

Systematics and Phylogenetic Placement of Panicum L. Species within the Melinidinae Based on Morphological, Anatomical, and Molecular Data (Poaceae, Panicoideae, Paniceae)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
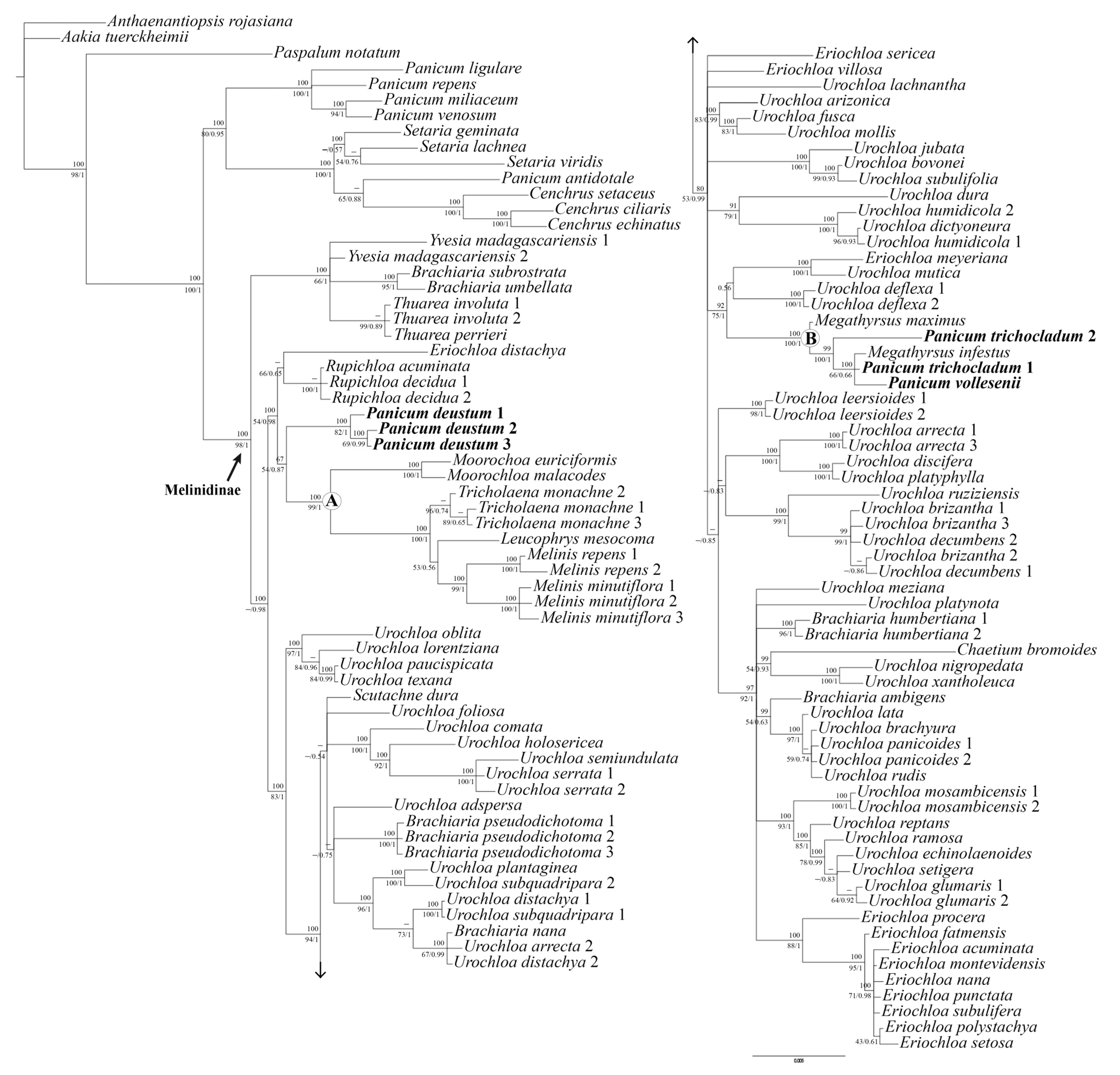
2. Results
2.1. Phylogenetic Reconstruction
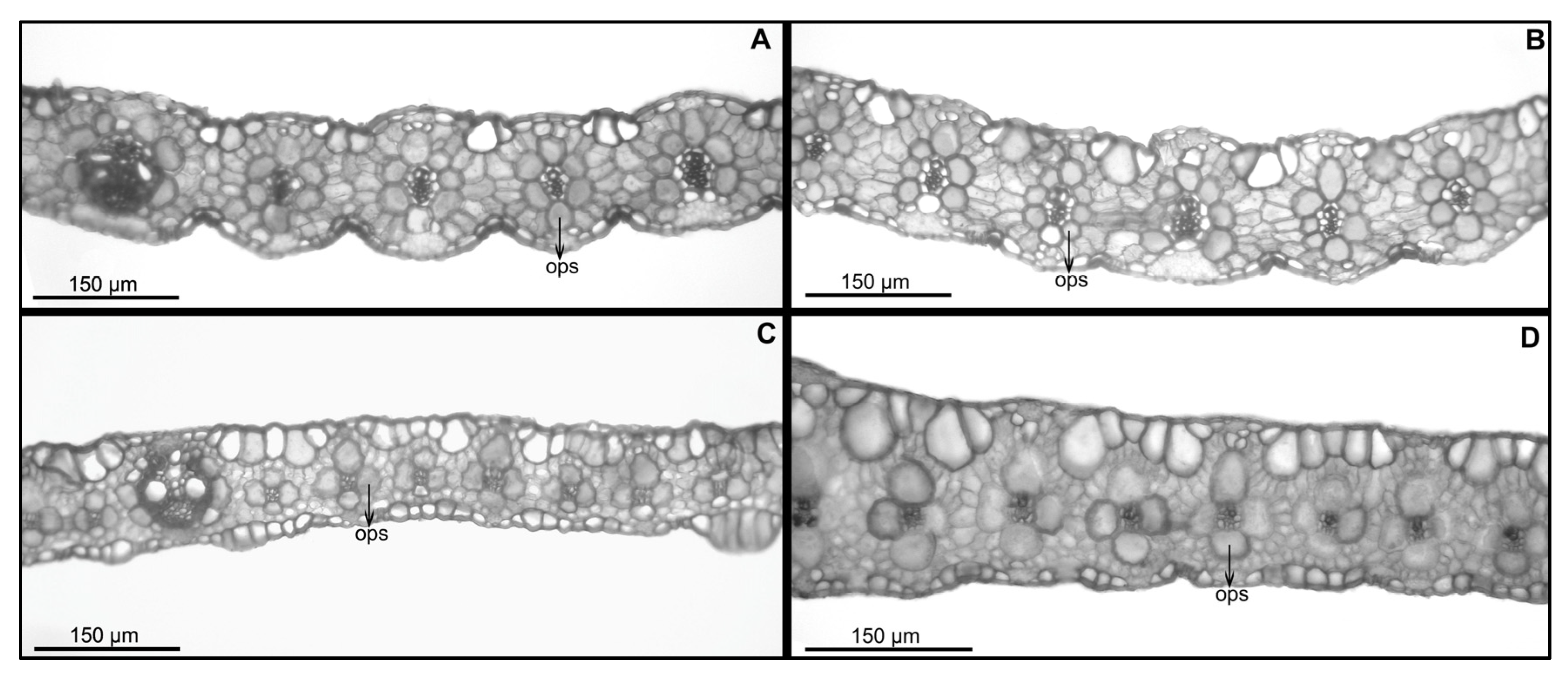
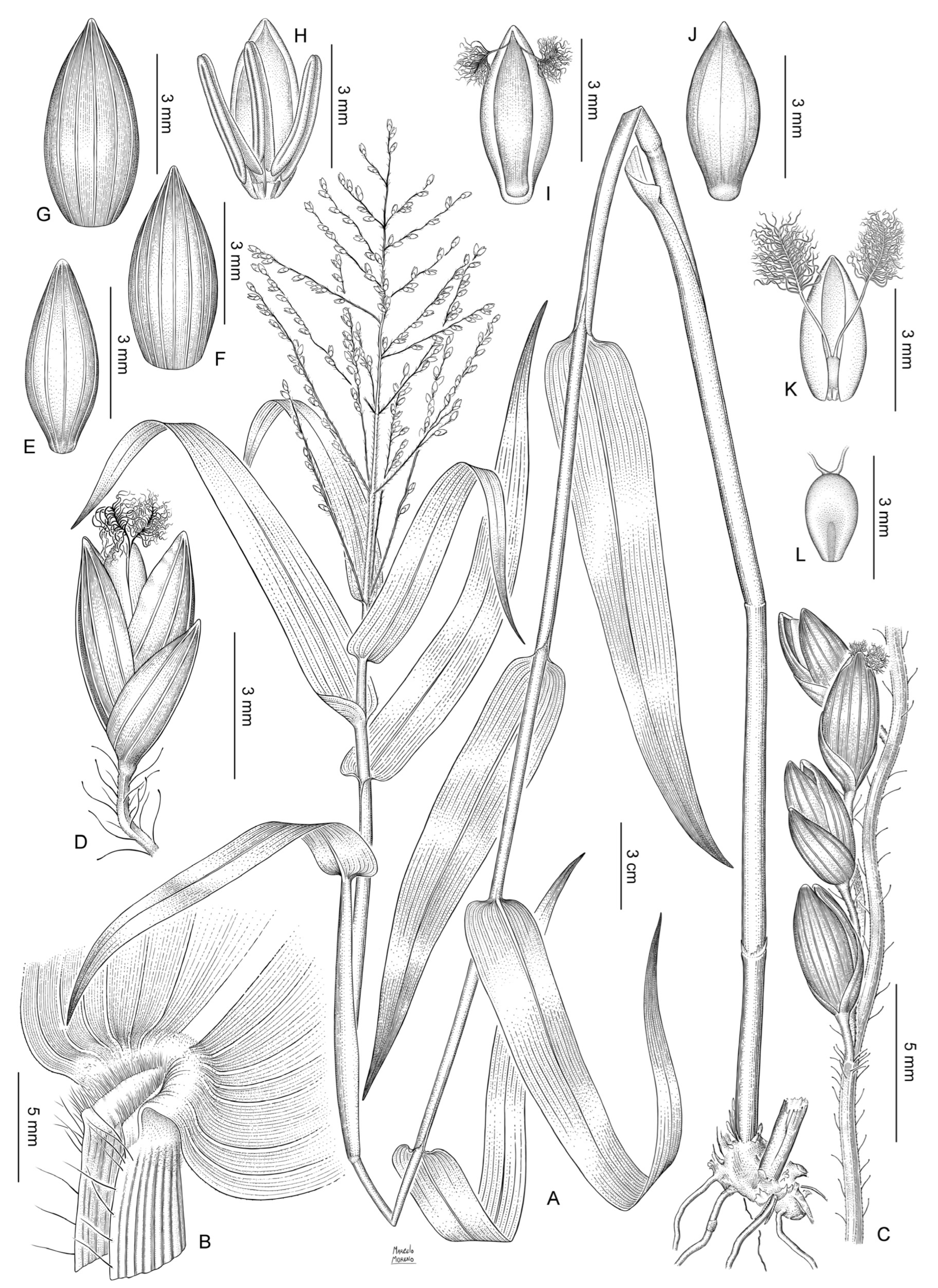
2.2. Anatomy and Morphology
2.2.1. Foliar Anatomy of Panicum deustum (Bequaert 3398, de Witte 10792)
2.2.2. Foliar Anatomy of Panicum trichocladum (Eichinger 5876, Hitchcock 24927) and P. vollesenii (Vollesen 3871)
2.2.3. Ornamentation of the Upper Anthecia
3. Discussion
| Key points to the genera of Melinidinae: | |
| 1. Lower and upper glume awned | Chaetium |
| 1′. Lower and upper glume muticous | 2 |
| 2(1′). Plants andromonoic; inflorescences deciduous as a whole at maturity; rachis broadly winged, foliaceous, and folded transversely to form a capsule | Thuarea |
| 2′. Plants not andromonoic; inflorescences not early deciduous; rachis triquetrous, subcylindric, or broadly winged | 3 |
| 3(2′). Upper anthecium smooth, shiny, and coriaceous to membranous or hyaline; apex of the upper anthecium muticous | 4 |
| 3′. Upper anthecium longitudinal or transversely rugose, with simple or verrucose papillae all over its surface, coriaceous to crustaceous; apex of the upper anthecium mucronate, crestate, or awned | 10 |
| 4(3). Spikelets adaxial | 5 |
| 4′. Spikelets abaxial | 6 |
| 5(4). Upper anthecium not stipitate, nearly as long as the upper glume and the lower lemma; upper glume and lower pilose, without transverse veinlets | Leucophrys |
| 5′. Upper anthecium stipitate, shorter than the upper glume and lower lemma; upper glume and lower lemma glabrous, with transverse veinlets | Eccoptocarpha |
| 6(4′). Plants annual; inflorescences digitate | Yvesia |
| 6′. Plants perennial; inflorescences not digitate | 7 |
| 7(6′). Upper anthecium falling with glumes and lower anthecium, not early deciduous, and laterally compressed | Melinis |
| 7′. Upper anthecium early deciduous, without the bracts of the spikelet, dorsiventrally compressed | 8 |
| 8(7′). Inflorescence racemose, not pyramidal, with spikelets secund on short pedicels | Moorochloa |
| 8′. Inflorescences lax, pyramidal, with spikelets not secund on long, capilliform pedicels | 9 |
| 9(8′). Inflorescence without glandular hairs; lower glume absent or vestigial; upper anthecium readily deciduous | Tricholaena |
| 9′. Inflorescence with glandular hairs; lower glume ⅘ to nearly as long as the spikelet; upper anthecium not readily deciduous | Batochloa |
| 10(3′). Spikelets with a globular bead-like callus at its base | Eriochloa |
| 10′. Spikelets without a globular bead-like callus at its base | 11 |
| 11(10′). Upper anthecium longitudinally rugose, with flat macrohairs at the apex | 12 |
| 11′. Upper anthecium transversely rugose to slightly or strongly reticulate, without flat macrohairs at the apex | 13 |
| 12(11). Spikelets arranged in racemose inflorescences. Endemic to the Caribbean | Scutachne |
| 12′. Spikelets dispersed in open, and no racemose inflorescences. Brazil | Rupichloa |
| 13(11′) Inflorescences lax, pyramidal, with third-order branches present | Megathyrsus |
| 13′. Inflorescences racemose or lax, but only with first- and second-order branches | Urochloa |
Taxonomic Treatment
4. Materials and Methods
4.1. Phylogenetic Reconstruction
4.1.1. Taxon Sampling
4.1.2. DNA Amplification and Sequencing
4.1.3. Phylogenetic Analyses
4.2. Anatomical and Morphological Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gallaher, T.J.; Peterson, P.M.; Soreng, R.J.; Zuloaga, F.O.; Li, D.Z.; Clark, L.G.; Tyrrell, C.D.; Welker, C.A.D.; Kellogg, E.A.; Teisher, J.K. Grass evolution through space and time: A biogeographical reconstruction of the Poaceae. J. Syst. Evol. 2022, 60, 522–569. [Google Scholar] [CrossRef]
- Pilger, R. Gramineae. III: Unterfamilie Panicoideae. In Die Natürlichen Pflanzenfamilien, 2nd ed.; Engler, A., Prantl, K., Eds.; Engelmann: Leipzig, Germany, 1940; Volume 14e, pp. 1–208. [Google Scholar]
- Soreng, R.; Peterson, P.M.; Zuloaga, F.O.; Romaschenko, K.; Clark, L.G.; Teisher, J.; Gillespie, L.; Barberá, P.; Welker, C.; Kellogg, E.A.; et al. A worldwide phylogenetic classification of the Poaceae (Gramineae) III: An update. J. Syst. Evol. 2022, 60, 476–521. [Google Scholar] [CrossRef]
- POWO (Plants of the World Online). Facilitated by the Royal Botanic Gardens, Kew. Available online: http://www.plantsoftheworldonline.org/ (accessed on 31 October 2022).
- Gómez-Martínez, R.; Culham, A. Phylogeny of the subfamily Panicoideae with emphasis on the tribe Paniceae: Evidence from the trnL-F cpDNA region. In Grasses: Systematics and Evolution; Jacobs, S.W.L., Everett, J.E., Eds.; Commonwealth Scientific and Industrial Research Organization (CSIRO) Publishing: Collingwood, Australia, 2000; pp. 136–140. [Google Scholar]
- Giussani, L.M.; Cota-Sánchez, J.H.; Zuloaga, F.O.; Kellogg, E.A. A molecular phylogeny of the grass subfamily Panicoideae (Poaceae) shows multiple origins of C4 photosynthesis. Am. J. Bot. 2001, 88, 1993–2012. [Google Scholar] [CrossRef]
- Aliscioni, S.S.; Giussani, L.M.; Zuloaga, F.O.; Kellogg, E.A. A molecular phylogeny of Panicum (Poaceae: Paniceae). Tests of monophyly and phylogenetic placement within the Panicoideae. Am. J. Bot. 2003, 90, 796–821. [Google Scholar] [CrossRef] [PubMed]
- Christin, P.A.; Salamin, N.; Savolainen, V.; Duvall, M.R.; Besnard, G. C4 photosynthesis evolved in grasses via parallel adaptive genetic changes. Curr. Biol. 2007, 17, 1241–1247. [Google Scholar] [CrossRef] [PubMed]
- Christin, P.A.; Besnard, G.; Samaritani, E.; Duvall, M.R.; Hodkinson, T.R.; Savolainen, V.; Salamin, N. Oligocene CO2 Decline Promoted C4 Photosynthesis in Grasses. Curr. Biol. 2008, 18, 37–43. [Google Scholar] [CrossRef]
- Vicentini, A.; Barber, J.C.; Aliscioni, S.S.; Giussani, L.M.; Kellogg, E.A. The age of the grasses and clusters of origins of C4 photosynthesis. Glob Chang Biol. 2008, 14, 2963–2977. [Google Scholar] [CrossRef]
- Salariato, D.L.; Zuloaga, F.O.; Giussani, L.M.; Morrone, O. Molecular phylogeny of the C4–PCK photosynthetic subtype group (Poaceae: Panicoideae: Paniceae) and evolutionary trends in the homogenization of inflorescences. Mol. Phylogenet. Evol. 2010, 56, 355–369. [Google Scholar] [CrossRef]
- Morrone, O.; Aagesen, L.; Scataglini, M.A.; Salariato, D.L.; Denham, S.S.; Chemisquy, M.A.; Sede, S.M.; Giussani, L.M.; Kellogg, E.A.; Zuloaga, F.O. Phylogeny of the Paniceae (Poaceae: Panicoideae): Integrating plastid DNA sequences and morphology into a new classification. Cladistics 2012, 28, 333–356. [Google Scholar] [CrossRef]
- Hattersley, P.W. Variations in Photosynthetic Pathway. In Grass Systematics and Evolution; Soderstrom, T.R., Hilu, K.W., Campbell, C.S., Barkworth, M.E., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1987; pp. 49–64. [Google Scholar]
- Hattersley, P.W.; Watson, L. Diversification of Photosynthesis. In Grass Evolution and Domestication; Chapman, G.P., Ed.; Cambridge University Press: London, UK, 1992; pp. 38–116. [Google Scholar]
- GPWG (Grass Phylogeny Working Group). Phylogeny and subfamilial classification of the Poaceae. Ann. Mo. Bot. Gard. 2001, 88, 373–457. [Google Scholar] [CrossRef]
- GPWG II (Grass Phylogeny Working Group II). New grass phylogeny resolves deep evolutionary relationships and discovers C4 origins. New Phytol. 2012, 193, 304–312. [Google Scholar] [CrossRef]
- Ellis, R.P. Distribution of the Kranz syndrome in the southern African Eragrostoideae and Panicoideae according to bundle sheath anatomy and cytology. Agroplantae 1977, 9, 73–109. [Google Scholar]
- Ellis, R.P. Leaf anatomy and systematics of Panicum (Poaceae: Panicoideae) in Southern Africa. Monogr. Syst. Bot. Mo. Bot. Gard. 1988, 25, 129–156. [Google Scholar]
- Clayton, W.D.; Renvoize, S.A. Genera graminum: Grasses of the world. Kew Bull. 1986, 13, 1–389. [Google Scholar]
- Zuloaga, F.O.; Salariato, D.L.; Scataglini, M.A. Molecular phylogeny of Panicum s. str. (Poaceae, Panicoideae, Paniceae) and insights into its biogeography and evolution. PLoS ONE 2018, 13, e0191529. [Google Scholar] [CrossRef] [PubMed]
- Duvall, M.R.; Noll, J.D.; Minn, A.H. Phylogenetics of Paniceae (Poaceae). Am. J. Bot. 2001, 88, 1988–1992. [Google Scholar] [CrossRef] [PubMed]
- Duvall, M.R.; Saar, D.E.; Grayburn, W.S.; Holbrook, G.P. Complex transitions between C3 and C4 photosynthesis during the evolution of Paniceae: A phylogenetic case study emphasizing the position of Steinchisma hians (Poaceae), a C3—C4 intermediate. Int. J. Plant Sci. 2003, 164, 949–958. [Google Scholar] [CrossRef]
- Zuloaga, F.O.; Salomón, L.; Scataglini, M.A. Phylogeny of sections Clavelligerae and Pectinatae of Panicum (Poaceae, Panicoideae, Paniceae). Plant Syst. Evol. 2015, 301, 1693–1711. [Google Scholar] [CrossRef]
- Dengler, N.G.; Dengler, R.E.; Donnelly, P.M.; Hattersley, P.W. Quantitative Leaf Anatomy of C3 and C4 Grasses (Poaceae): Bundle Sheath and Mesophyll Surface Area Relationships. Ann. Bot. 1994, 63, 241–255. [Google Scholar] [CrossRef]
- Watson, L.; Macfarlane, T.D.; Dallwitz, M.J. 1992 onwards. The Grass Genera of the World: Descriptions, Illustrations, Identification, and Information Retrieval; Including Synonyms, Morphology, Anatomy, Physiology, Phytochemistry, Cytology, Classification, Pathogens, World and Local Distribution, and References. Version: 14th September 2022. Available online: https://www.delta-intkey.com/grass/index.htm (accessed on 20 September 2022).
- De Wet, J.M.J. Chromosome numbers of a few South American grasses. Cytologia 1954, 19, 97–103. [Google Scholar] [CrossRef]
- Rao, P.N.; Mwasumbi, L.B. Chromosome number reports LXXII. Taxon 1981, 30, 701. [Google Scholar]
- Renvoize, S.A. A new species of Panicum (Gramineae) from Tanzania. Kew Bull. 1980, 35, 202. [Google Scholar] [CrossRef]
- Hackel, J.; Vorontsova, M.S.; Nanjarisoa, O.P.; Hall, R.C.; Razanatsoa, J.; Malakasi, P.; Besnard, G. Grass diversification in Madagascar: In situ radiation of two large C3 shade clades and support for a Miocene to Pliocene origin of C4 grassy biomes. J. Biogeogr. 2018, 45, 750–761. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. Bot. Soc. Amer. 1987, 19, 11–15. [Google Scholar]
- Olmstead, R.G.; Sweere, J.A. Combining data in phylogenetic systematics: An empirical approach using three molecular data sets in the Solanaceae. Syst. Biol. 1994, 43, 467–481. [Google Scholar] [CrossRef]
- Kelchner, S.; Clark, L.G. Molecular evolution and phylogenetic utility of the chloroplast rpl16 intron in Chusquea and the Bambusoideae (Poaceae). Mol. Phylogenet. Evol. 1997, 8, 385–397. [Google Scholar] [CrossRef]
- Zhang, W. Phylogeny of the grass family (Poaceae) from rpl16 intron sequence data. Mol. Phylogenet. Evol. 2000, 15, 385–397. [Google Scholar] [CrossRef]
- Cialdella, A.M.; Giussani, L.M.; Aagesen, L.; Zuloaga, F.O.; Morrone, O. A phylogeny of Piptochaetium based on a combined analysis including trnL-F, rpl16 and morphology. Syst. Bot. 2007, 32, 545–559. [Google Scholar] [CrossRef]
- Jordan, W.C.; Courtney, W.M.; Neigel, E.J. Low levels of intraspecific genetic variation at a rapidly evolving chloroplast DNA locus in North American duckwoods (Lemnaceae). Am. J. Bot. 1996, 83, 430–439. [Google Scholar] [CrossRef]
- Giussani, L.M.; Zuloaga, F.O.; Quarín, C.L.; Cota-Sánchez, J.H.; Ubayasena, K.; Morrone, O. Phylogenetic relationships in the genus Paspalum (Poaceae: Panicoideae: Paniceae): An assessment of the Quadrifaria and Virgata informal groups. Syst. Bot. 2009, 34, 32–43. [Google Scholar] [CrossRef]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipskia, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Fitch, W.M. Toward defining the course of evolution: Minimal change for a specific tree topology. Syst. Zool. 1971, 20, 406–416. [Google Scholar] [CrossRef]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Crandall, K.A. Phylogeny estimation and hypothesis testing using maximum likelihood. Annu. Rev. Ecol. Evol. Syst. 1997, 28, 437–466. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Larget, B.; Miller, R.E.; Ronquist, F. Potential applications and pitfalls of Bayesian inference of phylogeny. Syst. Biol. 2002, 51, 673–688. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Goloboff, P.A.; Farris, J.S.; Nixon, K.C. TNT, a free program for phylogenetic analysis. Cladistics 2008, 24, 774–786. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihoodbased phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, MS, USA; 2010; pp. 1–8. [Google Scholar] [CrossRef]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web-servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Ullah, F.; Gao, Y.; Sari, İ.; Jiao, R.-F.; Saqib, S.; Gao, X.-F. Macro-Morphological and Ecological Variation in Rosa sericea Complex. Agronomy 2022, 12, 1078. [Google Scholar] [CrossRef]
- Zaman, W.; Ullah, F.; Parmar, G.; Saqib, S.; Ayaz, A.; Park, S. Foliar micromorphology of selected medicinal Lamiaceae taxa and their taxonomic implication using scanning electron microscopy. Microsc. Res. Tech. 2022, 85, 3217–3236. [Google Scholar] [CrossRef]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff. New York Botanical Garden’s Virtual Herbarium. Available online: http://sweetgum.nybg.org/science/ih/ (accessed on 14 October 2022).
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; et al. International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code) Adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017; Regnum Vegetabile 159; Koeltz Botanical Books: Glashütten, Germany, 2018. [Google Scholar] [CrossRef]
- Cutler, D.F. Applied Plant Anatomy; Addison-Wesley Longman Ltd.: Boston, MA, USA, 1978; pp. 1–103. [Google Scholar]
- Ellis, R.P. A procedure for standardizing comparative leaf anatomy in the Poaceae. I. The leaf blade as viewed in transverse section. Bothalia 1976, 12, 65–109. [Google Scholar] [CrossRef]
- Ellis, R.P. A procedure for standardizing comparative leaf anatomy in the Poaceae. II. The epidermis as seen in surface view. Bothalia 1979, 12, 641–671. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delfini, C.; Salariato, D.L.; Aliscioni, S.S.; Zuloaga, F.O. Systematics and Phylogenetic Placement of Panicum L. Species within the Melinidinae Based on Morphological, Anatomical, and Molecular Data (Poaceae, Panicoideae, Paniceae). Plants 2023, 12, 399. https://doi.org/10.3390/plants12020399
Delfini C, Salariato DL, Aliscioni SS, Zuloaga FO. Systematics and Phylogenetic Placement of Panicum L. Species within the Melinidinae Based on Morphological, Anatomical, and Molecular Data (Poaceae, Panicoideae, Paniceae). Plants. 2023; 12(2):399. https://doi.org/10.3390/plants12020399
Chicago/Turabian StyleDelfini, Carolina, Diego L. Salariato, Sandra S. Aliscioni, and Fernando O. Zuloaga. 2023. "Systematics and Phylogenetic Placement of Panicum L. Species within the Melinidinae Based on Morphological, Anatomical, and Molecular Data (Poaceae, Panicoideae, Paniceae)" Plants 12, no. 2: 399. https://doi.org/10.3390/plants12020399
APA StyleDelfini, C., Salariato, D. L., Aliscioni, S. S., & Zuloaga, F. O. (2023). Systematics and Phylogenetic Placement of Panicum L. Species within the Melinidinae Based on Morphological, Anatomical, and Molecular Data (Poaceae, Panicoideae, Paniceae). Plants, 12(2), 399. https://doi.org/10.3390/plants12020399