
Nanopriming of Barley Seeds—A Shotgun Approach to Improve Germination under Salt Stress Conditions by Regulating of Reactive Oxygen Species



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
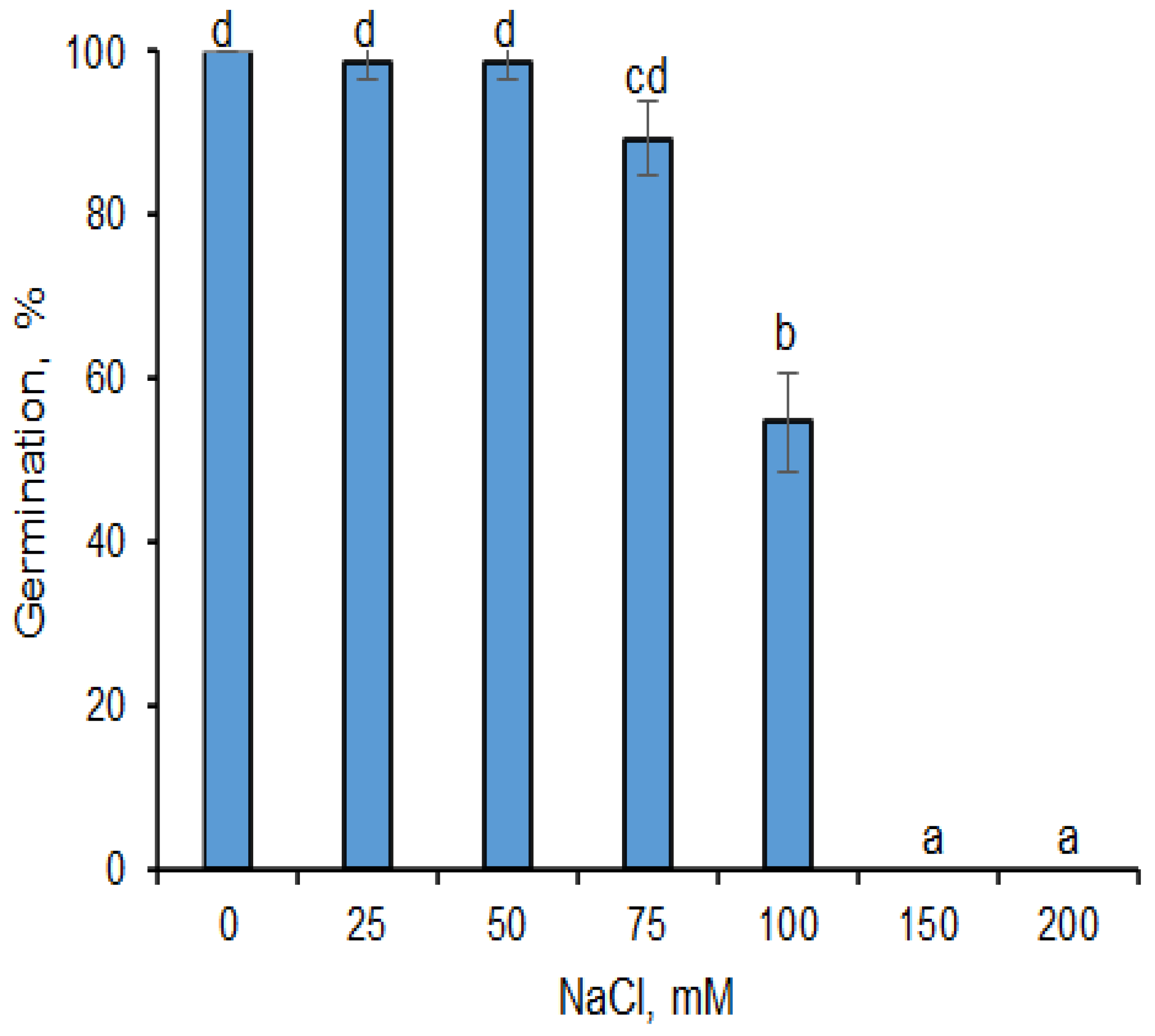
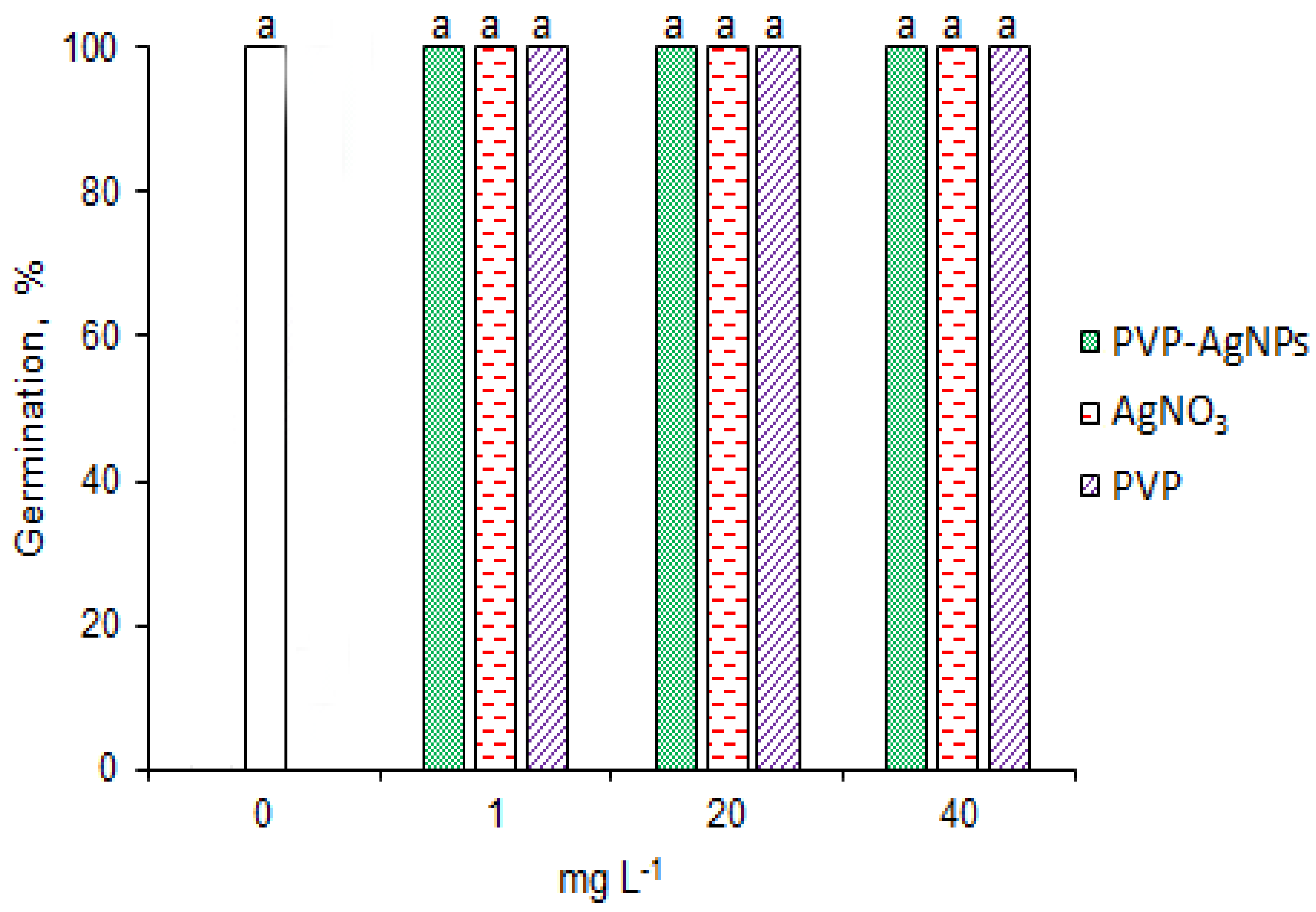
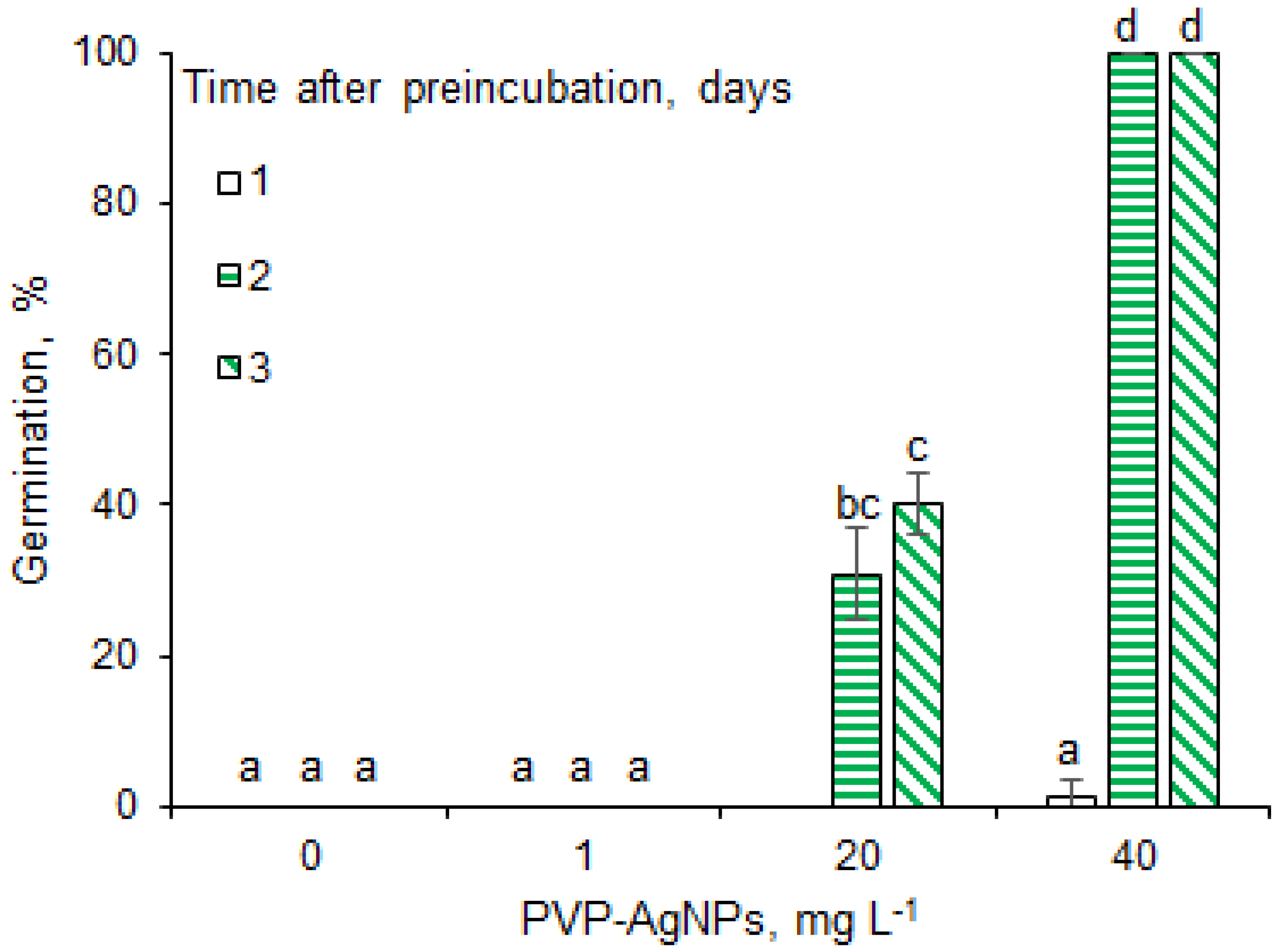
2.1. Effects of AgNPs on Seed Germination under Salinity Stress Conditions
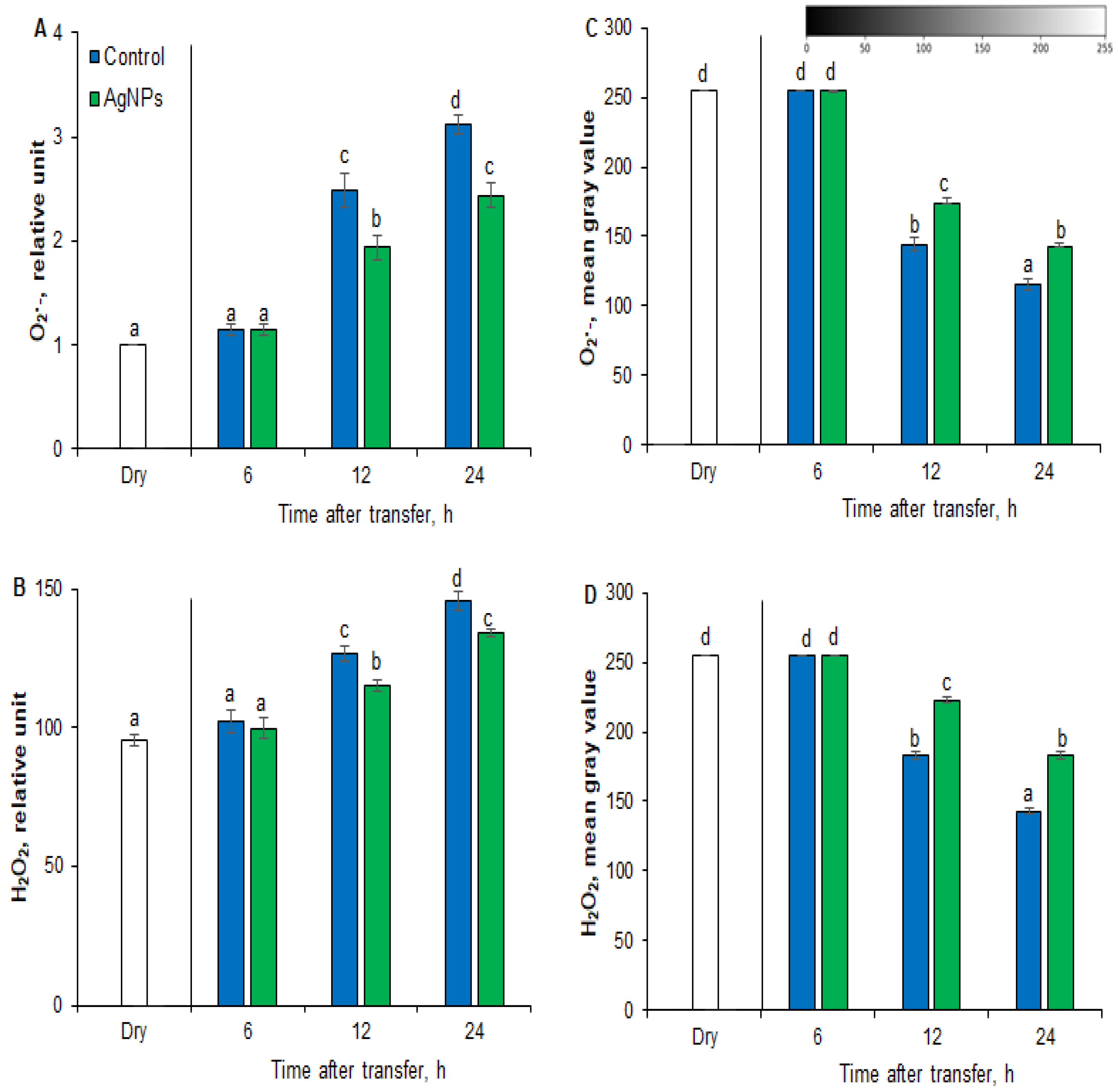
2.2. Effects of AgNPs on ROS Level
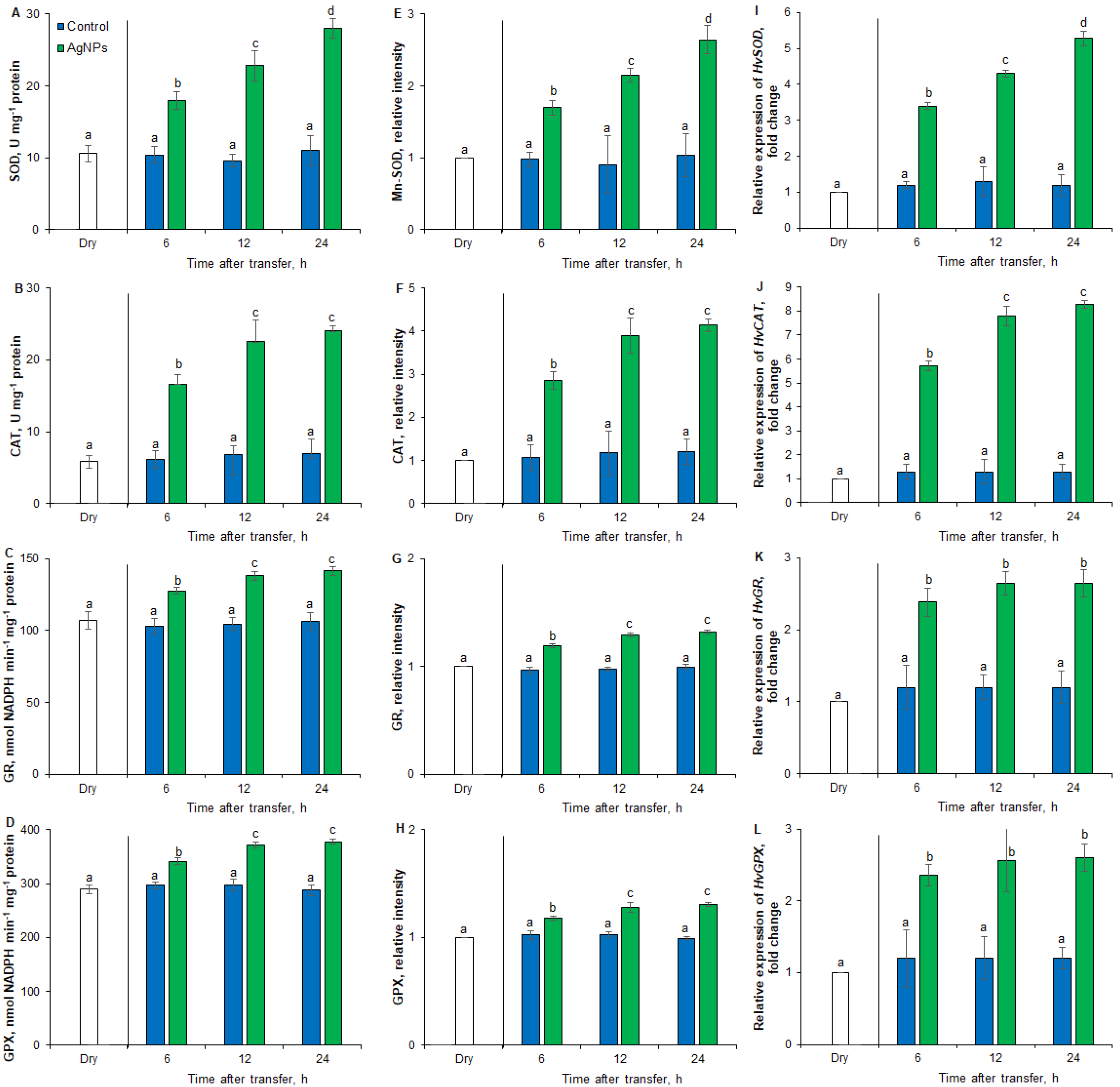
2.3. Effects of AgNPs on SOD, CAT, GR and GPX Activity and Gene Expression
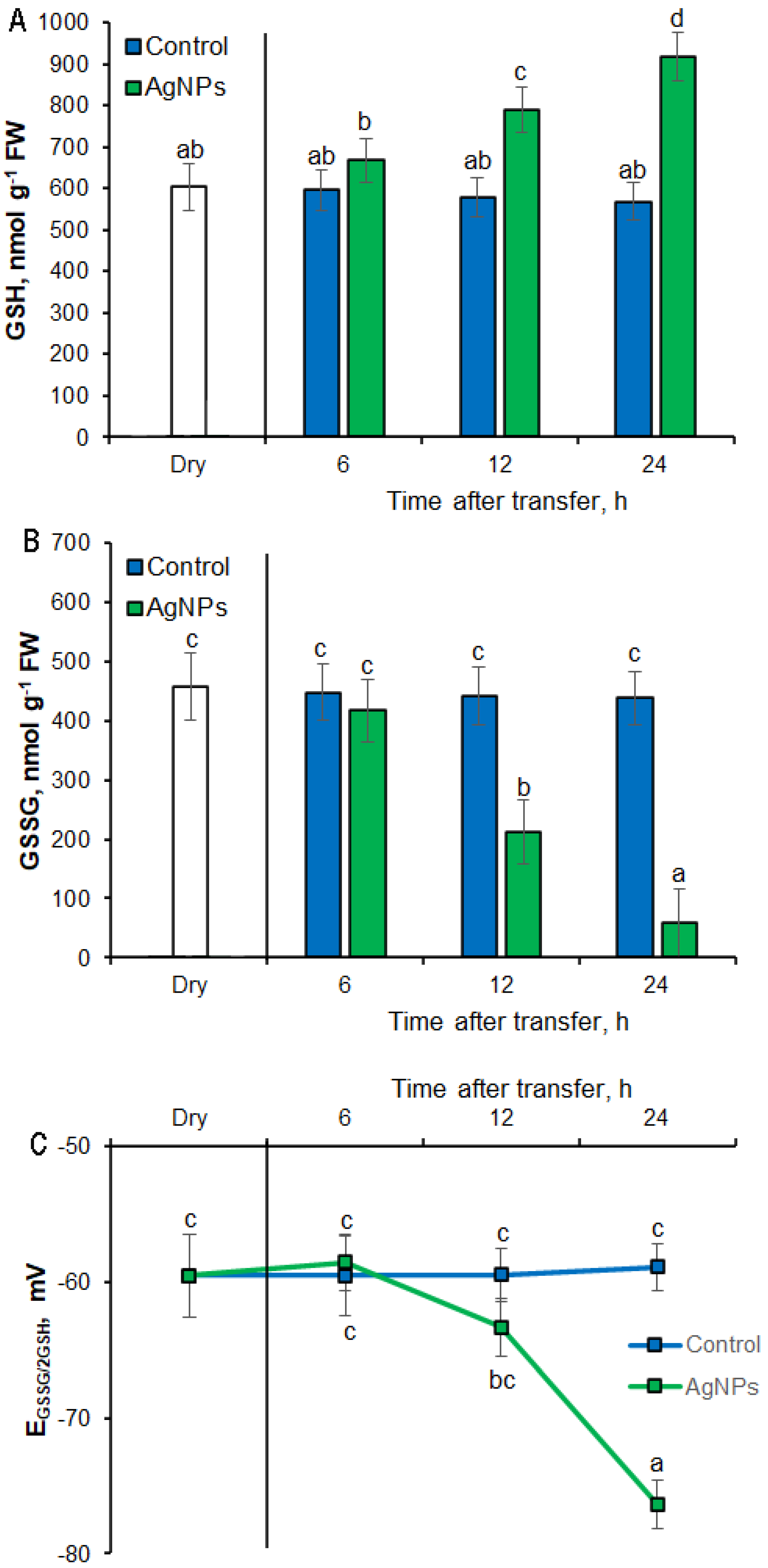
2.4. Effets of AgNPs on Glutathione Level and Half-Cell Redox Potential
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Ag Suspensions
4.3. Germination Assay and Treatment Experiments
4.4. Superoxide Anion and Hydrogen Peroxide Content
4.5. Enzymes Extraction and Assays
4.6. Protein Assay
4.7. Western Blot Analysis
4.8. Determination of Glutathione Content and Glutathione Half-Cell Reduction Potential
4.9. RNA Extraction, cDNA Synthesis and RT-qPCR
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy, 3rd ed.; Springer: New York, NY, USA, 2013. [Google Scholar]
- Liu, S.J.; Xu, H.H.; Wang, W.Q.; Li, N.; Wang, W.P.; Møller, I.M.; Song, S.Q. A proteomic analysis of rice seed germination as affected by high temperature and ABA treatment. Physiol. Plant. 2015, 154, 142–161. [Google Scholar] [CrossRef] [PubMed]
- Rengasamy, P. World salinization with emphasis on Australia. J. Exp. Bot. 2006, 57, 1017–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qadir, M.; Quillerou, E.; Nangia, V.; Murtaza, G.; Singh, M.; Thomas, R.J.; Drechsel, P.; Noble, A.D. Economics of salt-induced land degradation and restoration. Nat. Res. Forum 2014, 38, 282–295. [Google Scholar] [CrossRef]
- Rea, P.A.; Kim, Y.C.; Sarafian, V.; Poole, R.J.; Davies, J.M.; Sanders, D. Vacuolar H+-translocating pyrophosphatases—A new category of ion translocase. Trends Biochem. Sci. 1992, 17, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Horie, T.; Schroeder, J.I. Sodium transporters in plants. Diverse genes and physiological functions. Plant Physiol. 2004, 136, 2457–2462. [Google Scholar] [CrossRef] [Green Version]
- Bliss, R.D.; Plattaloia, K.A.; Thomson, W.W. Osmotic sensitivity in relation to salt sensitivity in germinating barley seeds. Plant Cell Environ. 1986, 9, 721–725. [Google Scholar] [CrossRef]
- Hampson, C.R.; Simpson, G.M. Effects of temperature, salt, and osmotic potential on early growth of wheat (Triticum aestivum). Can. J. Bot. 1990, 68, 524–528. [Google Scholar] [CrossRef]
- Huang, J.; Redmann, R.E. Salt tolerance of Hordeum and Brassica species during germination and early seedling growth. Can. J. Plant Sci. 1995, 75, 815–819. [Google Scholar] [CrossRef]
- Ibrahim, E.A. Seed Priming to Alleviate Salinity Stress in Germinating Seeds. J. Plant Physiol. 2016, 192, 38–46. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- El-Maarouf-Bouteau, H.; Bailly, C. Oxidative signaling in seed germination and dormancy. Plant Signal. Behav. 2008, 3, 175–182. [Google Scholar] [CrossRef] [Green Version]
- Oracz, K.; Karpiński, S. Phytohormones Signaling Pathways and ROS Involvement in Seed Germination. Front. Plant Sci. 2016, 15, 864. [Google Scholar] [CrossRef] [Green Version]
- Bailly, C.; Leymarie, J.; Lehner, A.; Rousseau, S.; Côme, D.; Corbineau, F. Catalase activity and expression in developing sunflower seeds as related to drying. J. Exp. Bot. 2004, 55, 475–483. [Google Scholar] [CrossRef] [Green Version]
- Gomes, M.P.; Garcia, Q.S. Reactive oxygen species and seed germination. Biologia 2013, 68, 351–357. [Google Scholar] [CrossRef]
- Debnath, B.; Sarkar, S.; Das, R. Effects of saponin capped triangular silver nanocrystals on the germination of Pisum sativum, Cicer arietinum, Vigna radiata seeds and their subsequent growth study. IET Nanobiotechnol. 2020, 14, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Szőllősi, R.; Molnár, Á.; Kondak, S.; Kolbert, Z. Dual effect of nanomaterials on germination and seedling growth: Stimulation vs. phytotoxicity. Plants 2020, 9, 1745. [Google Scholar] [CrossRef]
- Lin, D.; Xing, B. Phytotoxicity of Nanoparticles Inhibition of Seed Germination and Root Growth. Environ. Pollut. 2007, 50, 243–250. [Google Scholar] [CrossRef]
- Baik, B.K.; Ullrich, S.E. Barley for food: Characteristics, improvement, and renewed interest. J. Cereal Sci. 2008, 48, 233–242. [Google Scholar] [CrossRef]
- Seckin, B.; Türkan, I.; Sekmen, A.H.; Ozfidan, C. The role of antioxidant defense system at differential salt tolerance of Hordeum marinum Huds. (sea barley grass) and Hordeum vulgare L. (cultivated barley). Environ. Exp. Bot. 2010, 69, 76–85. [Google Scholar] [CrossRef]
- Hernández, J.A. Salinity tolerance in plants: Trends and perspectives. Int. J. Mol. Sci. 2019, 20, 2408. [Google Scholar] [CrossRef] [PubMed]
- El-Hendawy, S.; Elshafei, A.; Al-Suhaibani, N.; Alotabi, M.; Hassan, W.; Dewir, Y.H.; Abdella, K. Assessment of the salt tolerance of wheat genotypes during the germination stage based on germination ability parameters and associated SSR markers. J. Plant Interact. 2019, 14, 151–163. [Google Scholar] [CrossRef] [Green Version]
- Bakhshandeh, E.; Jamali, M. Population-based threshold models: A reliable tool for describing aged seeds response of rapeseed under salinity and water stress. Environ. Exp. Bot. 2020, 176, 104077. [Google Scholar] [CrossRef]
- Tavakkoli, E.; Rengasamy, P.; McDonald, G. High concentrations of Na+ and Cl- ions in soil solution have simultaneous detrimental effects on growth of faba bean under salinity stress. J. Exp. Bot. 2010, 61, 4449–4459. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Semida, W.M.; Rady, M.M.; Mohamed, G.F.; Hemida, K.A.; Alhammad, B.A.; Hassan, M.; Shami, A. Sequential application of antioxidants rectifies ion imbalance and strengthens antioxidant systems in salt-stressed cucumber. Plants 2020, 9, 1783. [Google Scholar] [CrossRef]
- Alkharabsheh, H.M.; Seleiman, M.F.; Hewedy, O.A.; Battaglia, M.L.; Jalal, R.S.; Alhammad, B.A.; Schillaci, C.; Ali, N.; Al-Doss, A. Field crop responses and management strategies to mitigate soil salinity in modern agriculture: A review. Agronomy 2021, 11, 2299. [Google Scholar] [CrossRef]
- Taha, R.S.; Seleiman, M.F.; Shami, A.; Alhammad, B.A.; Mahdi, A.H.A. Integrated application of selenium and silicon enhances growth and anatomical structure, antioxidant defense system and yield of wheat grown in salt-stressed soil. Plants 2021, 10, 1040. [Google Scholar] [CrossRef] [PubMed]
- El-Tayeb, M.A. Response of Barley Grains to the Interactive Effect of Salinity and Salicylic Acid. Plant Growth Regul. 2005, 45, 215–224. [Google Scholar] [CrossRef]
- Othman, Y.; Al-Karaki, G.; Al-Tawaha, A.R.; Al-Horani, A. Variation in germination and ion uptake in barley genotypes under salinity conditions. World J. Agric. Sci. 2006, 2, 11–15. [Google Scholar]
- Khan, T.A.; Mazid, M.; Mohammad, F. Status of Secondary Plant Products under Abiotic Stress: An Overview. J. Stress Physiol. Biochem. 2011, 7, 75–98. [Google Scholar]
- Baiazidi, T. Effects of Nanoparticulate Anatase Titanium Dioxide on Physiological and Biochemical Performance of Linum usitatissimum (Linaceae) under Well-Watered and Drought Stress Conditions. Rev. Bras. Bot. 2016, 39, 139–146. [Google Scholar]
- Marthandan, V.; Geetha, R.; Kumutha, K.; Renganathan, V.G.; Karthikeyan, A.; Ramalingam, J. Seed priming: A feasible strategy to enhance drought tolerance in crop plants. Int. J. Mol. Sci. 2020, 21, 8258. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.N.; Li, Y.; Khan, Z.; Chen, L.; Liu, J.; Hu, J.; Wu, H.; Li, Z. Nanoceria seed priming enhanced salt tolerance in rapeseed through modulating ROS homeostasis and α-amylase activities. J. Nanobiotechnol. 2021, 19, 276. [Google Scholar] [CrossRef]
- Acharya, P.; Jayaprakasha, G.K.; Crosby, K.M.; Jifon, J.L.; Patil, B.S. Green-synthesized nanoparticles enhanced seedling growth, yield, and quality of onion (Allium cepa L.). ACS Sustain. Chem. Eng. 2019, 7, 14580–14590. [Google Scholar] [CrossRef]
- Duran, N.M.; Savassa, S.M.; De Lima, R.G.; De Almeida, E.; Linhares, F.S.; Van Gestel, C.A.M.; De Carvalho, H.W.P. X-ray spectroscopy uncovering the effects of Cu based nanoparticle concentration and structure on Phaseolus vulgaris germination and seedling development. J. Agric. Food Chem. 2017, 65, 7874–7884. [Google Scholar] [CrossRef] [PubMed]
- Montanha, G.S.; Rodrigues, E.S.; Marques, J.P.R.; de Almeida, E.; Colzato, M.; de Carvalho, H.W.P. Zinc nanocoated seeds: An alternative to boost soybean seed germination and seedling development. SN Appl. Sci. 2020, 2, 857. [Google Scholar] [CrossRef] [Green Version]
- Xin, X.; Zhao, F.; Rho, J.Y.; Goodrich, S.L.; Sumerlin, B.S.; He, Z. Use of polymeric nanoparticles to improve seed germination and plant growth under copper stress. Sci. Total Environ. 2020, 745, 141055. [Google Scholar] [CrossRef]
- Tondey, M.; Kalia, A.; Singh, A.; Dheri, G.S.; Taggar, M.S.; Nepovimova, E.; Krejcar, O.; Kuca, K. Seed priming and coating by nano-scale zinc oxide particles improved vegetative growth, yield and quality of fodder maize (Zea mays). Agronomy 2021, 11, 729. [Google Scholar] [CrossRef]
- Sharifan, H.; Moore, J.; Ma, X. Zinc oxide (ZnO) nanoparticles elevated iron and copper contents and mitigated the bioavailability of lead and cadmium in different leafy greens. Ecotoxicol. Environ. Saf. 2020, 191, 110177. [Google Scholar] [CrossRef]
- Dimkpa, C.O.; McLean, J.E.; Latta, D.E.; Manangón, E.; Britt, D.W.; Johnson, W.P.; Boyanov, M.I.; Anderson, A.J. CuO and ZnO nanoparticles: Phytotoxicity, metal speciation, and induction of oxidative stress in sand-grown wheat. J. Nanopartic. Res. 2012, 14, 1125. [Google Scholar] [CrossRef]
- Boonyanitipong, P.; Kositsup, B.; Kumar, P.; Baruah, S.; Dutta, J. Toxicity of ZnO and TiO2 nanoparticles on germinating rice seed Oryza sativa L. Int. J. Biosci. Biochem. Bioinf. 2011, 1, 282–285. [Google Scholar] [CrossRef] [Green Version]
- Mahakham, W.; Sarmah, A.K.; Maensiri, S.; Theerakulpisut, P. Nanopriming technology for enhancing germination and starch metabolism of aged rice seeds using phytosynthesized silver nanoparticles. Sci. Rep. 2017, 7, 8263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abou-Zeid, H.M.; Ismail, G.S.M.; Abdel-Latif, S.A. Influence of Seed Priming with ZnO Nanoparticles on the Salt-Induced Damages in Wheat (Triticum aestivum L.). J. Plant Nutr. 2021, 44, 629–643. [Google Scholar] [CrossRef]
- An, J.; Hu, P.; Li, F.; Wu, H.; Shen, Y.; White, J.C.; Tian, X.; Li, Z.; Giraldo, J.P. Emerging investigator series: Molecular mechanisms of plant salinity stress tolerance improvement by seed priming with cerium oxide nanoparticles. Environ. Sci. Nano 2020, 7, 2214–2228. [Google Scholar] [CrossRef]
- Ye, Y.; Cota-Ruiz, K.; Hernández-Viezcas, J.A.; Valdés, C.; Medina-Velo, I.A.; Turley, R.S.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Manganese nanoparticles control salinity-modulated molecular responses in Capsicum annuum L. through priming: A sustainable approach for agriculture. ACS Sustain. Chem. Eng. 2020, 8, 1427–1436. [Google Scholar] [CrossRef]
- Sonawane, H.; Arya, S.; Math, S.; Shelke, D. Myco-Synthesized Silver and Titanium Oxide Nanoparticles as Seed Priming Agents to Promote Seed Germination and Seedling Growth of Solanum Lycopersicum: A Comparative Study. Int. Nano Lett. 2021, 11, 371–379. [Google Scholar] [CrossRef]
- Shah, T.; Latif, S.; Saeed, F.; Ali, I.; Ullah, S.; Abdullah Alsahli, A.; Jan, S.; Ahmad, P. Seed Priming with Titanium Dioxide Nanoparticles Enhances Seed Vigor, Leaf Water Status, and Antioxidant Enzyme Activities in Maize (Zea mays L.) under Salinity Stress. J. King Saud. Univ. Sci. 2021, 33, 101207. [Google Scholar] [CrossRef]
- Kumar, V.K.; Muthukrishnan, S.; Rajalakshmi, R. Phytostimulatory Effect of Phytochemical Fabricated Nanosilver (AgNPs) on Psophocarpus tetragonolobus (L.) DC. Seed Germination: An Insight from Antioxidative Enzyme Activities and Genetic Similarity Studies. Curr. Plant Biol. 2020, 23, 100158. [Google Scholar] [CrossRef]
- Ishibashi, Y.; Tawaratsumida, T.; Zheng, S.H.; Yuasa, T.; Iwaya-Inoue, M. NADPH Oxidases Act as Key Enzyme on Germination and Seedling Growth in Barley (Hordeum vulgare L.). Plant Prod. Sci. 2010, 13, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Cembrowska-Lech, D. Tissue Printing and Dual Excitation Flow Cytometry for Oxidative Stress-New Tools for Reactive Oxygen Species Research in Seed Biology. Int. J. Mol. Sci. 2020, 21, 8656. [Google Scholar] [CrossRef]
- Beyer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences for minor changes in conditions. Annu. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Thordal-Christensen, H.; Zhang, Z.; Wei, Y.; Collinge, D.B. Subcellular localization of H2O2 in plants: H2O2 accumulation in papillae and hypersensitive response during the barley powdery mildew interaction. Plant J. 1997, 11, 1187–1194. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.V.; Paloyath, G.; Ormrod, D.P. Ultraviolet-B- and ozone-induced biochemical changes in antioxidant enzymes of Arabidopsis thaliana. Plant Physiol. 1996, 110, 125–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esterbauer, H.; Grill, D. Seasonal variation of glutathione and glutathione reductase in needles of Picea abies. Plant Physiol. 1978, 61, 119–121. [Google Scholar] [CrossRef] [Green Version]
- Nagalakshmi, N.; Prasad, M.N. Responses of glutathione cycle enzymes and glutathione metabolism to copper stress in Scenedesmus bijugatus. Plant Sci. 2001, 160, 291–299. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities utilizing the principle of protein dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head phase of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Smith, I. Stimulation of glutathione synthesis in photorespiring plants by catalase inhibitors. Plant Physiol. 1985, 79, 1044–1047. [Google Scholar] [CrossRef] [Green Version]
- Schafer, F.Q.; Buettner, G.R. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. Free Radic. Bio. Med. 2001, 30, 1191–1212. [Google Scholar] [CrossRef]
- Oñate-Sánchez, L.; Vincente-Carbajosa, J. DNA-free RNA isolation protocols for Arabidopsis thaliana, including seeds and siliques. BMC Res. Notes 2008, 1, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trevaskis, B.; Hemming, M.N.; Peacock, W.J.; Dennis, E.J. HvVRN2 responds to daylength, whereas HvVRN1 is regulated by vernalization and developmental status. Plant Physiol. 2006, 140, 1397–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, 2002–2007. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cembrowska-Lech, D.; Rybak, K. Nanopriming of Barley Seeds—A Shotgun Approach to Improve Germination under Salt Stress Conditions by Regulating of Reactive Oxygen Species. Plants 2023, 12, 405. https://doi.org/10.3390/plants12020405
Cembrowska-Lech D, Rybak K. Nanopriming of Barley Seeds—A Shotgun Approach to Improve Germination under Salt Stress Conditions by Regulating of Reactive Oxygen Species. Plants. 2023; 12(2):405. https://doi.org/10.3390/plants12020405
Chicago/Turabian StyleCembrowska-Lech, Danuta, and Kinga Rybak. 2023. "Nanopriming of Barley Seeds—A Shotgun Approach to Improve Germination under Salt Stress Conditions by Regulating of Reactive Oxygen Species" Plants 12, no. 2: 405. https://doi.org/10.3390/plants12020405
APA StyleCembrowska-Lech, D., & Rybak, K. (2023). Nanopriming of Barley Seeds—A Shotgun Approach to Improve Germination under Salt Stress Conditions by Regulating of Reactive Oxygen Species. Plants, 12(2), 405. https://doi.org/10.3390/plants12020405