. Growth Conditions
4.2. Reactive Oxygen Species Activity Measurement
4.3. Antioxidant Enzymes
4.3.1. Extract
4.3.2. Superoxide Dismutase
4.3.3. Peroxidase
4.4. MDA
4.5. Anatomical Development
4.6. Fresh and Dry Weight
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. FAOSTAT: Crop Production Data. 2019. Available online: http://www.fao.org/faostat/es/#data/QC/visualize (accessed on 21 March 2023).
- Kumar, P.; Rouphael, Y.; Cardarelli, M.; Colla, G. Vegetable grafting as a tool to improve drought resistance and water use efficiency Front. Plant Sci. 2017, 8, 1130–1139. [Google Scholar] [CrossRef]
- Singh, H.; Kumar, P.; Chaudhari, S.; Edelstein, M. Tomato grafting: A global perspective. HortScience 2017, 52, 1328–1336. [Google Scholar] [CrossRef]
- Leonardi, C.; Giuffrida, F. Variation of plant growth and macronutrient uptake in grafted tomatoes and eggplants on three different rootstocks. Eur. J. Hortic. Sci. 2006, 71, 97–101. [Google Scholar]
- Khah, E.M.; Kakava, E.; Mavromatis, A.; Chachalis, D.; Goulas, C. Effect of grafting on growth and yield of tomato (Lycopersicon esculentum Mill.) in greenhouse and open-field. J. Appl. Hortic. 2006, 8, 3–7. [Google Scholar] [CrossRef]
- Krumbein, A.; Schwarz, D. Grafting: A possibility to enhance health-promoting and flavor compounds in tomato fruits of shaded plants. Sci. Hortic. 2013, 149, 97–107. [Google Scholar] [CrossRef]
- Naik, S.A.; Hongal, S.; Harshavardhan, M.; Chandan, K.; Kumar, A.J.; Ashok; Kyriacou, M.C.; Rouphael, Y.; Kumar, P. Productive characteristics and fruit quality traits of cherry tomato hybrids as modulated by grafting on different Solanum spp. rootstocks under Ralstonia solanacearum infested greenhouse soil. Agronomy 2021, 11, 1311. [Google Scholar] [CrossRef]
- Schwarz, D.; Rouphael, Y.; Colla, G.; Venema, J. Grafting as a tool to improve tolerance of vegetables to abiotic stresses: Thermal stress, water stress and organic pollutants. Sci. Hortic. 2010, 127, 162–171. [Google Scholar] [CrossRef]
- Aidoo, M.; Sherman, T.; Ephrath, J.E.; Fait, A.; Rachmilevitch, S.; Lazarovitch, N. Grafting as a method to increase the tolerance response of bell pepper to extreme temperatures. Vadose Zone J. 2018, 17, 1–8. [Google Scholar] [CrossRef]
- Hartmann, H.; Kester, D.; Davies, F.; Geneve, R. Plant Propagation. Principles and Practices, 8th ed.; Prentice-Hall: London, UK, 2018; pp. 433–475. ISBN 0-13-679235-679239. [Google Scholar]
- Lee, J.; Oda, M. Grafting of herbaceous vegetable and ornamental crops. In Horticultural Reviews; Janick, J., Ed.; John Wiley & Sons: Oxford, UK, 2010; Volume 28, pp. 61–124. [Google Scholar] [CrossRef]
- Tambussi, E. Fotosíntesis, Fotoprotección, Productividad y Estrés Abiótico: Algunos Casos de Estudio. Doctoral Thesis, Universidad de Barcelona, Barcelona, España, 2004. [Google Scholar]
- Soares, N.; Wojtkowska, J.; Jackson, P. A proteomic analysis of the wound response in Medicago leaves reveals the early activation of a ROS-sensitive signal pathway. J. Proteom. 2011, 74, 1411–1420. [Google Scholar] [CrossRef]
- Muneer, S.; Ko, C.; Yang, Z.; Chen, Y.; Jeong, B. Physiological and proteomic investigations to study the response of tomato graft unions under temperature stress. PLoS ONE 2016, 11, e0157439. [Google Scholar] [CrossRef]
- Meloni, D.; Gulotta, M.; Oliva, M. El estrés salino incrementa la actividad de enzimas antioxidantes y la concentración de polifenoles en Vinal (Prosopis ruscifolia). Quebracho-Rev. Cienc. For. 2008, 15, 27–31. [Google Scholar]
- Fernandez, N. Graft Union Formation in Tomato Plants: Peroxidase and Catalase Involvement. Ann. Bot. 2004, 91, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Boudet, A.; Kajita, S.; Grima-Pettenati, J.; Goffner, D. Lignins and lignocellulosics: A better control of synthesis for new and improved uses. Plant Sci. 2003, 8, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Raihan, M.R.H.; Masud, A.A.C.; Rahman, K.; Nowroz, F.; Rahman, M.; Nahar, K.; Fujita, M. Regulation of Reactive Oxygen Species and Antioxidant Defense in Plants under Salinity. Int. J. Mol. Sci. 2021, 22, 9326. [Google Scholar] [CrossRef] [PubMed]
- Adil, M.; Haider Abbasi, B.; ul Haq, I. Red light controlled callus morphogenetic patterns and secondary metabolites production in Withania somnifera L. Biotech. Rep. 2019, 24, e00380. [Google Scholar] [CrossRef]
- Johnson, S.; Miles, C. Effect of healing chamber design on the survival of grafted eggplant, tomatoand watermelon. HortTechnology 2011, 21, 752–758. [Google Scholar] [CrossRef]
- Verma, M.; Bhat, P.; Bhartiya, S.; Venkatesh, K. A steady-state modeling approach to validate an in vivo mechanism of the GAL regulatory network in Saccharomyces cerevisiae. Eur. J. Biochem. 2004, 271, 4064–4074. [Google Scholar] [CrossRef]
- Frey, C.; Álvarez, R.; Encina, A.; Acebes, J. Tomato graft union failure is associated with alterations in tissue development and the onset of cell wall defense responses. Agronomy 2021, 11, 1197. [Google Scholar] [CrossRef]
- Frey, C.; Acebes, J.L.; Encina, A.; Álvarez, R. Histological changes associated with the graft union development in tomato. Plants 2020, 9, 1479. [Google Scholar] [CrossRef]
- Martínez, M.; Alcaraz, C.; Muries, B.; Mota, C.; Carvajal, M. Physiological aspects of rootstock–scion interactions. Sci. Hortic. 2010, 127, 112–118. [Google Scholar] [CrossRef]
- Vu, N.-T.; Xu, Z.-H.; Kim, Y.-S.; Kang, H.-M.; Kim, I.-S. Effect of nursery environmental condition and different cultivars on survival rate of grafted tomato seedling. Acta Hortic. 2014, 1037, 765–770. [Google Scholar] [CrossRef]
- Zeist, A.; de Resende, J.; Zanin, D.; Silva, A.; Perrud, A.; Bueno, G.; Arantes, J.; de Lima, D. Effect of acclimation environments, grafting methods and rootstock RVTC-66 on the seedling development and production of tomato. Sci. Hortic. 2020, 271, 109496. [Google Scholar] [CrossRef]
- Wei, H.; Muneer, S.; Manivannan, A.; Liu, Y.; Park, J.; Jeong, B. Slight vapor deficit accelerates graft union healing of tomato plug seedling. Acta Physiol. Plant 2018, 40, 147. [Google Scholar] [CrossRef]
- Nguy-Robertson, A.; Suyker, A.; Xiao, X. Modeling gross primary production of maize and soybean croplands using light quality, temperature, water stress, and phenology. Agric. For. Meteorol. 2015, 213, 160–172. [Google Scholar] [CrossRef]
- Casierra, F.; Matallana, Y.; Zapata, E. Growth of bell pepper plants (Capsicum annuum) affected by coloured covers. Ges. Pflanzen 2014, 6, 149–155. [Google Scholar] [CrossRef]
- Devlin, P.; Christie, J.; Terry, M. Many hands make light work. J. Exp. Bot. 2007, 58, 3071–3077. [Google Scholar] [CrossRef]
- Kaiser, E.; Weerheim, K.; Schipper, R.; Dieleman, J. Partial replacement of red and blue by green light increases biomass and yield in tomato. Sci. Hortic. 2019, 249, 271–279. [Google Scholar] [CrossRef]
- Bian, Z.; Zhang, X.; Wang, Y.; Lu, C. Improving drought tolerance by altering the photosynthetic rate and stomatal aperture via green light in tomato (Solanum lycopersicum L.) seedlings under drought conditions. Environ. Exp. Bot. 2019, 167, 103844. [Google Scholar] [CrossRef]
- Dieleman, J.; de Visser, P.; Meinen, E.; Grit, J.; Dueck, T. Integrating morphological and physiological responses of tomato plants to light quality to the crop level by 3D modeling. Plant Sci. 2019, 10, 839. [Google Scholar] [CrossRef]
- Schenkels, L.; Saeys, W.; Lauwers, A.; De Proft, M. Green light induces shade avoidance to alter plant morphology and increases biomass production in Ocimum basilicum L. Sci. Hortic. 2019, 261, 109002. [Google Scholar] [CrossRef]
- Singla, A.; Sharma, R.; Chhabra, R. Effect of varying shade intensities of green net on growth and stomatal attributes of different Ocimum species. Proc. Natl. Acad. Sci. USA 2021, 91, 865–878. [Google Scholar] [CrossRef]
- Ayala, F.; Sánchez, R.; Partida, L.; Yáñez, M.; Ruiz, F.; Velázquez, T.; Valenzuela, M.; Parra, J. Bell pepper production under colored shade nets. Rev. Fitotec. Mex. 2015, 38, 93–99. [Google Scholar]
- Thwe, A.; Kasemsap, P.; Vercambre, G.; Phattaralerphong, J.; Gautier, H. Impact of red and blue nets on physiological and morphological traits, fruit yield and quality of tomato (Solanum lycopersicum mill.). Sci. Hortic. 2020, 264, 09185. [Google Scholar] [CrossRef]
- Yousef, A.; Ali, M.; Rizwan, H. Light quality and quantity affect graft union formation of tomato plants. Sci. Rep. 2021, 11, 9870. [Google Scholar] [CrossRef]
- Tan, Y.; Duan, Y.; Chi, Q.; Wang, R.; Yin, Y.; Cui, D.; Li, S. The role of reactive oxygen species in plant response to radiation. Int. J. Mol. Sci. 2023, 24, 3346. [Google Scholar] [CrossRef]
- Milkovic, L.; Cipak, A.; Cindric, M.; Mouthuy, P.; Zarkovic, N. Short Overview of ROS as Cell Func-tion Regulators and Their Implications in Therapy Concepts. Cells 2019, 8, 793. [Google Scholar] [CrossRef]
- Manivannan, A.; Soundararajan, P.; Park, Y.; Wei, H.; Kim, S.; Jeong, B. Blue and red light-emitting diodes improve the growth and physiology of in vitro-grown carnations “Green Beauty” and “Purple Beauty”. Hortic. Environ. Biotechnol. 2017, 58, 12–20. [Google Scholar] [CrossRef]
- Xu, Y.; Liang, Y.; Yang, M. Effects of composite led light on root growth and antioxidant capacity of Cunninghamia lanceolata tissue culture seedlings cunninghamia lanceolata. Sci. Rep. 2019, 9, 9766. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Li, Y.; Li, S.; Wu, G.; Niu, X.; Shen, A. Green light promotes healing and root regeneration in double-root-cutting grafted tomato seedlings. Sci. Hortic. 2021, 289, 110503. [Google Scholar] [CrossRef]
- Fan, J.; Yang, R.; Li, X.; Zhao, W.; Zhao, F.; Wang, S. The processes of graft union formation in tomato. Hortic. Environ. Biotechnol. 2015, 56, 569–574. [Google Scholar] [CrossRef]
- Lara, O. Efecto de los Filtros Fotoselectivos en el Rendimiento y Calidad Postcosecha de Espinaca (Spinacia oleracea L.) “Baby” cv Viroflay Cultivada en Hidroponía. Master’s Thesis, Universidad de Chile, Santiago, Chile, 2019. [Google Scholar]
- Johkan, M.; Shoji, K.; Goto, F.; Hahida, S.; Yoshihara, T. Effect of green light wavelength and intensity on photomorphogenesis and photosynthesis in Lactuca sativa. Environ. Exp. Bot. 2012, 75, 128–133. [Google Scholar] [CrossRef]
- Kim, H.H.; Goins, G.D.; Wheeler, R.M.; Sager, J.C. Green light supplementation for enhaced lettuce growth under red and blue light emitting diodes. HortScience 2004, 39, 1617–1622. [Google Scholar] [CrossRef] [PubMed]
- Ilić, Z.; Milenković, L.; Šunić, L.; Barać, S.; Cvetković, D.; Stanojević, L.; Kevrešan, Z.; Mastilović, J. Bioactive constituents of red and green lettuce grown under colour shade nets. Emir. J. Food Agric. 2019, 31, 937–944. [Google Scholar] [CrossRef]
- Arias, J.P.; Zapata, K.; Rojano, B.; Arias, M. Effect of light wavelength on cell growth, content of phenolic compounds and antioxidant activity in cell suspension cultures of Thevetia peruviana. J. Photochem. Photobiol. Biol. 2016, 163, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Gálvez, A.; Albacete, A.; Del Amor, F.M.; López-Marín, J. The use of red shade nets improves growth in salinized pepper (Capsicum annuum L.) plants by regulating their ion homeostasis and hormone balance. Agronomy 2020, 10, 1766. [Google Scholar] [CrossRef]
- Elavarthi, S.; Martin, B. Spectrophotometric assays for antioxidant enzymes in plants. In Plant Stress Tolerance. Methods in Molecular Biology; Sunkar, R., Ed.; Springer: New York, NY, USA, 2010; Volume 639. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Wei, G.; Li, J.; Qian, Q.-Q.; Yu, J.-Q. Silicon Alleviates Salt Stress and Increases Antioxidant Enzymes Activity in Leaves of Salt-Stressed Cucumber (Cucumis Sativus L.). Plant Sci. 2004, 167, 527–533. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Chamberlain, C. Methods in Plant Histology, 50th ed.; University of Chicago: Chicago, IL, USA, 1932. [Google Scholar]
- Johansen, D. Plant Microtechnique; McGraw-Hill Book Co.: New York, NY, USA, 1940. [Google Scholar]
- D’ Ambrogio de Argüeso, A. Manual de Técnicas en Histología Vegetal; Hemisferio Sur Editorial: Buenos Aires, Argentina, 1986. [Google Scholar]
- Singh, D.; Mathur, S. Histopathology of Seed-Borne Infections; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Temperini, O.; Calabrese, N.; Temperini, A.; Rouphael, Y.; Tesi, R.; Lenzi, A.; Carito, A.; Colla, G. Grafting artichoke onto cardoon rootstocks: Graft compatibility, yield and Verticillium wilt incidence. Sci. Hortic. 2013, 149, 22–27. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Treatment | Effect | Reference |
|---|---|---|---|
| Solanum lycopersicum L. cv. Komeett | Green light | Increases in stem length, and fresh and dry weight. Leaf size is reduced | [31] |
| Solanum lycopersicum L. cv. Ailsa Craig | Green light | Under water stress conditions, lipid peroxidation (MDA) decreased, and photosynthetic capacity was maintained. | [32] |
| Solanum lycopersicum cv. Ingar F1 | Red and green light | Green light increased the efficiency of light use in young plants, resulted in stem elongation, and decreased biomass compared with red light, which also increased the photosynthetic rate. | [33] |
| Ocimum basilicum L. | Green light | Biomass, number of leaves, stem length, and leaf area increased. | [34] |
| Ocimum sp. | Green net | Fresh and dry weight increased, and so did the number of shoots/plant. | [35] |
| Spinacia oleracea L. | Red net | Fresh weight, total phenolic content, and antioxidant capacity increased. | [45] |
| Capsicum annuum L. | Red net | Increased water-use efficiency, phytohormone expression, and plant height. | [50] |
| Solanum lycopersicum Mill. var. “Sida” | Red net | Increases in biomass, shoot/root ratio growth, height, and leaf area. | [37] |
| Capsicum annuum L. | Red and green net | Both increased leaf area and commercial yield. | [36] |
| Solanum lycopersicum L. cv. Gangmu No. 1 | Red and blue light | Promoted rapid cell proliferation | [38] |
| Cunninghamia lanceolata | Green light Tissue culture | Enhanced root growth in tissue culture. Increased the production of antioxidant enzymes, and decreased lipid peroxidation. | [42] |
| Solanum lycopersicum L. cv. Beaded Curtain and Rootstock No. 1 | Green light | Increased the production of antioxidant enzymes and improved vascular regeneration. | [43] |
| Withania somnifera L. | Red light Tissue culture | Increased callus formation in tissue culture, biomass accumulation, and antioxidant activity. | [19] |
| Stage or Process | Duration of Stage | Duration of Exposure | Total Age of Plant | Environmental Conditions |
|---|---|---|---|---|
| Sowing and growth | 30 days | 0 days | 30 days | 92.8% RH and 16.2 °C |
| Grafting chamber | 3 h | 0 days | 30 days | 96.8% RH and 17.3 °C |
| Incubation chamber | 4 days | 0 days | 34 days | 98.8% RH and 24.5 °C |
| Curing chamber 1 | 3 days | 3 days | 37 days | 96.8% RH, 17.1 °C heating with furnace, and the opening/closing of curtains during the day/night |
| Curing chamber 2 | 4 days | 7 days | 41 days | 92.4% RH, 16.4 °C, and the opening/closing of curtains during the day/night |
| Acclimation | 7 days | 14 days | 48 days | 92.9% RH and 15.1 °C |
| Apical incision | 2 h | 15 days | 49 days | 92.9% RH and 15.1 °C |
| Shoot growth 1 | 7 days | 21 days | 55 days | 92.7% RH and 14.9 °C |
| Shoot growth 2 | 7 days | 28 days | 62 days | 92.6% RH and 14.9 °C |
| Ready plant | 62 days | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carmach, C.; Castro, M.; Peñaloza, P.; Guzmán, L.; Marchant, M.J.; Valdebenito, S.; Kopaitic, I. Positive Effect of Green Photo-Selective Filter on Graft Union Formation in Tomatoes. Plants 2023, 12, 3402. https://doi.org/10.3390/plants12193402
Carmach C, Castro M, Peñaloza P, Guzmán L, Marchant MJ, Valdebenito S, Kopaitic I. Positive Effect of Green Photo-Selective Filter on Graft Union Formation in Tomatoes. Plants. 2023; 12(19):3402. https://doi.org/10.3390/plants12193402
Chicago/Turabian StyleCarmach, Constanza, Mónica Castro, Patricia Peñaloza, Leda Guzmán, María José Marchant, Samuel Valdebenito, and Iván Kopaitic. 2023. "Positive Effect of Green Photo-Selective Filter on Graft Union Formation in Tomatoes" Plants 12, no. 19: 3402. https://doi.org/10.3390/plants12193402
APA StyleCarmach, C., Castro, M., Peñaloza, P., Guzmán, L., Marchant, M. J., Valdebenito, S., & Kopaitic, I. (2023). Positive Effect of Green Photo-Selective Filter on Graft Union Formation in Tomatoes. Plants, 12(19), 3402. https://doi.org/10.3390/plants12193402