
Effects of Thinning Intensity on Litterfall Production, Soil Chemical Properties, and Fine Root Distribution in Pinus koraiensis Plantation in Republic of Korea

 , ,
, ,  , , and
, , and 
Abstract
:1. Introduction
2. Results
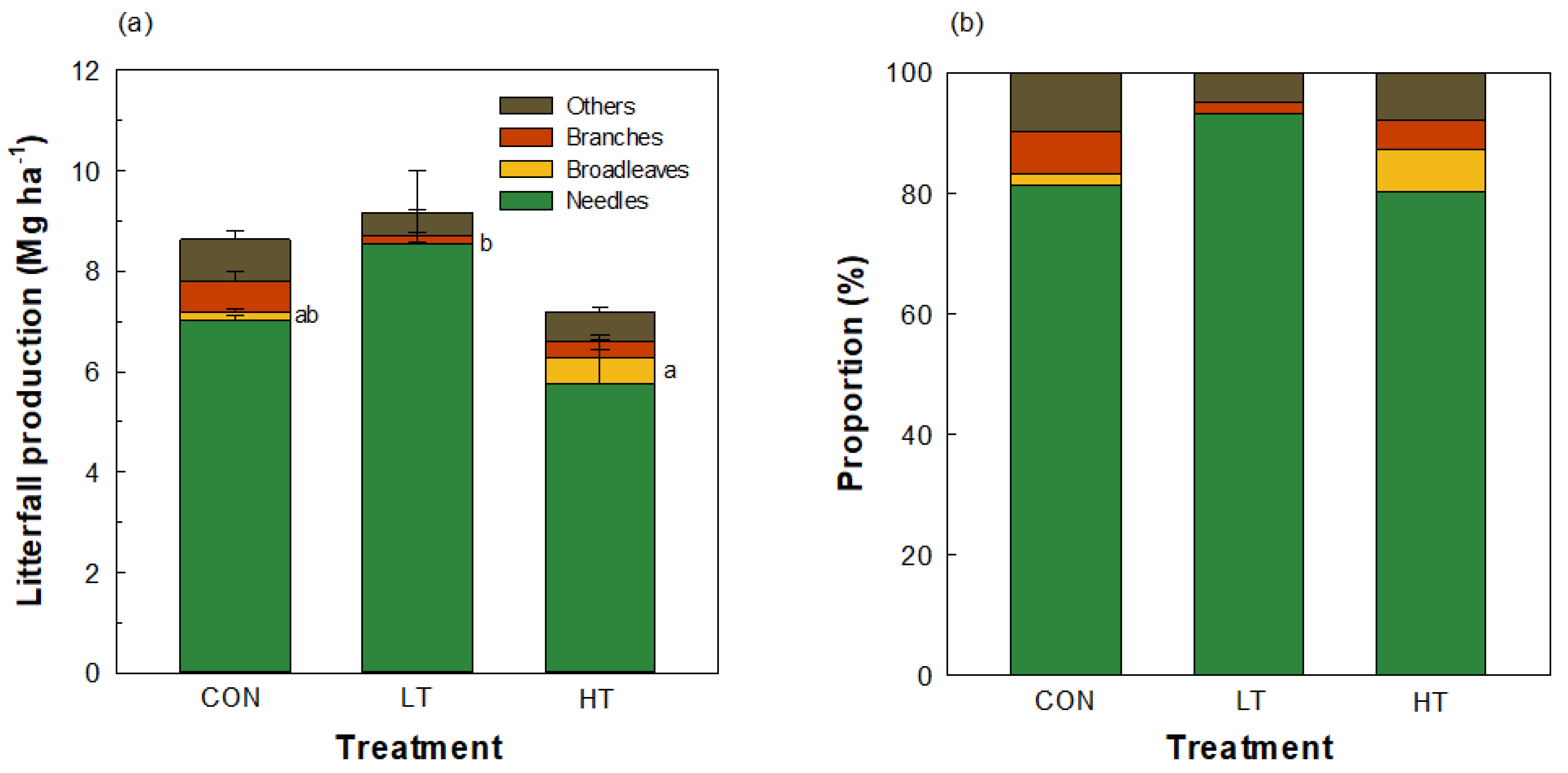
2.1. Litterfall Production by Thinning Intensity
2.2. Thinning-Induced Changes in Soil Chemical Properties
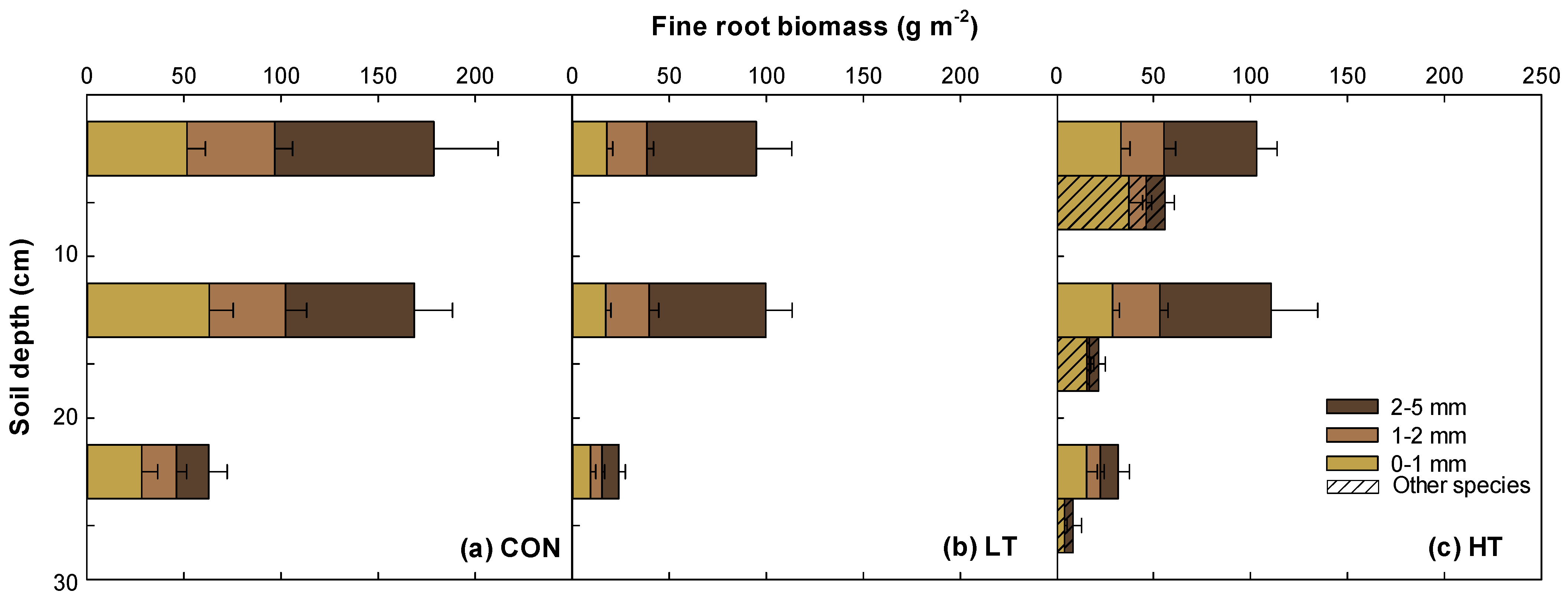
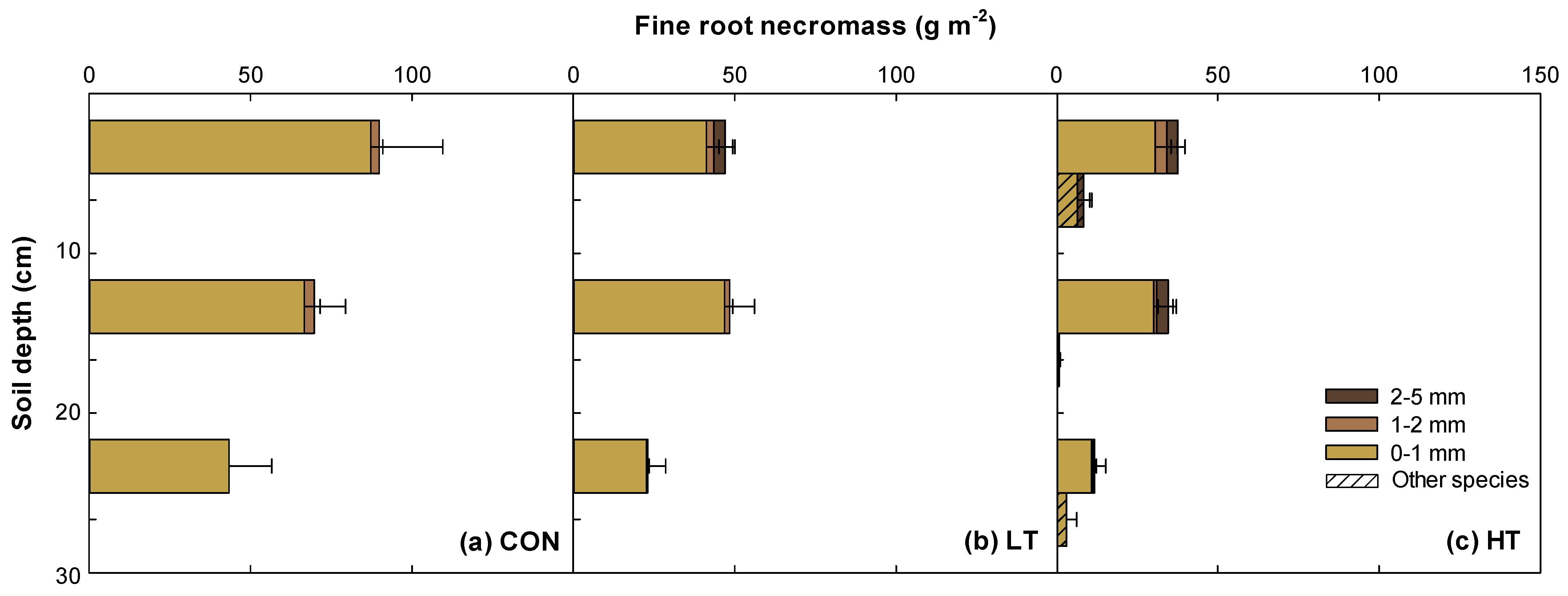
2.3. Vertical Distribution of Fine Roots in Different Thinning Intensity Treatments
3. Discussion
3.1. Significant Increase in Broadleaf Litterfall Production in a Pinus koraiensis Plantation Ten Years after Thinning
3.2. Heavy Thinning Intensity Increased Abundance of Other Species’ Roots in P. koraiensis Plantation
3.3. Vertical Distribution Patterns of Fine Root Biomass and Necromass in P. koraiensis Plantation
3.4. Ecological Implications
4. Materials and Methods
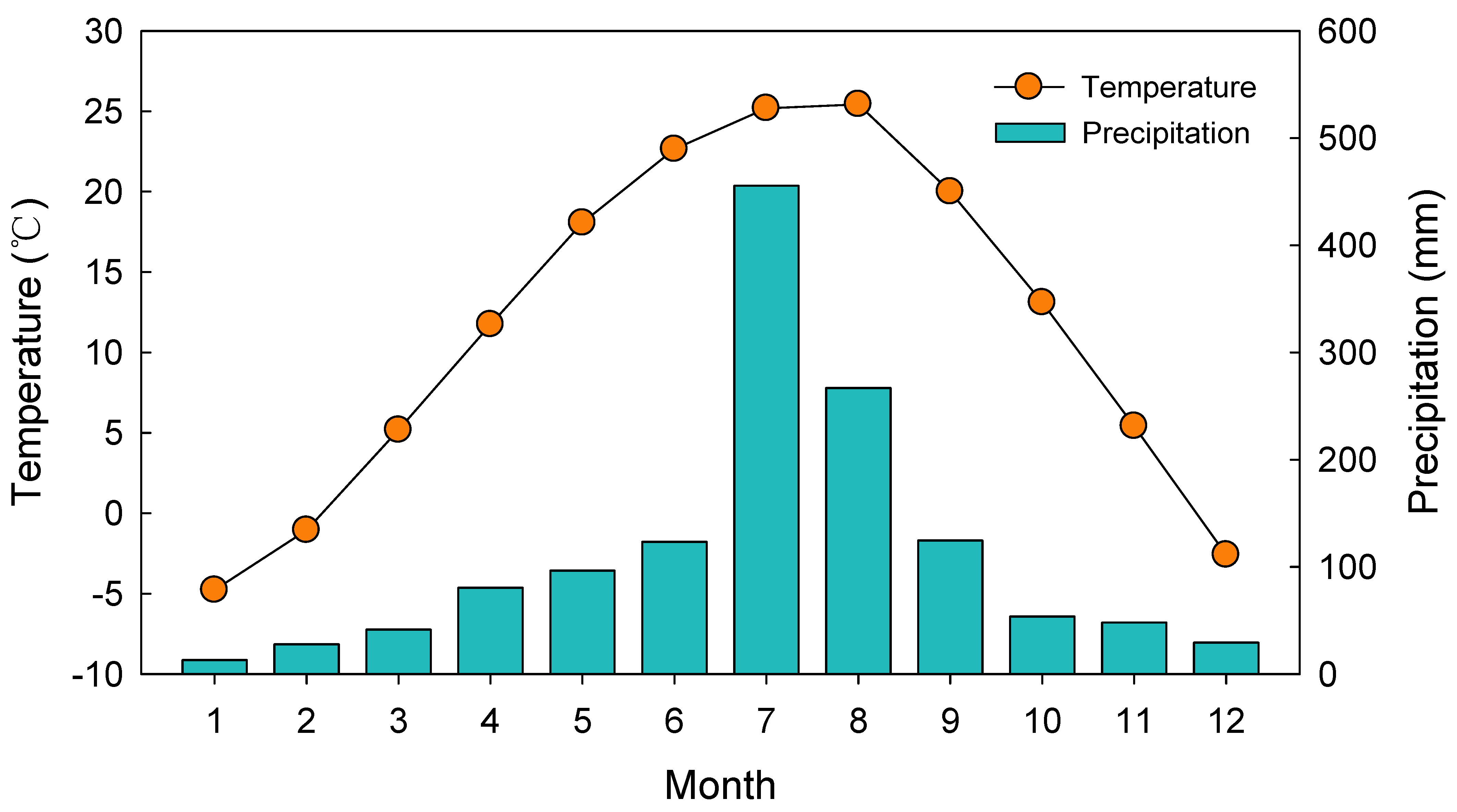
4.1. Site Descriptions
4.2. Estimation of Annual Litterfall Production
4.3. Soil Sampling and Analyses
4.4. Measurement of Vertical Distribution of Fine Roots
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collalti, A.; Trotta, C.; Keenan, T.F.; Ibrom, A.; Bond-Lamberty, B.; Grote, R.; Vicca, S.; Reyer, C.P.; Migliavacca, M.; Veroustraete, F.; et al. Thinning can reduce losses in carbon use efficiency and carbon stocks in managed forests under warmer climate. J. Adv. Model. Earth Syst. 2018, 10, 2427–2452. [Google Scholar] [CrossRef] [PubMed]
- Sankey, T.; Tatum, J. Thinning increases forest resiliency during unprecedented drought. Sci. Rep. 2022, 12, 9041. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, M.; Li, G.; Wang, M.; Li, Z.; De Boeck, H.J. Changes of aboveground and belowground biomass allocation in four dominant grassland species across a precipitation gradient. Front. Plant Sci. 2021, 12, 650802. [Google Scholar] [CrossRef]
- Meena, A.; Bidalia, A.; Hanief, M.; Dinakaran, J.; Rao, K.S. Assessment of above- and belowground carbon pools in a semi-arid forest ecosystem of Delhi, India. Ecol. Process 2019, 8, 8. [Google Scholar] [CrossRef]
- Yang, H.; Miao, N.; Li, S.; Ma, R.; Liao, Z.; Wang, W.; Sun, H. Relationship between stand characteristics and soil properties of two typical forest plantations in the mountainous area of western Sichuan, China. J. Mt. Sci. 2019, 16, 1816–1832. [Google Scholar] [CrossRef]
- McCormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.; Helmisaari, H.S.; Hobbie, E.A.; Iversen, C.M.; Jackson, R.B. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 2015, 207, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Wang, N.; Cheng, R.; Xiao, W.; Yang, S.; Guo, Y.; Lei, L.; Zeng, L.; Wang, X. Characteristics of fine roots of Pinus massoniana in the Three Gorges Reservoir Area, China. Forests 2017, 8, 183. [Google Scholar] [CrossRef]
- Pang, Y.; Tian, J.; Yang, H.; Zhang, K.; Wang, D. Responses of fine roots at different soil depths to different thinning intensities in a secondary forest in the Qinling Mountains, China. Biology 2022, 11, 351. [Google Scholar] [CrossRef]
- Dawoe, E.K.; Isaac, M.E.; Quashie-Sam, J. Litterfall and litter nutrient dynamics under cocoa ecosystems in lowland humid Ghana. Plant Soil 2009, 330, 55–64. [Google Scholar] [CrossRef]
- Wei, X.; Yang, Y.; Shen, Y.; Chen, Z.; Dong, Y.; Wu, F.; Zhang, L. Effects of litterfall on the accumulation of extracted soil humic substances in subalpine forests. Front. Plant Sci. 2020, 11, 254. [Google Scholar] [CrossRef]
- Doukalianou, F.; Spyroglou, G.; Orfanoudakis, M.; Radoglou, K.; Stefanou, S.; Kitikidou, K.; Milios, E.; Lagomarsino, A. Effects of forest thinning on soil litter input nutrients in relation to soil CO2, CH4, and N2O fluxes in Greece. Atmosphere 2022, 13, 376. [Google Scholar] [CrossRef]
- Navarro, F.B.; Romero-Freire, A.; del Castillo, T.; Foronda, A.; Jiménez, M.N.; Ripoll, M.A.; Sánchez-Miranda, A.; Huntsingerd, L.; Fernández-Ondoño, E. Effects of thinning on litterfall were found after years in a Pinus halepensis afforestation area at tree and stand levels. For. Ecol. Manag. 2013, 289, 354–362. [Google Scholar] [CrossRef]
- Bueis, T.; Bravo, F.; Pando, V.; Turrión, M.B. Local basal area affects needle litterfall, nutrient concentration, and nutrient release during decomposition in Pinus halepensis Mill. plantations in Spain. Ann. For. Sci. 2018, 75, 21. [Google Scholar] [CrossRef]
- Teste, F.P.; Lieffers, V.J.; Strelkov, S.E. Ectomycorrhizal community responses to intensive forest management: Thinning alters impacts of fertilization. Plant Soil 2012, 360, 333–347. [Google Scholar] [CrossRef]
- Park, B.B.; Lee, I.K.; Yang, H.M. The effects of thinning on fine root distribution and litterfall in a Pinus koraiensis plantation. Ecol. Field. Biol. 2009, 32, 159–165. [Google Scholar] [CrossRef]
- Stefanowicz, A.M.; Kapusta, P.; Stanek, M.; Rola, K.; Zubek, S. Herbaceous plant species support soil microbial performance in deciduous temperate forests. Sci. Total Environ. 2022, 810, 151313. [Google Scholar] [CrossRef]
- Molina, A.J.; Bautista, I.; Lull, C.; del Campo, A.; González-Sanchis, M.; Lidón, A. effects of thinning intensity on forest floor and soil biochemical properties in an Aleppo pine plantation after 13 Years: Quantity but also quality matters. Forests 2022, 13, 255. [Google Scholar] [CrossRef]
- Lado-Monserrat, L.; Lidón, A.; Bautista, I. Erratum to: Litterfall, litter decomposition and associated nutrient fluxes in Pinus halepensis: Influence of tree removal intensity in a Mediterranean forest. Eur. J. For. Res. 2016, 135, 203–214. [Google Scholar] [CrossRef]
- James, J.; Harrison, R. The effect of harvest on forest soil carbon: A meta-analysis. Forests 2016, 7, 308. [Google Scholar] [CrossRef]
- Aussenac, G. Interactions between forest stands and microclimate: Ecophysiological aspects and consequences for silviculture. Ann. For. Sci. 2000, 57, 287–301. [Google Scholar] [CrossRef]
- Wang, D.; Olatunji, O.A.; Xiao, J. Thinning increased fine root production, biomass, turnover rate and understory vegetation yield in a Chinese fir plantation. For. Ecol. Manag. 2019, 440, 92–100. [Google Scholar] [CrossRef]
- Ruiz-Peinado, R.; Bravo-Oviedo, A.; López-Senespleda, E.; Montero, G.; Río, M. Do thinnings influence biomass and soil carbon stocks in Mediterranean maritime pinewoods? Eur. J. For. Res. 2013, 132, 253–262. [Google Scholar] [CrossRef]
- Nave, L.E.; Vance, E.D.; Swanston, C.W.; Curtis, P.S. Harvest impacts on soil carbon storage in temperate forests. For. Ecol. Manag. 2010, 259, 857–866. [Google Scholar] [CrossRef]
- Son, Y.H.; Noh, N.J.; Kim, R.H.; Koo, J.W.; Yi, M.J. Biomass and nutrients of planted and naturally occurring Pinus koraiensis in Korea. Eurasian J. For. Res. 2007, 10, 41–50. [Google Scholar]
- Korea Forest Service. 2020 Statistical Yearbook of Forestry; Korea Forest Service: Daejeon, Republic of Korea, 2020; p. 448. (In Korean) [Google Scholar]
- Dong, Y.; Liu, Y. Response of Korean pine’s functional traits to geography and climate. PLoS ONE 2017, 12, e0184051. [Google Scholar] [CrossRef] [PubMed]
- Hale, S.E. The effect of thinning intensity on the below-canopy light environment in a sitka spruce plantation. For. Ecol. Manag. 2003, 179, 341–349. [Google Scholar] [CrossRef]
- Giweta, M. Role of litter production and its decomposition, and factors affecting the processes in a tropical forest ecosystem: A Review. J. Ecol. Environ. 2020, 44, 11. [Google Scholar] [CrossRef]
- Umaña, M.N.; Arellano, G.; Swenson, N.G.; Zambrano, J. Tree seedling trait optimization and growth in response to local-scale soil and light variability. Ecology 2021, 102, e03252. [Google Scholar] [CrossRef]
- Song, X.; Yang, J.; Cao, M.; Lin, L.; Sun, Z.; Wen, H.; Swenson, N.G. Traits mediate a trade-off in seedling growth response to light and conspecific density in a diverse subtropical forest. J. Ecol. 2021, 109, 703–713. [Google Scholar] [CrossRef]
- Seiwa, K.; Eto, Y.; Hishita, M.; Masaka, K. Effects of thinning intensity on species diversity and timber production in a conifer (Cryptomeria japonica) plantation in Japan. J. For. Res. 2012, 17, 468–478. [Google Scholar] [CrossRef]
- Yu, J.; Zhang, X.; Xu, C.; Hao, M.; Choe, C.; He, H. Thinning can increase shrub diversity and decrease herb diversity by regulating light and soil environments. Front. Plant Sci. 2022, 13, 948648. [Google Scholar] [CrossRef]
- Willms, J.; Bartuszevige, A.; Schwilk, D.W.; Kennedy, P.L. The effects of thinning and burning on understory vegetation in North America: A meta-analysis. For. Ecol. Manag. 2017, 392, 184–194. [Google Scholar] [CrossRef]
- Thrippleton, T.; Bugmann, H.; Folini, M.; Snell, R.S. Overstorey–understorey interactions intensify after drought induced forest die-off: Long-term effects for forest structure and composition. Ecosystems 2018, 21, 723–739. [Google Scholar] [CrossRef]
- Lyu, S.; Wang, X.; Zhang, Y.; Li, Z. Different responses of Korean pine (Pinus koraiensis) and Mongolia Oak (Quercus mongolica) growth to recent climate warming in Northeast China. Dendrochronologia 2017, 45, 113–122. [Google Scholar] [CrossRef]
- Yacht, V.A.L.; Kobe, R.K.; Walters, M.B. Hardwood regeneration in red pine plantations: Thinning and site quality effects on changes in density and species composition from edges to interiors. For. Ecol. Manag. 2022, 526, 120576. [Google Scholar] [CrossRef]
- Hartman, J.P.; Buckley, D.S.; Sharik, T.L. Differential success of oak and red maple regeneration in oak and pine stands on intermediate-quality sites in northern Lower Michigan. For. Ecol. Manag. 2005, 216, 77–90. [Google Scholar] [CrossRef]
- Granger, J.J.; Buckley, D.S.; Sharik, T.L.; Zobel, J.M.; DeBord, W.W.; Hartman, J.P.; Henning, J.G.; Keyser, T.L.; Marshall, J.M. Northern red oak regeneration: 25-year results of cutting and prescribed fire in Michigan oak and pine stands. For. Ecol. Manag. 2018, 429, 467–479. [Google Scholar] [CrossRef]
- Zhou, L.B.; Yan, X.F.; Wang, J.L.; Zhou, Y.F. Predation and removal of rodents on the seeds with different size and pericarp traits. Chin. J. Appl. Ecol. 2013, 24, 2325–2332. [Google Scholar]
- Yu, F.; Shi, X.X.; Wang, D.X.; Yi, X.F.; Fan, D.F.; Guo, T.D.; Lou, Y.G. Effects of insect infestation on Quercus aliena var. acuteserrata acorn dispersal in the Qinling Mountains, China. New For. 2015, 46, 51–61. [Google Scholar]
- Kang, H.; Chang, M.; Liu, S.; Chao, Z.; Zhang, X.; Wang, D. Rodent-mediated plant community competition: What happens to the seeds after entering the adjacent stands? For. Ecosyst. 2020, 7, 56. [Google Scholar] [CrossRef]
- Ganbaatar, B.; Jamsran, T.; Gradel, A.; Sukhbaatar, G. Assessment of the effects of thinnings in Scots pine plantations in Mongolia: A comparative analysis of tree growth and crown development based on dominant trees. For. Sci. Technol. 2021, 17, 135–143. [Google Scholar] [CrossRef]
- Prescott, C.E.; Zabek, L.M.; Staley, C.L.; Kabzems, R. Decomposition of broadleaf and needle litter in forests of British Columbia: Influences of litter type, forest type, and litter mixtures. Can. J. For. Res. 2000, 30, 1742–1750. [Google Scholar] [CrossRef]
- Chen, F.L.; Jiang, B.; Zhang, K.; Zheng, H.; Xiao, Y.; Ouyang, Z.Y.; Tu, N.M. Relationships between initial chemical composition of forest leaf litters and their decomposition rates in degraded red soil hilly region of Southern China. Chin. J. Appl. Ecol. 2011, 22, 565–570. [Google Scholar]
- Pitkänen, T.P.; Bianchi, S.; Kangas, A. Quantifying the effects of competition on the dimensions of Scots Pine and norway spruce crowns. Int. J. Appl. Earth Obs. Geoinf. 2022, 112, 102941. [Google Scholar] [CrossRef]
- Ara, M.; Pinno, B.D.; Scaria, F.; Froese, R.E.; Bokalo, M. Thinning increases individual tree growth while reducing the growth heterogeneity of Lodgepole Pine. Forests 2023, 14, 1091. [Google Scholar] [CrossRef]
- Shu, W.; Shen, X.; Lei, P.; Xiang, W.; Ouyang, S.; Yan, W. Temporal changes of fine root overyielding and foraging strategies in planted monoculture and mixed forests. BMC Ecol. 2018, 18, 9. [Google Scholar] [CrossRef]
- Wang, W.; Wu, X.; Hu, K.; Liu, J.; Tao, J. Understorey fine root mass and morphology in the litter and upper soil layers of three Chinese subtropical forests. Plant Soil 2016, 406, 219–230. [Google Scholar] [CrossRef]
- Du, H.; Liu, L.; Su, L.; Zeng, F.; Wang, K.; Peng, W.; Zhang, H.; Song, T. Seasonal changes and vertical distribution of fine root biomass during vegetation restoration in a karst area, Southwest China. Front. Plant Sci. 2019, 9, 2001. [Google Scholar] [CrossRef]
- Zhou, H.; Zhao, W.; Yang, Q. Root biomass distribution of planted Haloxylon ammodendron in a duplex soil in an OASIS: Desert Boundary Area. Ecol. Res. 2016, 31, 673–681. [Google Scholar] [CrossRef]
- Pandey, R.; Bargali, S.S.; Bargali, K.; Karki, H.; Kumar, M.; Sahoo, U.K. Fine root dynamics and associated nutrient flux in Sal dominated forest ecosystems of Central Himalaya, India. Front. For. Glob. Change 2023, 5, 1064502. [Google Scholar] [CrossRef]
- Montagnoli, A.; Terzaghi, M.; Di Iorio, A.; Scippa, G.S.; Chiatante, D. Fine-root morphological and growth traits in a Turkey-oak stand in relation to seasonal changes in soil moisture in the Southern Apennines, Italy. Ecol. Res. 2012, 27, 1015–1025. [Google Scholar] [CrossRef]
- Sun, M.; Zhai, B.-C.; Chen, Q.-W.; Li, G.; Du, S. Response of density-related fine root production to soil and leaf traits in coniferous and broad-leaved plantations in the semiarid loess hilly region of China. J. For. Res. 2021, 33, 1071–1082. [Google Scholar] [CrossRef]
- Chen, L.; Mu, X.; Yuan, Z.; Deng, Q.; Chen, Y.; Yuan, L.Y.; Ryan, L.T.; Kallenbach, R.L. Soil nutrients and water affect the age-related fine root biomass but not production in two plantation forests on the Loess Plateau, China. J. Arid Environ. 2016, 135, 173–180. [Google Scholar] [CrossRef]
- Msimbira, L.A.; Smith, D.L. The roles of plant growth promoting microbes in enhancing plant tolerance to acidity and alkalinity stresses. Front. Sustain. Food Syst. 2020, 4, 106. [Google Scholar] [CrossRef]
- Mori, A.S. Environmental controls on the causes and functional consequences of tree species diversity. J. Ecol. 2017, 106, 113–125. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Barbaro, L.; Castagneyrol, B.; Forrester, D.I.; Gardiner, B.; González-Olabarria, J.R.; Lyver, P.O.; Meurisse, N.; Oxbrough, A.; Taki, H.; et al. Forest Biodiversity, ecosystem functioning and the provision of ecosystem services. Biodivers. Conserv. 2017, 26, 3005–3035. [Google Scholar] [CrossRef]
- Huang, Y.; Ma, K.; Niklaus, P.A.; Schmid, B. Leaf-litter overyielding in a forest biodiversity experiment in subtropical China. For. Ecosyst. 2018, 5, 38. [Google Scholar] [CrossRef]
- Meng, Y.; Zhang, Y.; Li, C.; Wang, Z.; Li, Y. The effect of thinning management on the carbon density of the tree layers in larch–birch mixed natural secondary forests of the Greater Khingan Range, northeastern China. Forests 2022, 13, 1035. [Google Scholar] [CrossRef]
- Sohng, J.; Han, A.; Jeong, M.-A.; Park, Y.; Park, B.; Park, P. Seasonal pattern of decomposition and N, P, and C dynamics in leaf litter in a Mongolian oak forest and a korean pine plantation. Forests 2014, 5, 2561–2580. [Google Scholar] [CrossRef]
- Neina, D. The role of soil ph in plant nutrition and soil remediation. Appl. Environ. Soil Sci. 2019, 2019, 5794869. [Google Scholar] [CrossRef]
- Wambsganss, J.; Beyer, F.; Freschet, G.T.; Scherer-Lorenzen, M.; Bauhus, J. Tree species mixing reduces biomass but increases length of absorptive fine roots in European forests. J. Ecol. 2021, 109, 2678–2691. [Google Scholar] [CrossRef]
- Hernandez, J.O.; An, J.Y.; Combalicer, M.S.; Chun, J.-P.; Oh, S.-K.; Park, B.B. Morpho-anatomical traits and soluble sugar concentration largely explain the responses of three deciduous tree species to progressive water stress. Front. Plant Sci. 2021, 12, 738301. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Depth | Treatment | pH | EC | OM | TN | AP | CEC | Exchangeable Cations (cmolc kg−1) | ||
|---|---|---|---|---|---|---|---|---|---|---|
| (cm) | (dS m−1) | (%) | (%) | (mg kg−1) | (cmolc kg−1) | K+ | Ca2+ | Mg2+ | ||
| 0–10 | CON | 4.95 | 0.49 | 5.93 | 0.29 | 29.30 | 10.83 | 0.19 | 3.14 | 2.05 |
| (0.06) | (0.02) | (1.01) | (0.02) | (5.73) | (0.63) | (0.005) | (0.51) | (0.09) | ||
| LT | 4.97 | 0.28 | 4.09 | 0.26 | 9.86 | 8.19 | 0.19 | 1.27 | 1.62 | |
| (0.08) | (0.03) | (0.28) | (0.01) | (3.50) | (0.27) | (0.003) | (0.42) | (0.12) | ||
| HT | 5.25 | 0.58 | 5.44 | 0.34 | 22.92 | 10.82 | 0.19 | 3.95 | 2.27 | |
| (0.05) | (0.08) | (0.41) | (0.01) | (3.25) | (0.63) | (0.006) | (0.47) | (0.10) | ||
| 10–20 | CON | 4.88 | 0.34 | 2.77 | 0.13 | 8.98 | 8.83 | 0.19 | 1.71 | 1.77 |
| (0.11) | (0.03) | (0.56) | (0.06) | (1.67) | (0.47) | (0.003) | (0.31) | (0.07) | ||
| LT | 4.85 | 0.22 | 1.91 | 0.05 | 4.85 | 7.05 | 0.18 | 0.51 | 1.41 | |
| (0.08) | (0.02) | (0.09) | (0.00) | (1.71) | (0.24) | (0.00) | (0.25) | (0.06) | ||
| HT | 5.23 | 0.40 | 3.45 | 0.28 | 12.27 | 8.79 | 0.19 | 2.47 | 1.96 | |
| (0.04) | (0.04) | (0.07) | (0.00) | (1.63) | (0.40) | (0.003) | (0.08) | (0.12) | ||
| 20–30 | CON | 4.84 | 0.30 | 1.97 | 0.08 | 12.70 | 8.06 | 0.18 | 1.24 | 1.64 |
| (0.08) | (0.03) | (0.34) | (0.04) | (6.47) | (0.16) | (0.00) | (0.23) | (0.07) | ||
| LT | 4.84 | 0.19 | 1.47 | 0.04 | 4.61 | 7.38 | 0.18 | 0.62 | 1.47 | |
| (0.08) | (0.01) | (0.16) | (0.01) | (0.84) | (0.38) | (0.00) | (0.35) | (0.08) | ||
| HT | 5.16 | 0.31 | 2.30 | 0.24 | 7.57 | 8.30 | 0.18 | 2.22 | 1.89 | |
| (0.02) | (0.03) | (0.22) | (0.00) | (1.52) | (0.62) | (0.00) | (0.19) | (0.13) | ||
| p-values | ||||||||||
| Treatment | <0.0001 | <0.0001 | 0.004 | <0.0001 | 0.003 | <0.0001 | 0.33 | <0.0001 | <0.0001 | |
| Soil depth | 0.19 | <0.0001 | <0.0001 | <0.0001 | 0.0002 | <0.0001 | 0.001 | <0.0001 | 0.001 | |
| T × D | 0.98 | 0.23 | 0.53 | 0.08 | 0.20 | 0.27 | 0.88 | 0.40 | 0.66 | |
| Before Thinning | 10 Years after Thinning | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CON | LT | HT | CON | LT | HT | ||||||||
| Stem density | (trees ha−1) | 538 | 588 | 550 | 500 | 363 | 188 | ||||||
| DBH | (cm) | 24.2 | (0.5) | 22.3 | (0.6) | 21.9 | (0.4) | 31.5 | (0.7) | 32.5 | (0.7) | 36.0 | (0.8) |
| Height | (m) | 15.1 | (0.3) | 13.4 | (0.2) | 14.5 | (0.2) | 19.4 | (0.2) | 18.3 | (0.2) | 17.4 | (0.2) |
| Basal area | (m2 ha−1) | 24.7 | 23.0 | 20.7 | 39.7 | 30.5 | 19.2 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, S.H.; An, J.Y.; Hernandez, J.O.; Yang, H.M.; Kim, E.-S.; Noh, N.J.; Seo, J.M.; Park, B.B. Effects of Thinning Intensity on Litterfall Production, Soil Chemical Properties, and Fine Root Distribution in Pinus koraiensis Plantation in Republic of Korea. Plants 2023, 12, 3614. https://doi.org/10.3390/plants12203614
Han SH, An JY, Hernandez JO, Yang HM, Kim E-S, Noh NJ, Seo JM, Park BB. Effects of Thinning Intensity on Litterfall Production, Soil Chemical Properties, and Fine Root Distribution in Pinus koraiensis Plantation in Republic of Korea. Plants. 2023; 12(20):3614. https://doi.org/10.3390/plants12203614
Chicago/Turabian StyleHan, Si Ho, Ji Young An, Jonathan O. Hernandez, Hee Moon Yang, Eun-Sook Kim, Nam Jin Noh, Jeong Min Seo, and Byung Bae Park. 2023. "Effects of Thinning Intensity on Litterfall Production, Soil Chemical Properties, and Fine Root Distribution in Pinus koraiensis Plantation in Republic of Korea" Plants 12, no. 20: 3614. https://doi.org/10.3390/plants12203614
APA StyleHan, S. H., An, J. Y., Hernandez, J. O., Yang, H. M., Kim, E. -S., Noh, N. J., Seo, J. M., & Park, B. B. (2023). Effects of Thinning Intensity on Litterfall Production, Soil Chemical Properties, and Fine Root Distribution in Pinus koraiensis Plantation in Republic of Korea. Plants, 12(20), 3614. https://doi.org/10.3390/plants12203614