

Comparative Transcriptome Analysis Reveals the Interaction of Sugar and Hormone Metabolism Involved in the Root Hair Morphogenesis of the Endangered Fir Abies beshanzuensis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
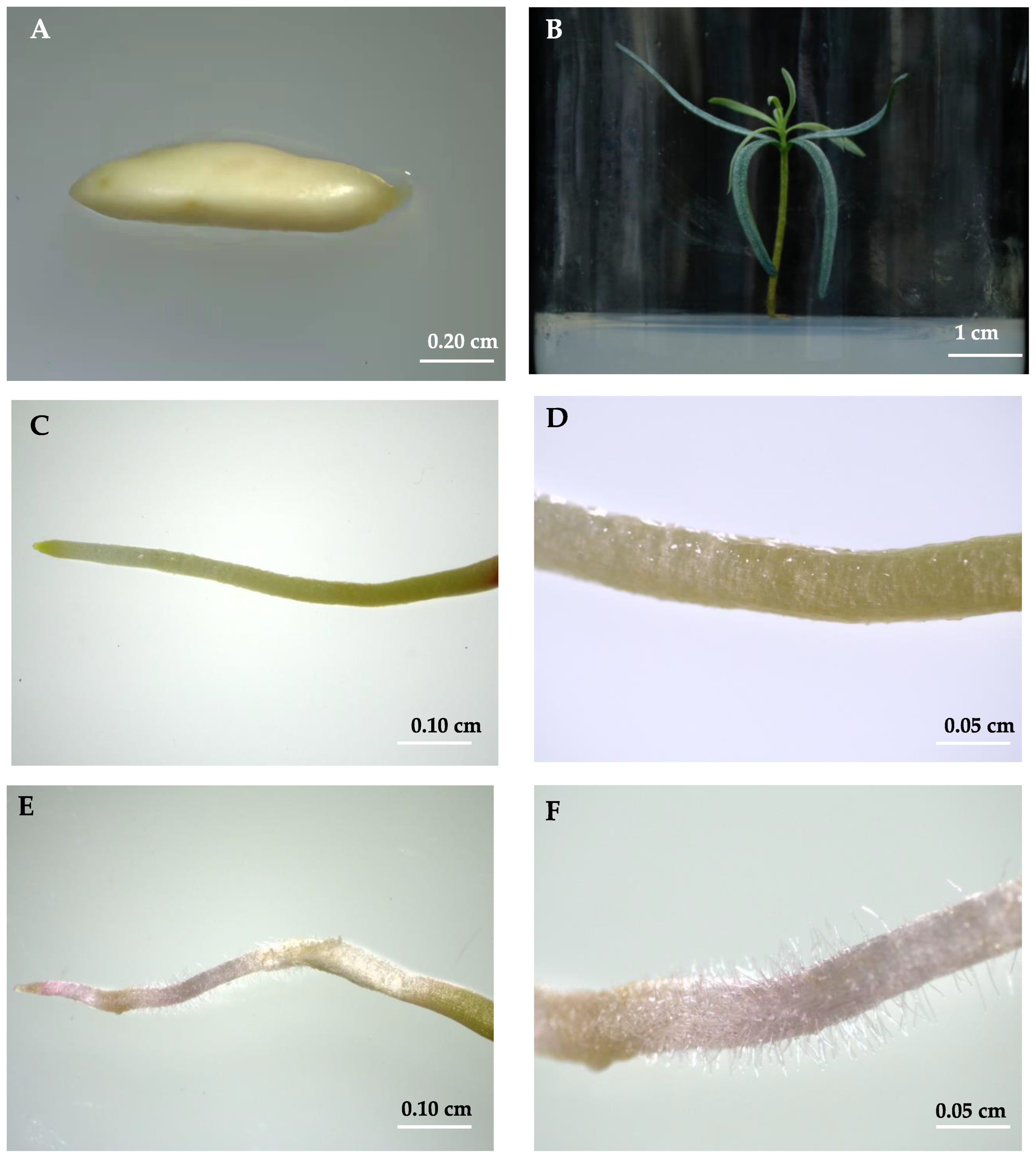
2.1. Comparison of Morphology from Test-Tube Plantlets of A. beshanzuensis In Vitro Conditions
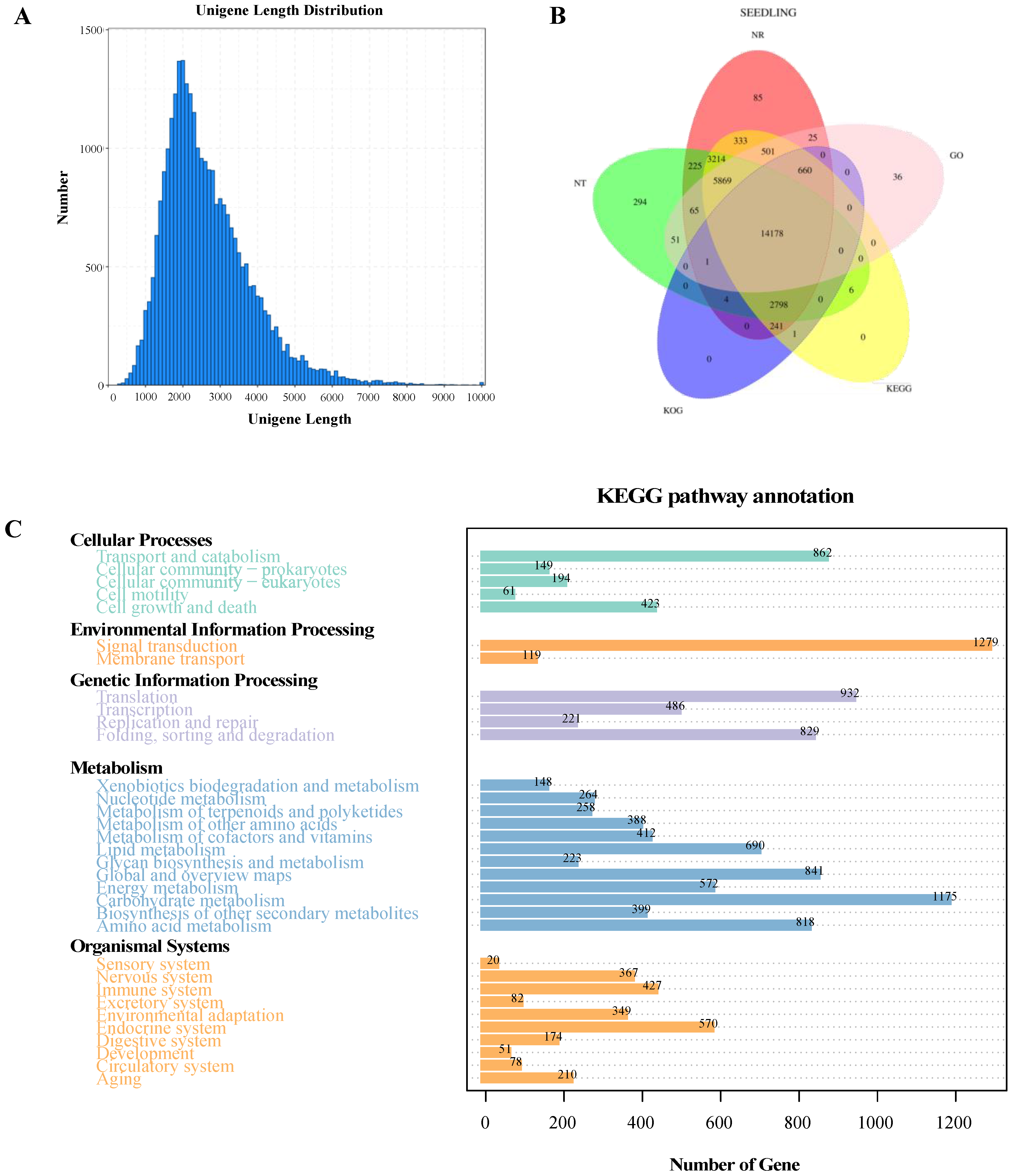
2.2. RNA-Seq Sequencing Analysis and Functional Annotation
2.3. Comparative Analysis of DEGs during the Development of the A. beshanzuensis Root with/without Root Hair
2.4. Identification of DEGs of Hormone Signals Involved in Root Hair Growth and Development
2.5. Identification of DEGs of Sugar Metabolism Involved in Root Hair Growth and Development
2.6. Interaction Analysis of Sucrose and Hormones Metabolism Signals Involved in Root Hair Development
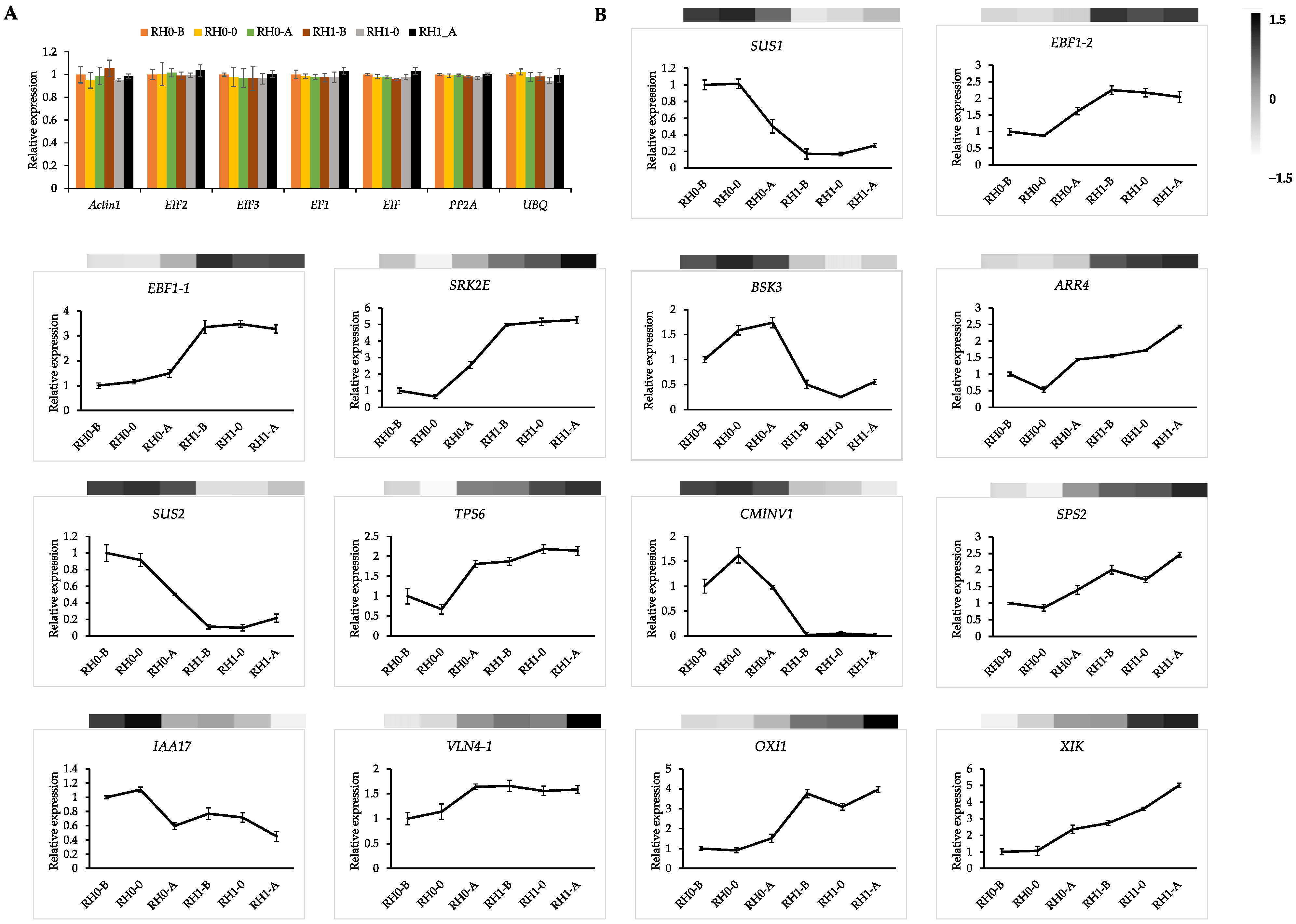
2.7. qPCR Validation for Reference Genes and DEGs of Sucrose and Hormones Signals Involved in Root Hair
3. Discussion
3.1. Root Hair Acts as a Pivotal Survival Strategy for A. beshanzuensis Breaking out of the Strong-Dependent Mycorrhizal Symbiosis
3.2. Root Hair Morphogenesis of A. beshanzuensis Can Be Regulated by Hormone and Sugar Signaling
3.3. The Interaction of Hormone and Sugar Metabolism Pathway Plays an Indispensable Role in Root Hair Growth and Development
4. Conclusions
5. Materials and Methods
5.1. Plant Materials
5.2. Immature Embryo Culture
5.3. RNA Sequencing and Data Analysis
5.4. Network Analysis of Key Genes Involved in Root Hair Grow and Development in A. beshanzuensis
5.5. Selection of Candidate Reference Genes
5.6. Quantitative Real-Time PCR (qRT-PCR) Validation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, Y.; Zhang, D.; Luscombe, D.; Liao, W.; Farjon, A.; Katsuki, T.; Xiang, Q.; Li, N. Abies beshanzuensis, The IUCN red list of threatened species 2013. RLTS 2013, 10, eT32318A2814360. [Google Scholar]
- Fu, L.K.; Jin, J.M. China Plant Red Data Book: Rare and Endangered Plants; Science Press: Beijing, China, 1992; pp. 50–51. (In Chinese) [Google Scholar]
- The State Forestry Administration and the Ministry of Agriculture. List of Wild Plants of National Priority Protection. 1999. Available online: http://www.gov.cn/gongbao/content/2000/content_60072.htm (accessed on 9 September 1999).
- Shao, Y.Z.; Hu, J.T.; Fan, P.Z.; Liu, Y.Y.; Wang, Y.H. The complete chloroplast genome sequence of Abies beshanzuensis, a highly endangered fir species from south China. Mitochondrial DNA B Resour. 2018, 3, 921–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maherali, H. Is there an association between root architecture and mycorrhizal growth response? New Phytol. 2014, 204, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.X.; Peng, C.; Yu, S.; Hu, J.F.; Li, J. Aflatoxins from the endophytic fungus Aspergillus sp. Y-2 isolated from the critically endangered conifer Abies beshanzuensis. Nat. Prod. Res. 2021, 35, 3248–3253. [Google Scholar] [CrossRef] [PubMed]
- Olanrewaju, O.S.; Ayangbenro, A.S.; Glick, B.R.; Babalola, O.O. Plant health: Feedback effect of root exudates-rhizobiome interactions. Appl. Microbiol. Biotechnol. 2019, 103, 1155–1166. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Liu, K.; Wu, Y.; Cao, L.; Wang, T.; Xu, D.; Yu, N.; Xiang, X.; Zhou, R.; Yu, M.; et al. A method for quick breeding of Abies beshanzuensis employing embryo rescue technology. In WO Patent; ZL2018100080786, 2019. Available online: http://epub.cnipa.gov.cn/Dxb/IndexQuery (accessed on 1 November 2022).
- Liu, K.; Liu, B.; Yuan, L.; Shuai, H.; Yang, Y.; Wang, T.J.; Chen, D.L.; Chen, X.R.; Yang, K.B.; Jin, X.F.; et al. Seed development and embryo culture of endangered Abies beshanzuensis. Chin. Bull. Bot. 2021, 56, 573–583. [Google Scholar]
- Puig, J.; Pauluzzi, G.; Guiderdoni, E.; Gantet, P. Regulation of shoot and root development through mutual signaling. Mol. Plant. 2012, 5, 974–983. [Google Scholar] [CrossRef] [Green Version]
- Ljung, K.; Nemhauser, J.L.; Perata, P. New mechanistic links between sugar and hormone signalling networks. Curr. Opin. Plant Biol. 2015, 25, 130–137. [Google Scholar] [CrossRef]
- Bates, T.R.; Lynch, J.P. Stimulation of root hair elongation in Arabidopsis thaliana by low phosphorus availability. Plant Cell Environ. 1996, 19, 529–538. [Google Scholar] [CrossRef]
- Emons, A.M.C.; Ketelaar, T. Root Hairs. In Plant Cell Monographs; Springer: Berlin/Heidelberg, Germany, 2009; Volume 12. [Google Scholar]
- Haling, R.E.; Brown, L.K.; Bengough, A.G.; Valentine, T.A.; White, P.J.; Young, I.M.; George, T.S. Root hair length and rhizosheath mass depend on soil porosity, strength and water content in barley genotypes. Planta 2014, 239, 643–651. [Google Scholar] [CrossRef]
- Krasilnikoff, G.; Gahoonia, T.; Nielsen, N.E. Variation in phosphorus uptake efficiency by genotypes of cowpea (Vigna unguiculata) due to differences in root and root hair length and induced rhizosphere processes. Plant Soil 2003, 251, 83–91. [Google Scholar] [CrossRef]
- Tanaka, N.; Kato, M.; Tomioka, R.; Kurata, R.; Fukao, Y.; Aoyama, T.; Maeshima, M. Characteristics of a root hair-less line of Arabidopsis thaliana under physiological stresses. J. Exp. Bot. 2014, 65, 1497–1512. [Google Scholar] [CrossRef] [PubMed]
- Rongsawat, T.; Peltier, J.B.; Boyer, J.C.; Very, A.A.; Sentenac, H. Looking for Root Hairs to Overcome Poor Soils. Trends Plant Sci. 2021, 26, 83–94. [Google Scholar] [CrossRef]
- Dolan, L.; Duckett, C.M.; Grierson, C.; Linstead, P.; Schneider, K.; Lawson, E.; Dean, C.; Poethig, S.; Roberts, K. Clonal Relationships and Cell Patterning in the Root Epidermis of Arabidopsis. Development 1994, 120, 2465–2474. [Google Scholar]
- Yang, C.; Czech, L.; Gerboth, S.; Kojima, S.I.; Scita, G.; Svitkina, T. Novel roles of formin mDia2 in lamellipodia and filopodia formation in motile cells. PLoS Biol. 2007, 5, e317. [Google Scholar] [CrossRef] [Green Version]
- Vissenberg, K.; Claeijs, N.; Balcerowicz, D.; Schoenaers, S. Hormonal regulation of root hair growth and responses to the environment in Arabidopsis. J. Exp. Bot. 2020, 71, 2412–2427. [Google Scholar] [CrossRef] [Green Version]
- Schoenaers, S.; Balcerowicz, D.; Vissenberg, K. Molecular Mechanisms Regulating Root Hair Tip Growth: A Comparison with Pollen Tubes. In Pollen Tip Growth, Obermeyer, G., Feijó, J.A., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 167–243. [Google Scholar]
- Masucci, J.D.; Schiefelbein, J.W. The Rhd6 Mutation of Arabidopsis-Thaliana Alters Root-Hair Initiation through an Auxin-Associated and Ethylene-Associated Process. Plant Physiol. 1994, 106, 1335–1346. [Google Scholar] [CrossRef] [Green Version]
- Porco, S.; Pencik, A.; Rashed, A.; Voss, U.; Casanova-Saez, R.; Bishopp, A.; Golebiowska, A.; Bhosale, R.; Swarup, R.; Swarup, K.; et al. Dioxygenase-encoding AtDAO1 gene controls IAA oxidation and homeostasis in Arabidopsis. Proc. Natl. Acad. Sci. USA 2016, 113, 11016–11021. [Google Scholar] [CrossRef] [Green Version]
- Mangano, S.; Denita-Juarez, S.P.; Choi, H.S.; Marzol, E.; Hwang, Y.; Ranocha, P.; Velasquez, S.M.; Borassi, C.; Barberini, M.L.; Aptekmann, A.A.; et al. Molecular link between auxin and ROS-mediated polar growth. Proc. Natl. Acad. Sci. USA 2017, 114, 5289–5294. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.S.; Seo, M.; Cho, H.T. Two TPL-Binding Motifs of ARF2 Are Involved in Repression of Auxin Responses. Front. Plant Sci. 2018, 9, 372. [Google Scholar] [CrossRef] [Green Version]
- Pitts, R.J.; Cernac, A.; Estelle, M. Auxin and ethylene promote root hair elongation in Arabidopsis. Plant J. 1998, 16, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Yi, K.; Menand, B.; Bell, E.; Dolan, L. A basic helix-loop-helix transcription factor controls cell growth and size in root hairs. Nat. Genet. 2010, 42, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Huang, L.L.; Yan, A.; Liu, Y.H.; Liu, B.H.; Yu, C.Y.; Zhang, A.D.; Schiefelbein, J.; Gan, Y.B. Multiple phytohormones promote root hair elongation by regulating a similar set of genes in the root epidermis in Arabidopsis. J. Exp. Bot. 2016, 67, 6363–6372. [Google Scholar] [CrossRef]
- Vanstraelen, M.; Benkova, E. Hormonal Interactions in the Regulation of Plant Development. Annu. Rev. Cell Dev. Biol. 2012, 28, 463–487. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Xu, P.; Li, B.S.; Li, P.P.; Wen, X.; An, F.Y.; Gong, Y.; Xin, Y.; Zhu, Z.Q.; Wang, Y.C.; et al. Ethylene promotes root hair growth through coordinated EIN3/EIL1 and RHD6/RSL1 activity in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, 13834–13839. [Google Scholar] [CrossRef] [Green Version]
- Qin, H.; Huang, R.F. Auxin Controlled by Ethylene Steers Root Development. Int. J. Mol. Sci. 2018, 19, 3656. [Google Scholar] [CrossRef] [Green Version]
- Gewin, V. Food an Underground Revolution. Nature 2010, 466, 552–553. [Google Scholar] [CrossRef] [Green Version]
- Handford, M.; Rodriguez-Furlan, C.; Marchant, L.; Segura, M.; Gomez, D.; Alvarez-Buylla, E.; Xiong, G.Y.; Pauly, M.; Orellana, A. Arabidopsis thaliana AtUTr7 Encodes a Golgi-Localized UDP-Glucose/UDP-Galactose Transporter that Affects Lateral Root Emergence. Mol. Plant 2012, 5, 1263–1280. [Google Scholar] [CrossRef] [Green Version]
- Harigaya, W.; Takahashi, H. Effects of glucose and ethylene on root hair initiation and elongation in lettuce (Lactuca sativa L.) seedlings. J. Plant Res. 2018, 131, 543–554. [Google Scholar] [CrossRef]
- Pang, C.Y.; Wang, H.; Pang, Y.; Xu, C.; Jiao, Y.; Qin, Y.M.; Western, T.L.; Yu, S.X.; Zhu, Y.X. Comparative Proteomics Indicates That Biosynthesis of Pectic Precursors Is Important for Cotton Fiber and Arabidopsis Root Hair Elongation. Mol. Cell. Proteom. 2010, 9, 2019–2033. [Google Scholar] [CrossRef] [Green Version]
- Raya-Gonzalez, J.; Lopez-Bucio, J.S.; Prado-Rodriguez, J.C.; Ruiz-Herrera, L.F.; Guevara-Garcia, A.A.; Lopez-Bucio, J. The MEDIATOR genes MED12 and MED13 control Arabidopsis root system configuration influencing sugar and auxin responses. Plant Mol. Biol. 2017, 95, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Libault, M.; Brechenmacher, L.; Cheng, J.; Xu DStacey, G. Root hair systems biology. Trends Plant Sci. 2010, 15, 641–650. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, Y.; Du, F.; Cao, L.; Dong, H.; Ren, H. Arabidopsis VILLIN4 is involved in root hair growth through regulating actin organization in a Ca2+-dependent manner. New Phytol. 2011, 190, 667–682. [Google Scholar] [CrossRef] [PubMed]
- Stetter, M.G.; Schmid, K.; Ludewig, U. Uncovering Genes and Ploidy Involved in the High Diversity in Root Hair Density, Length and Response to Local Scarce Phosphate in Arabidopsis thaliana. PLoS ONE 2015, 10, e0120604. [Google Scholar] [CrossRef] [Green Version]
- Nestler, J.; Liu, S.Z.; Wen, T.J.; Paschold, A.; Marcon, C.; Tang, H.M.; Li, D.L.; Li, L.; Meeley, R.B.; Sakai, H.; et al. Roothairless5, which functions in maize (Zea mays L.) root hair initiation and elongation encodes a monocot-specific NADPH oxidase. Plant J. 2014, 79, 729–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nestler, J.; Keyes, S.D.; Wissuwa, M. Root hair formation in rice (Oryza sativa L.) differs between root types and is altered in artificial growth conditions. J. Exp. Bot. 2016, 67, 3699–3708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.B.; Xie, Z.Z.; Hu, C.G.; Zhang, J.Z. A review of auxin response factors (ARFs) in plants. Front. Plant Sci. 2016, 7, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohme, K.; Li, Y.; Charlot, F.; Grierson, C.; Marrocco, K.; Okada, K.; Laloue, M.; Nogue, F. The Arabidopsis COW1 gene encodes a phosphatidylinositol transfer protein essential for root hair tip growth. Plant J. 2004, 40, 686–698. [Google Scholar] [CrossRef]
- Xiao, S.; Liu, L.T.; Zhang, Y.J.; Sun, H.C.; Zhang, K.; Bai, Z.Y.; Dong, H.Z.; Li, C.D. Fine root and root hair morphology of cotton under drought stress revealed with RhizoPot. J. Agron. Crop Sci. 2020, 206, 679–693. [Google Scholar] [CrossRef]
- Klinsawang, S.; Sumranwanich, T.; Wannaro, A.; Saengwilai, P. Effects of Root Hair Length on Potassium Acquisition in Rice (Oryza sativa L.). Appl. Ecol. Environ. Res. 2018, 16, 1609–1620. [Google Scholar] [CrossRef]
- Kwasniewski, M.; Daszkowska-Golec, A.; Janiak, A.; Chwialkowska, K.; Nowakowska, U.; Sablok, G.; Szarejko, I. Transcriptome analysis reveals the role of the root hairs as environmental sensors to maintain plant functions under water-deficiency conditions. J. Exp. Bot. 2016, 67, 1079–1094. [Google Scholar] [CrossRef] [PubMed]
- Galloway, A.F.; Akhtar, J.; Marcus, S.E.; Fletcher, N.; Field, K.; Knox, P. Cereal root exudates contain highly structurally complex polysaccharides with soil-binding properties. Plant J. 2020, 103, 1666–1678. [Google Scholar] [CrossRef] [PubMed]
- Schweiger, P.F.; Robson, A.D.; Barrow, N.J. Root Hair Length Determines Beneficial Effect of a Glomus Species on Shoot Growth of Some Pasture Species. New Phytol. 1995, 131, 247–254. [Google Scholar] [CrossRef]
- Borassi, C.; Dorosz, J.G.; Ricardi, M.M.; Sardoy, M.C.; Fachin, L.P.; Marzol, E.; Mangano, S.; Garcia, D.R.R.; Pacheco, J.M.; Guerrero, Y.D.R.; et al. A cell surface arabinogalactan-peptide influences root hair cell fate. New Phytol. 2020, 227, 732–743. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Grones, P.; Robert, S. Auxin signaling: A big question to be addressed by small molecules. J. Exp. Bot. 2018, 69, 313–328. [Google Scholar] [CrossRef] [Green Version]
- Weijers, D.; Wagner, D. Transcriptional Responses to the Auxin Hormone. Annu. Rev. Plant Biol. 2016, 67, 539–574. [Google Scholar] [CrossRef]
- Knox, K.; Grierson, C.S.; Leyser, O. AXR3 and SHY2 interact to regulate root hair development. Development 2003, 130, 5769–5777. [Google Scholar] [CrossRef] [Green Version]
- Hwang, I.; Sheen, J.; Muller, B. Cytokinin Signaling Networks. Annu. Rev. Plant Biol. 2012, 63, 353–380. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.Z.; Hou, L.Y.; Meng, J.J.; You, H.W.; Li, Z.; Gong, Z.Z.; Yang, S.H.; Shi, Y.T. The Antagonistic Action of Abscisic Acid and Cytokinin Signaling Mediates Drought Stress Response in Arabidopsis. Mol. Plant 2018, 11, 970–982. [Google Scholar] [CrossRef] [Green Version]
- Rahman, A.; Hosokawa, S.; Oono, Y.; Amakawa, T.; Goto, N.; Tsurumi, S. Auxin and ethylene response interactions during Arabidopsis root hair development dissected by auxin influx modulators. Plant Physiol. 2002, 130, 1908–1917. [Google Scholar] [CrossRef] [Green Version]
- Pires, N.D.; Yi, K.K.; Breuninger, H.; Catarino, B.; Menand, B.; Dolan, L. Recruitment and remodeling of an ancient gene regulatory network during land plant evolution. Proc. Natl. Acad. Sci. USA 2013, 110, 9571–9576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Gonzalez, J.; Lacek, J.; Retzer, K. Dissecting Hierarchies between Light, Sugar and Auxin Action Underpinning Root and Root Hair Growth. Plants 2021, 10, 111. [Google Scholar] [CrossRef] [PubMed]
- Stepanova, A.N.; Yun, J.; Likhacheva, A.V.; Alonso, J.M. Multilevel interactions between ethylene and auxin in Arabidopsis roots. Plant. Cell. 2007, 19, 2169–2185. [Google Scholar] [CrossRef]
- Swarup, R.; Perry, P.; Hagenbeek, D.; Van Der Straeten, D.; Beemster, G.T.S.; Sandberg, G.; Bhalerao, R.; Ljung, K.; Bennett, M.J. Ethylene upregulates auxin biosynthesis in Arabidopsis seedlings to enhance inhibition of root cell elongation. Plant. Cell. 2007, 19, 2186–2196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zemlyanskaya, E.V.; Omelyanchuk, N.A.; Ubogoeva, E.V.; Mironova, V.V. Deciphering Auxin-Ethylene Crosstalk at a Systems Level. Int. J. Mol. Sci. 2018, 19, 4060. [Google Scholar] [CrossRef] [Green Version]
- Bruex, A.; Kainkaryam, R.M.; Wieckowski, Y.; Kang, Y.H.; Bernhardt, C.; Xia, Y.; Zheng, X.H.; Wang, J.Y.; Lee, M.M.; Benfey, P.; et al. A Gene Regulatory Network for Root Epidermis Cell Differentiation in Arabidopsis. PLoS Genet. 2012, 8, e1002446. [Google Scholar] [CrossRef] [Green Version]
- Nordstrom, A.; Tarkowski, P.; Tarkowska, D.; Norbaek, R.; Astot, C.; Dolezal, K.; Sandberg, G. Auxin regulation of cytokinin biosynthesis in Arabidopsis thaliana: A factor of potential importance for auxin-cytokinin-regulated development. Proc. Natl. Acad. Sci. USA 2004, 101, 8039–8044. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.L.; Moore, S.; Chen, C.L.; Lindsey, K. Crosstalk Complexities between Auxin, Cytokinin, and Ethylene in Arabidopsis Root Development: From Experiments to Systems Modeling, and Back Again. Mol. Plant 2017, 10, 1480–1496. [Google Scholar] [CrossRef] [Green Version]
- El-Showk, S.; Ruonala, R.; Helariutta, Y. Crossing paths: Cytokinin signalling and crosstalk. Development 2013, 140, 1373–1383. [Google Scholar] [CrossRef] [Green Version]
- Machin, D.C.; Hamon-Josse, M.; Bennett, T. Fellowship of the rings: A saga of strigolactones and other small signals. New Phytol. 2020, 225, 621–636. [Google Scholar] [CrossRef] [Green Version]
- Ruyter-Spira, C.; Kohlen, W.; Charnikhova, T.; van Zeijl, A.; van Bezouwen, L.; de Ruijter, N.; Cardoso, C.; Lopez-Raez, J.A.; Matusova, R.; Bours, R.; et al. Physiological Effects of the Synthetic Strigolactone Analog GR24 on Root System Architecture in Arabidopsis: Another Belowground Role for Strigolactones? Plant Physiol. 2011, 155, 721–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Liu, Q.; Zhou, Z.; Yin, H.; Xie, Y.; Wei, Y. Two terpene synthases in resistant Pinus massoniana contribute to defence against Bursaphelenchus xylophilus. Plant Cell Environ. 2021, 44, 257–274. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative pcr and the 2-−ΔΔCT method. Methods 2000, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Jiao, Y.; Zhao, Y.; Gao, M.; Wu, L.; Wang, S.; Yang, J.; Wang, J.; Chen, Y.; Wang, Y. Phytohormone and transcriptome of pericarp reveals jasmonate and LcMYC2 are involved in neral and geranial biosynthesis in Litsea cubeba. Ind. Crops Prod. 2022, 177, 114423. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, B.; Liu, K.; Chen, X.; Xiao, D.; Wang, T.; Yang, Y.; Shuai, H.; Wu, S.; Yuan, L.; Chen, L. Comparative Transcriptome Analysis Reveals the Interaction of Sugar and Hormone Metabolism Involved in the Root Hair Morphogenesis of the Endangered Fir Abies beshanzuensis. Plants 2023, 12, 276. https://doi.org/10.3390/plants12020276
Liu B, Liu K, Chen X, Xiao D, Wang T, Yang Y, Shuai H, Wu S, Yuan L, Chen L. Comparative Transcriptome Analysis Reveals the Interaction of Sugar and Hormone Metabolism Involved in the Root Hair Morphogenesis of the Endangered Fir Abies beshanzuensis. Plants. 2023; 12(2):276. https://doi.org/10.3390/plants12020276
Chicago/Turabian StyleLiu, Bin, Ke Liu, Xiaorong Chen, Duohong Xiao, Tingjin Wang, Yang Yang, Hui Shuai, Sumei Wu, Lu Yuan, and Liping Chen. 2023. "Comparative Transcriptome Analysis Reveals the Interaction of Sugar and Hormone Metabolism Involved in the Root Hair Morphogenesis of the Endangered Fir Abies beshanzuensis" Plants 12, no. 2: 276. https://doi.org/10.3390/plants12020276
APA StyleLiu, B., Liu, K., Chen, X., Xiao, D., Wang, T., Yang, Y., Shuai, H., Wu, S., Yuan, L., & Chen, L. (2023). Comparative Transcriptome Analysis Reveals the Interaction of Sugar and Hormone Metabolism Involved in the Root Hair Morphogenesis of the Endangered Fir Abies beshanzuensis. Plants, 12(2), 276. https://doi.org/10.3390/plants12020276