
The Content of Anthocyanins in Cowpea (Vigna unguiculata (L.) Walp.) Seeds and Contribution of the MYB Gene Cluster to Their Coloration Pattern

 ,
,  ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
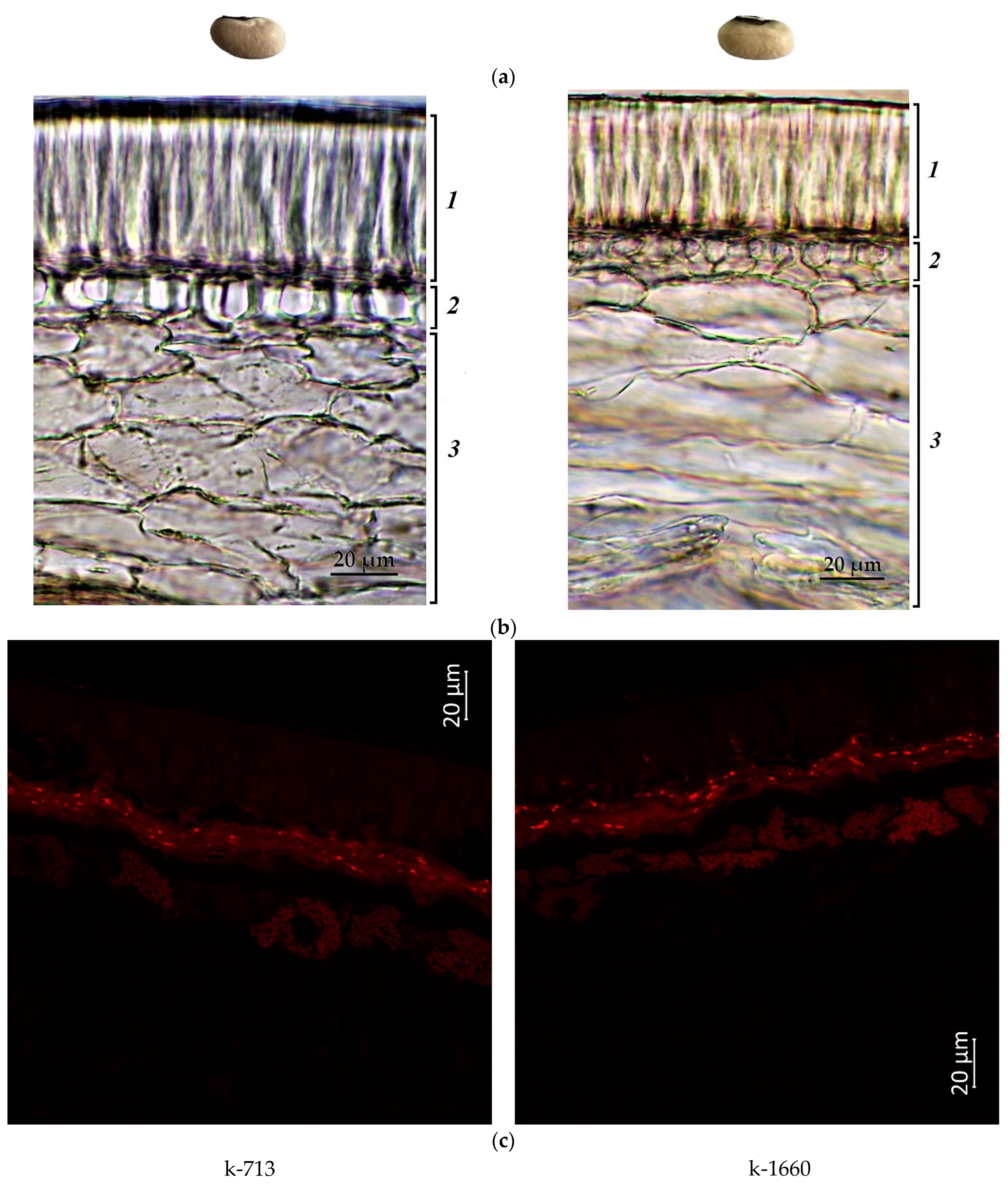
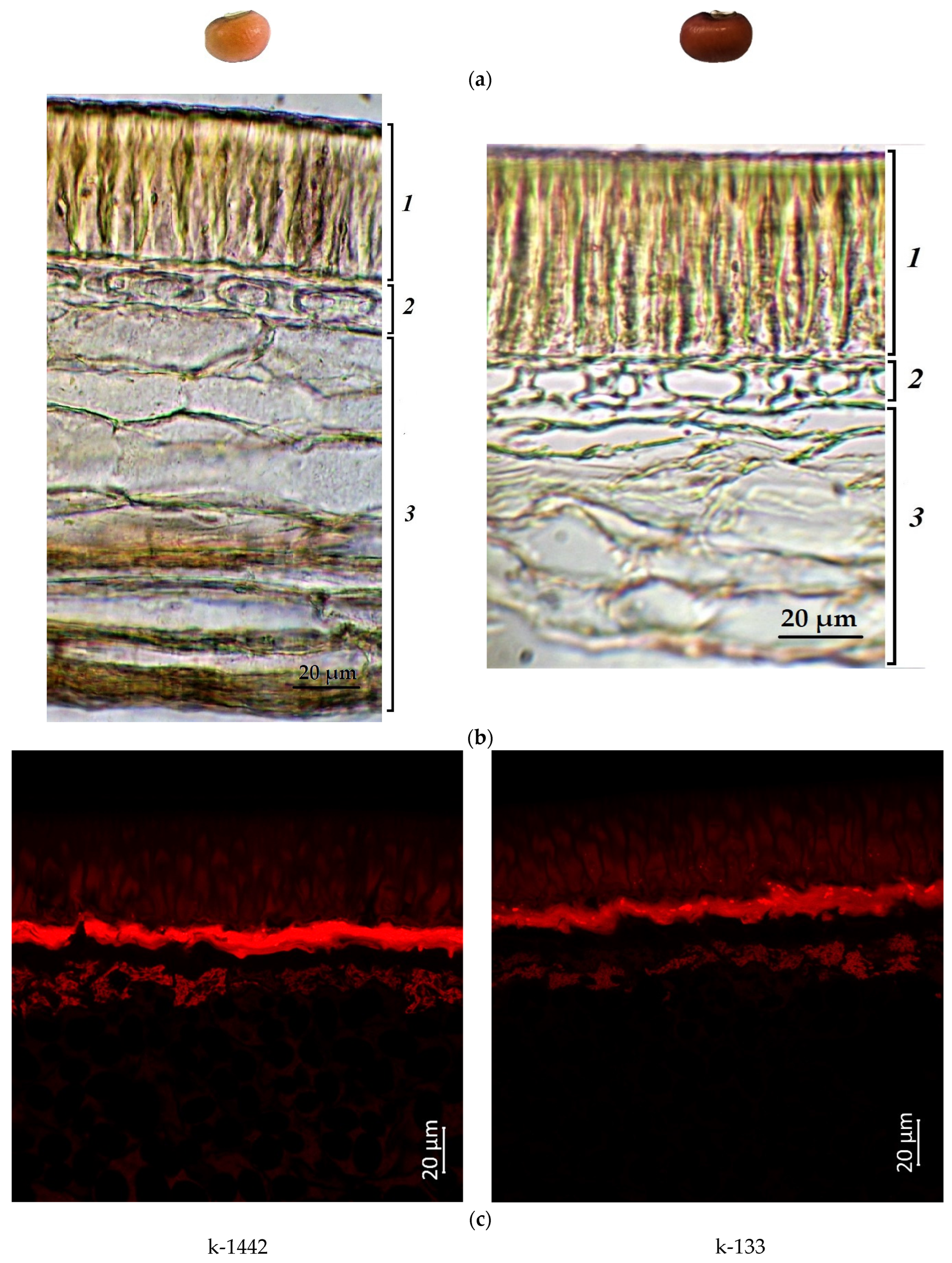
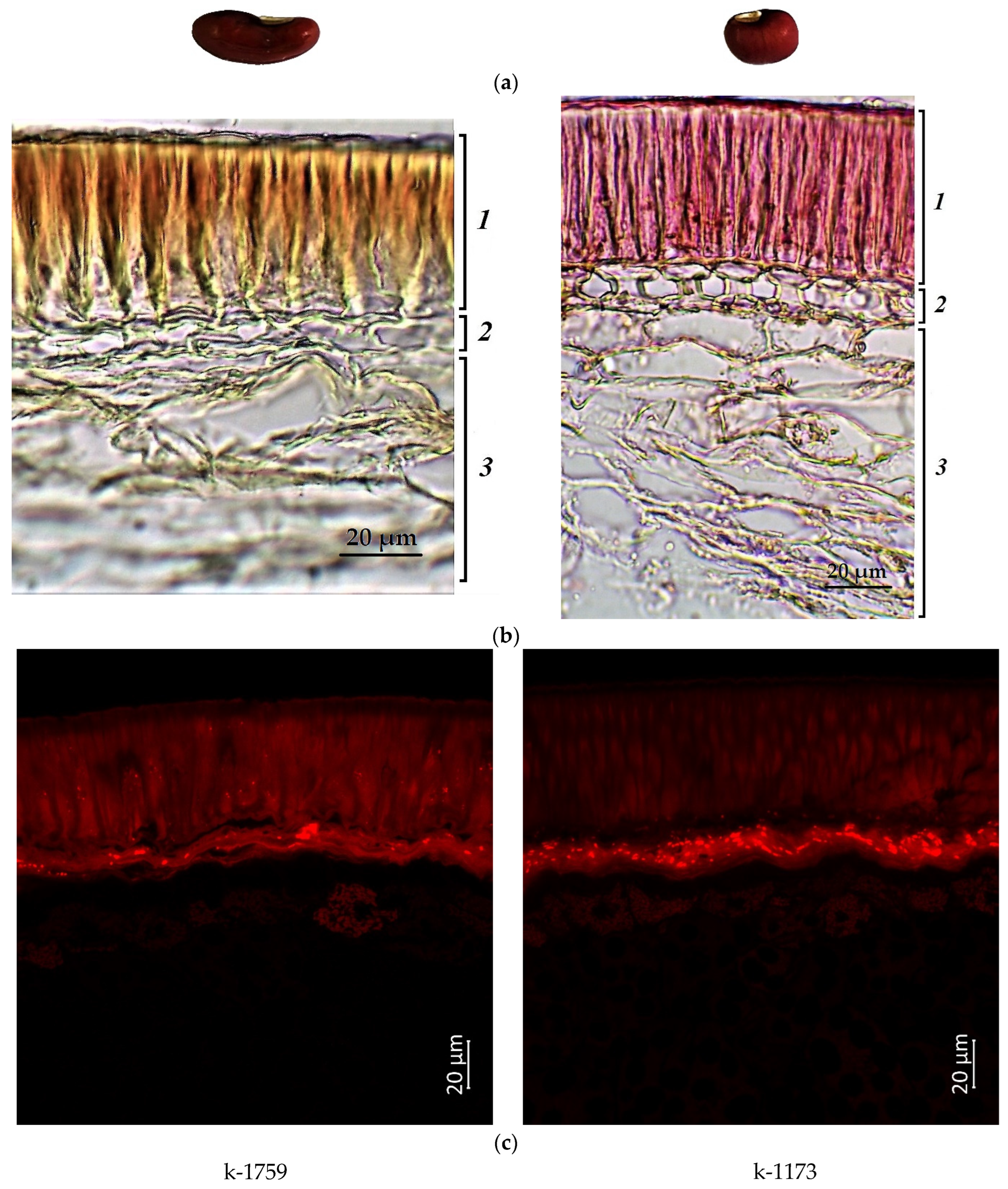
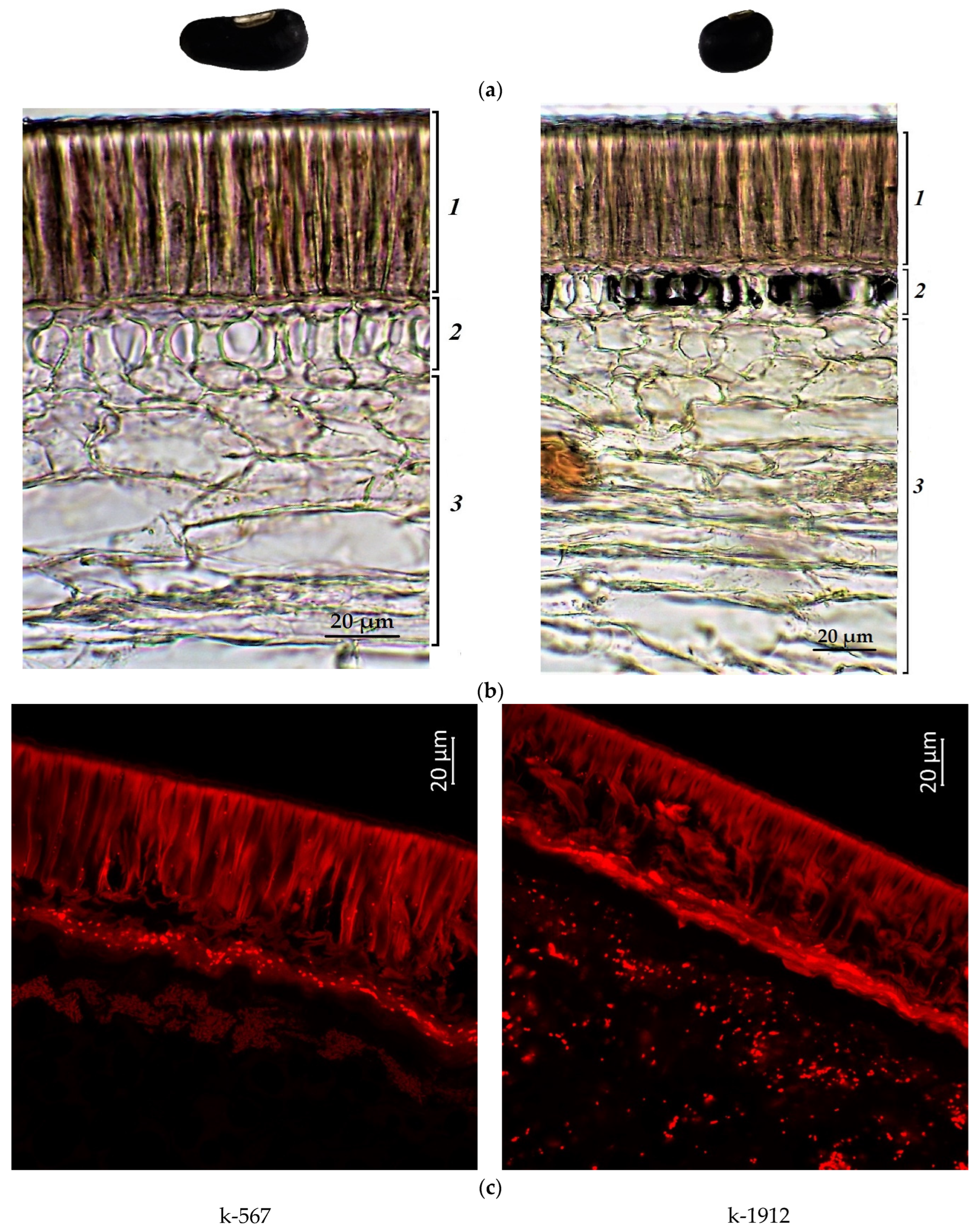
2.1. Confocal Laser Scanning Microscopy
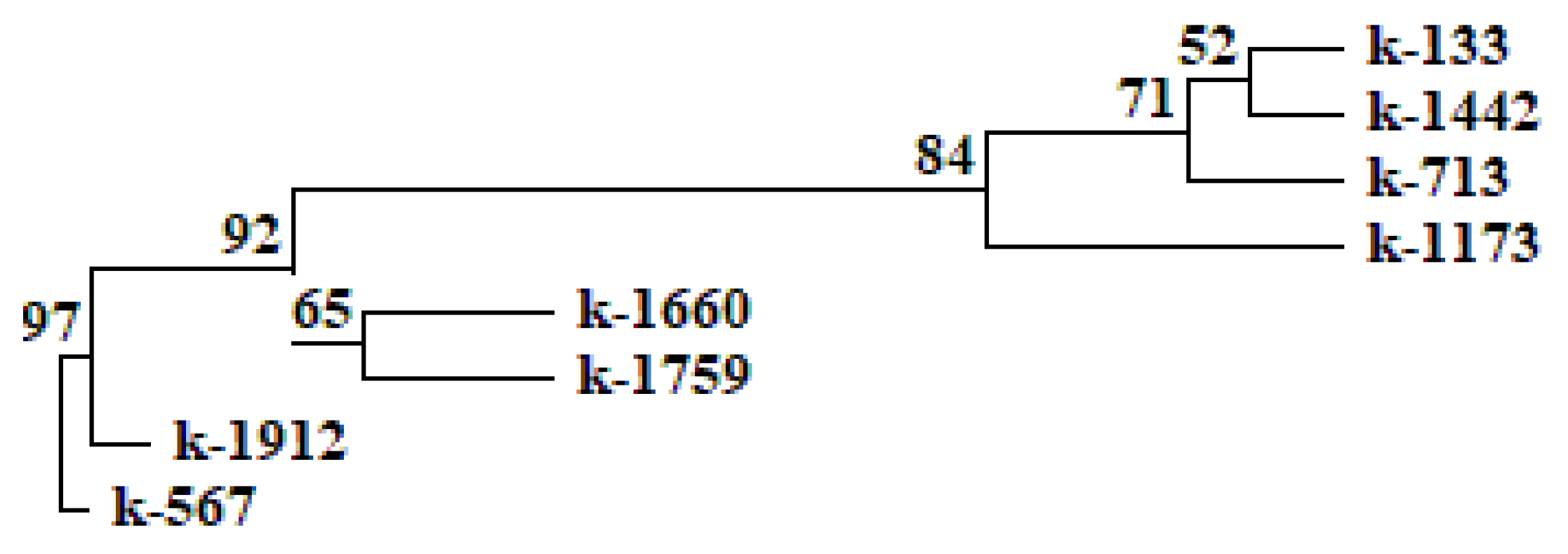
2.2. Anthocyanin Content in Cowpea Seeds
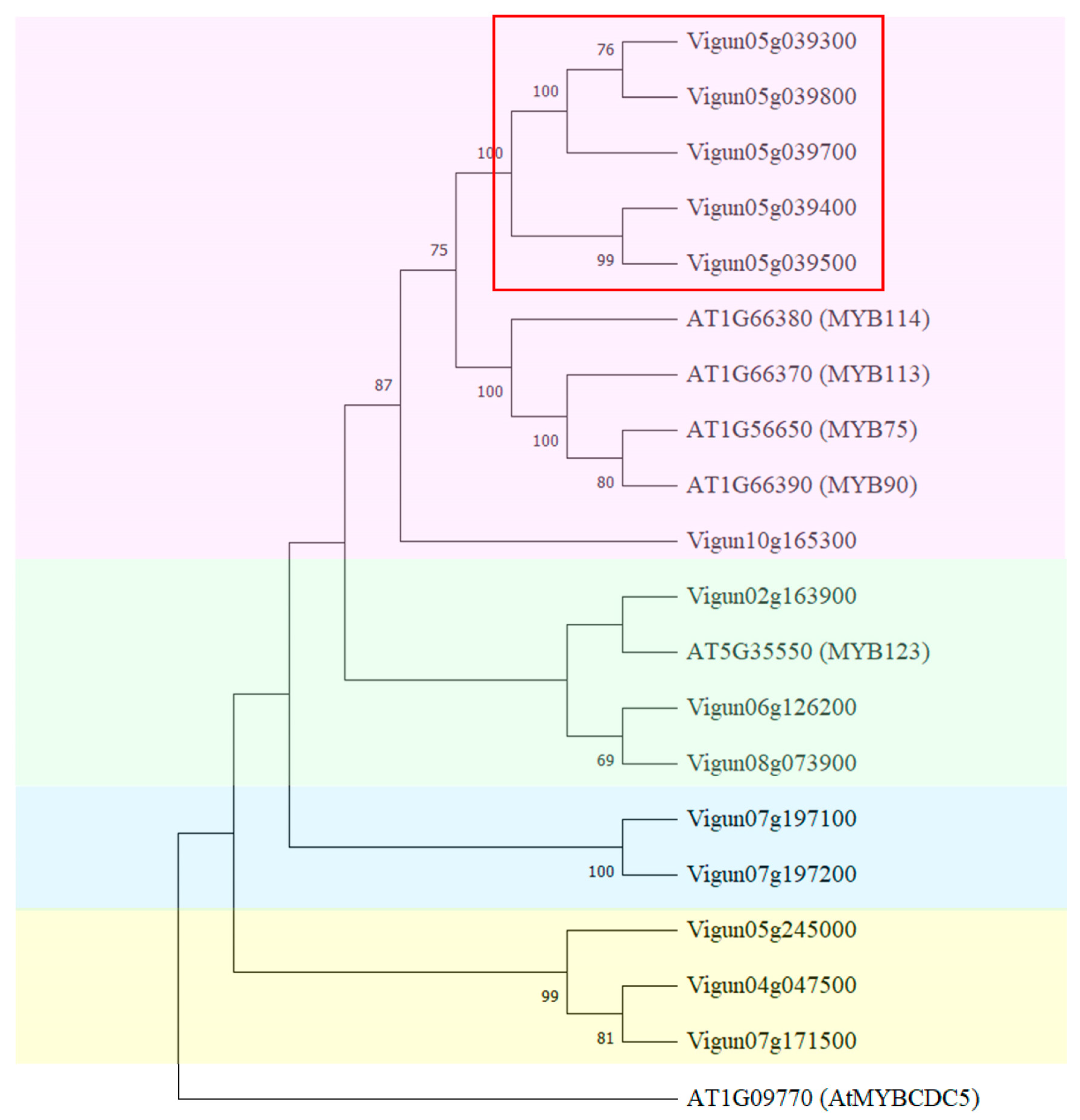
2.3. Identification and Phylogenetic Analysis of MYB-like Genes in Cowpea
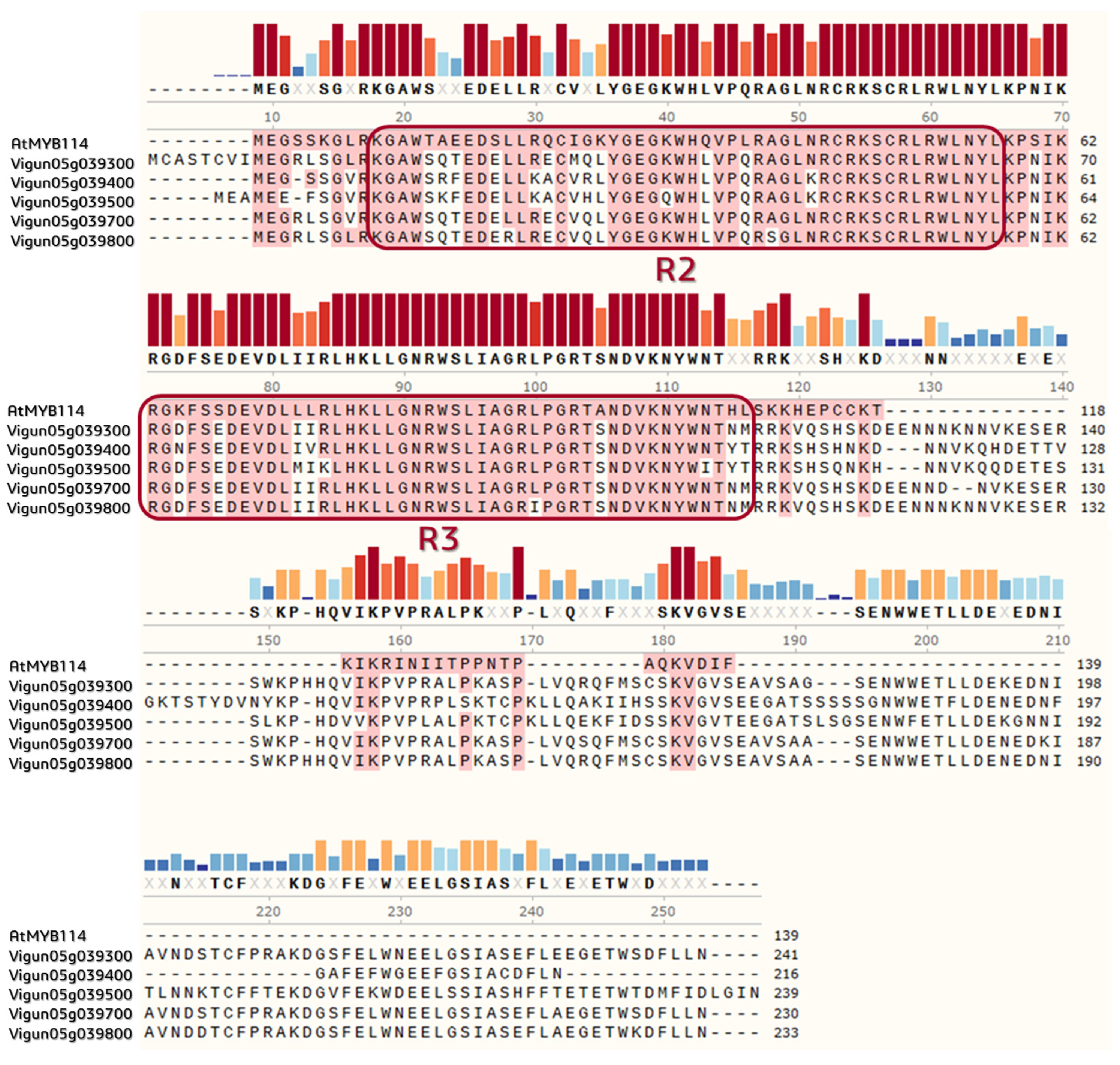
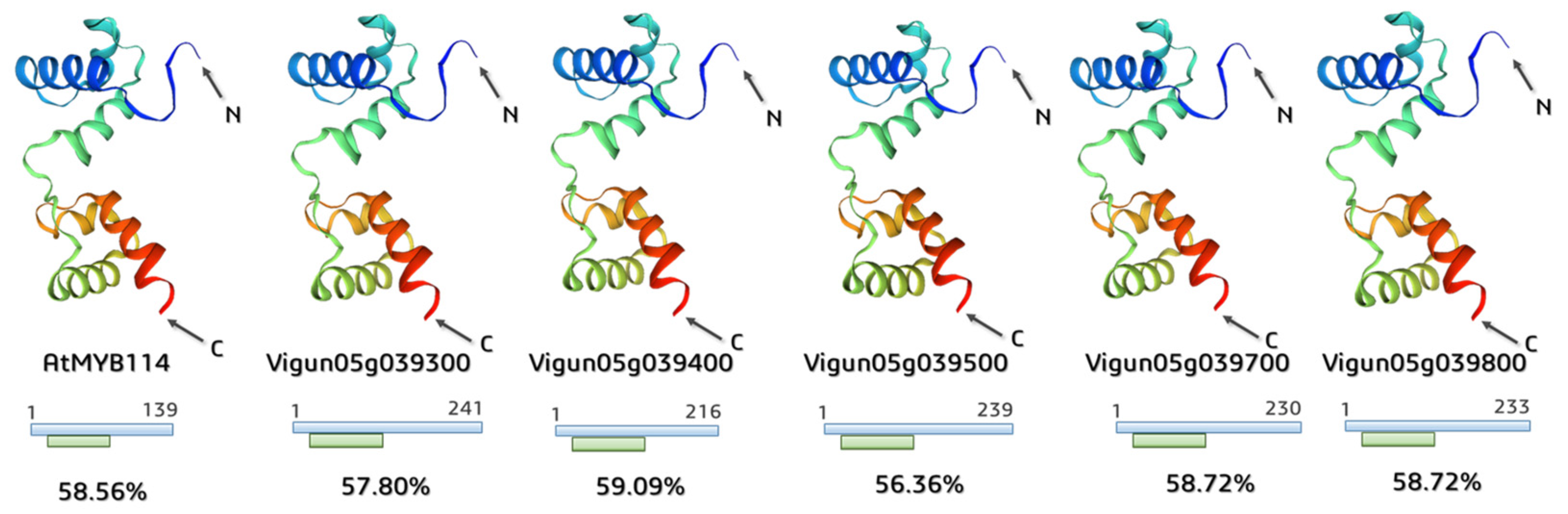
2.4. Annotation of the Functional Domains and Prediction of 3D MYB-Cluster Protein Structures
2.5. Amplification and Resequencing of the MYB-Cluster Genes on Chromosome 5
2.6. Statistical Analysis
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Confocal Laser Scanning Microscopy
4.3. Assessment of Anthocyanin Content in Seeds
4.4. Identification of Genes and Phylogenetic Analysis, Prediction of 3D Protein Structures
4.5. DNA Extraction
4.6. Primer Design and PCR Amplification, Sanger Sequencing
4.7. Statistical Data Processing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gerrano, A.S.; Jansen van Rensburg, W.S.; Venter, S.L.; Shargie, N.G.; Amelework, B.A.; Shimelis, H.A.; Labuschagne, M.T. Selection of cowpea genotypes based on grain mineral and total protein content. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2019, 69, 155–166. [Google Scholar] [CrossRef]
- Perchuk, I.; Shelenga, T.; Gurkina, M.; Miroshnichenko, E.; Burlyaeva, M. Composition of Primary and Secondary Metabolite Compounds in Seeds and Pods of Asparagus Bean (Vigna unguiculata (L.) Walp.) from China. Molecules 2020, 25, 3778. [Google Scholar] [CrossRef]
- Boukar, O.; Fatokun, C.A.; Roberts, P.A.; Abberton, M.; Huynh, B.L.; Close, T.J.; Kyei-Boahen, S.; Higgins, T.J.V.; Jeffrey, D.E. Cowpea. In Grain Legumes; De Ron, A.M., Ed.; Springer: New York, NY, USA, 2015; pp. 219–250. [Google Scholar] [CrossRef]
- Orita, A.; Musou-Yahada, A.; Shoji, T.; Oki, T.; Ohta, H. Comparison of Anthocyanins, Proanthocyanidin Oligomers and Antioxidant Capacity between Cowpea and Grain Legumes with Colored Seed Coat. Food Sci. Technol. Res. 2019, 25, 287–294. [Google Scholar] [CrossRef]
- Kan, L.; Nie, S.; Hu, J.; Liu, Z.; Xie, M. Antioxidant activities and anthocyanins composition of seed coats from twenty-six kidney bean cultivars. J. Funct. Foods 2016, 26, 622–631. [Google Scholar] [CrossRef]
- Awika, J.M.; Duodu, K.G. Bioactive polyphenols and peptides in cowpea (Vigna unguiculata) and their health promoting properties: A review. J. Funct. Foods 2017, 38, 686–697. [Google Scholar] [CrossRef]
- Gonçalves, A.; Goufo, P.; Barros, A.; Domínguez-Perles, R.; Trindade, H.; Rosa, E.A.S.; Ferreira, L.; Rodrigues, M. Cowpea (Vigna unguiculata L. Walp), a renewed multipurpose crop for a more sustainable agri-food system: Nutritional advantages and constraints. J. Sci. Food Agric. 2016, 96, 2941–2951. [Google Scholar] [CrossRef]
- Gonçalves, A.C.; Nunes, A.R.; Falcão, A.; Alves, G.; Silva, L.R. Dietary Effects of Anthocyanins in Human Health: A Comprehensive Review. Pharmaceuticals 2021, 14, 690. [Google Scholar] [CrossRef]
- Hershey, M.S.; Sotos-Prieto, M.; Ruiz-Canela, M.; Martinez-Gonzalez, M.A.; Cassidy, A.; Moffatt, S.; Kales, S.N. Anthocyanin Intake and Physical Activity: Associations with the Lipid Profile of a US Working Population. Molecules 2020, 25, 4398. [Google Scholar] [CrossRef]
- Lee, Y.M.; Yoon, Y.; Yoon, H.; Park, H.M.; Song, S.; Yeum, K.J. Dietary Anthocyanins against Obesity and Inflammation. Nutrients 2017, 9, 1089. [Google Scholar] [CrossRef]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Phenolic composition and antioxidant potential of grain legume seeds: A review. Food Res. Int. 2017, 101, 1–16. [Google Scholar] [CrossRef]
- Corso, M.; Perreau, F.; Mouille, G.; Lepiniec, L. Specialized phenolic compounds in seeds: Structures, functions, and regulations. Plant Sci. 2020, 296, 110471. [Google Scholar] [CrossRef]
- Lepiniec, L.; Debeaujon, I.; Routaboul, J.-M.; Baudry, A.; Pourcel, L.; Nesi, N.; Caboche, M. Genetics and biochemistry of seed flavonoids. Annu. Rev. Plant Biol. 2006, 57, 405–430. [Google Scholar] [CrossRef]
- Kumar, V.; Suman, U.; Rubal Yadav, S.K. Flavonoid Secondary Metabolite: Biosynthesis and Role in Growth and Development in Plants. In Recent Trends and Techniques in Plant Metabolic Engineering; Yadav, S.K., Kumar, V., Singh, S.P., Eds.; Springer: Singapore, 2018; pp. 19–45. [Google Scholar] [CrossRef]
- Havsteen, B.H. The biochemistry and medical significance of the flavonoids. Pharmacol. Ther. 2002, 96, 67–202. [Google Scholar] [CrossRef] [PubMed]
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, A.; Ohnishi, T. The Significance of the Study about the Biological Effects of Solar Ultraviolet Radiation Using the Exposed Facility on the International Space Station. Biol. Sci. Space 2004, 18, 255–260. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed]
- Makoi, J.H.J.R.; Belane, A.K.; Chimphango, S.B.M.; Dakora, F.D. Seed flavonoids and anthocyanins as markers of enhanced plant defence in nodulated cowpea (Vigna unguiculata L. Walp.). Field Crops Res. 2010, 118, 21–27. [Google Scholar] [CrossRef]
- Holton, T.A.; Cornish, E.C. Genetics and biochemistry of anthocyanin biosynthesis. Plant Cell 1995, 7, 1071–1083. [Google Scholar] [CrossRef] [PubMed]
- Winkel-Shirley, B. Flavonoid Biosynthesis. A Colorful Model for Genetics, Biochemistry, Cell Biology, and Biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Bogs, J.; Downey, M.O.; Harvey, J.S.; Ashton, A.R.; Tanner, G.J.; Robinson, S.P. Proanthocyanidin Synthesis and Expression of Genes Encoding Leucoanthocyanidin Reductase and Anthocyanidin Reductase in Developing Grape Berries and Grapevine Leaves. Plant Physiol. 2005, 139, 652–663. [Google Scholar] [CrossRef]
- Takahama, U.; Yamauchi, R.; Hirota, S. Isolation and characterization of a cyanidin-catechin pigment from adzuki bean (Vigna angularis). Food Chem. 2013, 141, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Wang, M.; Tu, L.; Nie, Y.; Lin, Y.; Zhang, X. The Flavonoid Pathway Regulates the Petal Colors of Cotton Flower. PLoS ONE 2013, 8, e72364. [Google Scholar] [CrossRef] [PubMed]
- Deroles, S. Anthocyanin Biosynthesis in Plant Cell Cultures: A Potential Source of Natural Colourants. In Anthocyanins; Springer: New York, NY, USA, 2008; pp. 108–167. [Google Scholar] [CrossRef]
- van der Kooi, C.J.; Ollerton, J. The origins of flowering plants and pollinators. Science 2020, 368, 1306–1308. [Google Scholar] [CrossRef] [PubMed]
- Narbona, E.; del Valle, J.C.; Arista, M.; Buide, M.L.; Ortiz, P.L. Major Flower Pigments Originate Different Colour Signals to Pollinators. Front. Ecol. Evol. 2021, 9, 743850. [Google Scholar] [CrossRef]
- Cardador-Martínez, A.; Loarca-Piña, G.; Oomah, B.D. Antioxidant Activity in Common Beans (Phaseolus vulgaris L.). J. Agric. Food Chem. 2002, 50, 6975–6980. [Google Scholar] [CrossRef]
- Beninger, C.W.; Hosfield, G.L. Antioxidant Activity of Extracts, Condensed Tannin Fractions, and Pure Flavonoids from Phaseolus vulgaris L. Seed Coat Color Genotypes. J. Agric. Food Chem. 2003, 51, 7879–7883. [Google Scholar] [CrossRef] [PubMed]
- Madhujith, T.; Naczk, M.; Shahidi, F. Antioxidant activity of common beans (Phaseolus vulgaris L.). J. Food Lipids 2004, 11, 220–233. [Google Scholar] [CrossRef]
- Espinosa-Alonso, L.G.; Lygin, A.; Widholm, J.M.; Valverde, M.E.; Paredes-Lopez, O. Polyphenols in Wild and Weedy Mexican Common Beans (Phaseolus vulgaris L.). J. Agric. Food Chem. 2006, 54, 4436–4444. [Google Scholar] [CrossRef]
- Chávez-Santoscoy, R.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Effect of Flavonoids and Saponins Extracted from Black Bean (Phaseolus vulgaris L.) Seed Coats as Cholesterol Micelle Disruptors. Plant Foods Hum. Nutr. 2013, 68, 416–423. [Google Scholar] [CrossRef]
- Aparicio-Fernandez, X.; Yousef, G.G.; Loarca-Pina, G.; de Mejia, E.; Lila, M.A. Characterization of Polyphenolics in the Seed Coat of Black Jamapa Bean (Phaseolus vulgaris L.). J. Agric. Food Chem. 2005, 53, 4615–4622. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhao, W.; Huang, Y.; Zhao, P.; Yang, K.; Wan, P.; Chu, L. Progress in Adzuki Bean Seed Coat Colour Studies. Plants 2023, 12, 3242. [Google Scholar] [CrossRef] [PubMed]
- Teixeira-Guedes, C.I.; Oppolzer, D.; Barros, A.I.; Pereira-Wilson, C. Phenolic rich extracts from cowpea sprouts decrease cell proliferation and enhance 5-fluorouracil effect in human colorectal cancer cell lines. J. Funct. Foods 2019, 60, 103452. [Google Scholar] [CrossRef]
- Harland, S.C. Inheritance of certain characters in the cowpea (Vigna sinensis). J. Genet. 1919, 8, 101–132. [Google Scholar] [CrossRef]
- Harland, S.C. Inheritance of certain characters in the cowpea (Vigna sinensis). II. J. Genet 1920, 10, 193–205. [Google Scholar] [CrossRef]
- Spillman, J. Inheritance of the “Eye” in Vigna. Am. Nat. 1911, XLV, 513–523. [Google Scholar] [CrossRef]
- Jin, H.; Cominelli, E.; Bailey, P.; Parr, A.; Mehrtens, F.; Jones, J.; Tonelli, C.; Weisshaar, B.; Martin, C. Transcriptional repression by AtMYB4 controls production of UV-protecting sunscreens in Arabidopsis. EMBO J. 2000, 19, 6150–6161. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.-F.; Fitzsimmons, K.; Khandelwal, A.; Kranz, R.G. CPC, a Single-Repeat R3 MYB, Is a Negative Regulator of Anthocyanin Biosynthesis in Arabidopsis. Mol. Plant 2009, 2, 790–802. [Google Scholar] [CrossRef]
- Herniter, I.A.; Muñoz-Amatriaín, M.; Lo, S.; Guom, Y.-N.; Close, T.J. Identification of Candidate Genes Controlling Black Seed Coat and Pod Tip Color in Cowpea (Vigna unguiculata [L.] Walp). G3 Genes|Genomes|Genet. 2018, 8, 3347–3355. [Google Scholar] [CrossRef]
- Herniter, I.A.; Lo, R.; Muñoz-Amatriaín, M.; Lo, S.; Guo, Y.-N.; Huynh, B.-L.; Lucas, M.; Jia, Z.; Roberts, P.A.; Lonardi, S.; et al. Seed Coat Pattern QTL and Development in Cowpea (Vigna unguiculata [L.] Walp.). Front. Plant Sci. 2019, 10, 1346. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Q.; Xie, X.; Cai, Y.; Li, J.; Feng, Y.; Zhang, Y. Integrated Metabolomics and Transcriptomics Analyses Reveal the Molecular Mechanisms Underlying the Accumulation of Anthocyanins and Other Flavonoids in Cowpea Pod (Vigna unguiculata L.). J. Agric. Food Chem. 2020, 68, 9260–9275. [Google Scholar] [CrossRef]
- Cheynier, V.; Comte, G.; Davies, K.M.; Lattanzio, V.; Martens, S. Plant phenolics: Recent advances on their biosynthesis, genetics, andecophysiology. Plant Physiol. Biochem. 2013, 72, 1–20. [Google Scholar] [CrossRef]
- Saito, K.; Yonekura-Sakakibara, K.; Nakabayashi, R.; Higashi, Y.; Yamazaki, M.; Tohge, T.; Fernie, A.R. The flavonoid biosynthetic pathway in Arabidopsis: Structural and genetic diversity. Plant Physiol. Biochem. 2013, 72, 21–34. [Google Scholar] [CrossRef]
- Weisshaar, B.; Jenkins, G.I. Phenylpropanoid biosynthesis and its regulation. Curr. Opin. Plant Biol. 1998, 1, 251–257. [Google Scholar] [CrossRef]
- Katiyar, A.; Smita, S.; Lenka, S.; Rajwanshi, R.; Chinnusamy, V.; Bansal, K. Genome-wide classification and expression analysis of MYB transcription factor families in rice and Arabidopsis. BMC Genom. 2012, 13, 544. [Google Scholar] [CrossRef] [PubMed]
- Prouse, M.B.; Campbell, M.M. The interaction between MYB proteins and their target DNA binding sites. Biochim. Biophys. Acta-Gene Regul. Mech. 2012, 1819, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Koes, R.; Verweij, W.; Quattrocchio, F. Flavonoids: A colorful model for the regulation and evolution of biochemical pathways. Trends Plant Sci. 2005, 10, 236–242. [Google Scholar] [CrossRef]
- Li, S. Transcriptional control of flavonoid biosynthesis. Plant Signal. Behav. 2014, 9, e27522. [Google Scholar] [CrossRef]
- Adzhieva, V.F.; Babak, O.G.; Shoeva, O.Y.; Kilchevsky, A.V.; Khlestkina, E.K. Molecular-genetic mechanisms underlying fruit and seed coloration in plants. Vavilov J. Genet. Breed. 2015, 19, 561–573. [Google Scholar] [CrossRef]
- Lipsick, J. One billion years of Myb. Oncogene 1996, 13, 223–235. [Google Scholar]
- Oda, M.; Furukawa, K.; Ogata, K.; Sarai, A.; Ishii, S.; Nishimura, Y.; Nakamura, H. Identification of indispensable residues for specific DNA-binding in the imperfect tandem repeats of c-Myb R2R3. Protein Eng. Des. Sel. 1997, 10, 1407–1414. [Google Scholar] [CrossRef]
- Ogata, K.; Hojo, H.; Aimoto, S.; Nakai, T.; Nakamura, H.; Sarai, A.; Ishii, S.; Nishimura, Y. Solution structure of a DNA-binding unit of Myb: A helix-turn-helix-related motif with conserved tryptophans forming a hydrophobic core. Proc. Natl. Acad. Sci. USA 1992, 89, 6428–6432. [Google Scholar] [CrossRef]
- Ogata, K.; Kanei-Ishii, C.; Sasaki, M.; Hatanaka, H.; Nagadoi, A.; Enari, M.; Nakamura, H.; Nishimura, Y.; Ishii, S.; Sarai, A. The cavity in the hydrophobic core of Myb DNA-binding domain is reserved for DNA recognition and trans-activation. Nat. Struct. Biol. 1996, 3, 178–187. [Google Scholar] [CrossRef]
- Wang, B.; Luo, Q.; Li, Y.; Yin, L.; Zhou, N.; Li, X.; Gan, J.; Dong, A. Structural insights into target DNA recognition by R2R3-MYB transcription factors. Nucleic Acids Res. 2019, 48, 460–471. [Google Scholar] [CrossRef]
- Stracke, R.; Werber, M.; Weisshaar, B. The R2R3-MYB gene family in Arabidopsis thaliana. Curr. Opin. Plant Biol. 2001, 4, 447–456. [Google Scholar] [CrossRef]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef]
- Stracke, R.; Jahns, O.; Keck, M.; Tohge, T.; Niehaus, K.; Fernie, A.R.; Weisshaar, B. Analysis of production of Flavonol glycosides-dependent flavonol glycoside accumulation in Arabidopsis thaliana plants reveals MYB11-, MYB12- and MYB111-independent flavonol glycoside accumulation. New Phytol. 2010, 188, 985–1000. [Google Scholar] [CrossRef]
- Gonzalez, A.; Zhao, M.; Leavitt, J.M.; Lloyd, A.M. Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex in Arabidopsis seedlings. Plant J. 2008, 53, 814–827. [Google Scholar] [CrossRef] [PubMed]
- Debeaujon, I.; Nesi, N.; Perez, P.; Devic, M.; Grandjean, O.; Caboche, M.; Lepiniec, L. Proanthocyanidin-Accumulating Cells in Arabidopsis Testa: Regulation of Differentiation and Role in Seed Development. Plant Cell 2003, 15, 2514–2531. [Google Scholar] [CrossRef] [PubMed]
- Baudry, A.; Heim, M.A.; Dubreucq, B.; Caboche, M.; Weisshaar, B.; Lepiniec, L. TT2, TT8, and TTG1 synergistically specify the expression of BANYULS and proanthocyanidin biosynthesis in Arabidopsis thaliana. Plant J. 2004, 39, 366–380. [Google Scholar] [CrossRef] [PubMed]
- LaFountain, A.M.; Yuan, Y.W. Repressors of anthocyanin biosynthesis. New Phytol. 2021, 231, 933–949. [Google Scholar] [CrossRef]
- Chen, L.H.; Hu, B.; Qin, Y.H.; Hu, G.B.; Zhao, J.T. Advance of the negative regulation of anthocyanin biosynthesis by MYB transcription factors. Plant Physiol. Biochem. 2019, 136, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Hiratsu, K.; Matsui, K.; Koyama, T.; Ohme-Takagi, M. Dominant repression of target genes by chimeric repressors that include the EAR motif, a repression domain, in Arabidopsis. Plant J. 2003, 34, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K.; Umemura, Y.; Ohme-Takagi, M. AtMYBL2, a protein with a single MYB domain, acts as a negative regulator of anthocyanin biosynthesis in Arabidopsis. Plant J. 2008, 55, 954–967. [Google Scholar] [CrossRef]
- Harris, N.; Boulter, D. Protein Body Formation in Cotyledons of Developing Cowpea (Vigna unguiculata) Seeds. Ann. Bot. 1976, 40, 739–744. [Google Scholar] [CrossRef]
- Sefa-Dedeh, S.; Stanley, D.W. Microstructure of cowpea variety Adua Ayera [grown in Ghana]. Cereal Chem. 1979, 56, 367–371. [Google Scholar]
- Talamond, P.; Verdeil, J.-L.; Conéjéro, G. Secondary Metabolite Localization by Autofluorescence in Living Plant Cells. Molecules 2015, 20, 5024–5037. [Google Scholar] [CrossRef]
- Collings, D.A. Anthocyanin in the Vacuole of Red Onion Epidermal Cells Quenches Other Fluorescent Molecules. Plants 2019, 8, 596. [Google Scholar] [CrossRef]
- Mackon, E.; Ma, Y.; Jeazet Dongho Epse Mackon, G.C.; Li, Q.; Zhou, Q.; Liu, P. Subcellular Localization and Vesicular Structures of Anthocyanin Pigmentation by Fluorescence Imaging of Black Rice (Oryza sativa L.) Stigma Protoplast. Plants 2021, 10, 685. [Google Scholar] [CrossRef]
- Jo, H.; Lee, J.Y.; Cho, H.; Choi, H.J.; Son, C.K.; Bae, J.S.; Bilyeu, K.; Song, J.T.; Lee, J.-D. Genetic Diversity of Soybeans (Glycine max (L.) Merr.) with Black Seed Coats and Green Cotyledons in Korean Germplasm. Agronomy 2021, 11, 581. [Google Scholar] [CrossRef]
- Hirayama, T.; Shinozaki, K. A cdc5+ homolog of a higher plant, Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1996, 93, 13371–13376. [Google Scholar] [CrossRef]
- Yao, S.; Jiang, C.; Huang, Z.; Torres-Jerez, I.; Chang, J.; Zhang, H.; Udvardi, M.; Liu, R.; Verdier, J. The Vigna unguiculata Gene Expression Atlas (Vu GEA) from de novo assembly and quantification of RNA-seq data provides insights into seed maturation mechanisms. Plant J. 2016, 88, 318–327. [Google Scholar] [CrossRef]
- Razgonova, M.P.; Burlyaeva, M.O.; Zinchenko, Y.N.; Krylova, E.A.; Chunikhina, O.A.; Ivanova, N.M.; Zakharenko, A.M.; Golokhvast, K.S. Identification and Spatial Distribution of Bioactive Compounds in Seeds Vigna unguiculata (L.) Walp. by Laser Microscopy and Tandem Mass Spectrometry. Plants 2022, 11, 2147. [Google Scholar] [CrossRef]
- Affrifah, N.S.; Phillips, R.D.; Saalia, F.K. Cowpeas: Nutritional profile, processing methods and products—A review. Legum Sci. 2022, 4, e131. [Google Scholar] [CrossRef]
- Ogawa, Y.; Miyashita, K.; Shimizu, H.; Sugiyama, J. Three-dimensional Internal Structure of a Soybean Seed by Observation of Autofluorescence of Sequential Sections. J. Jpn. Soc. Food Sci. Technol. 2003, 50, 213–217. [Google Scholar] [CrossRef]
- Gutiérrez-Uribe, J.A.; Romo-Lopez, I.; Serna-Saldívar, S.O. Phenolic composition and mammary cancer cell inhibition of extracts of whole cowpeas (Vigna unguiculata) and its anatomical parts. J Funct. Foods 2011, 3, 290–297. [Google Scholar] [CrossRef]
- Singh, B.; Singh, N.; Thakur, S.; Kaur, A. Ultrasound assisted extraction of polyphenols and their distribution in whole mung bean, hull and cotyledon. J. Food Sci. Technol. 2017, 54, 921–932. [Google Scholar] [CrossRef] [PubMed]
- Okafor, J.N.C.; Rautenbauch, F.; Meyer, M.; Le Roes-Hill, M.; Harris, T.; Jideani, V.A. Phenolic content, antioxidant, cytotoxic and antiproliferative effects of fractions of Vigna subterraenea (L.) verdc from Mpumalanga, South Africa. Heliyon 2021, 7, e08397. [Google Scholar] [CrossRef] [PubMed]
- Jeng, T.L.; Shih, Y.J.; Wu, M.T.; Sung, J.M. Comparisons of flavonoids and anti-oxidative activities in seed coat, embryonic axis and cotyledon of black soybeans. Food Chem. 2010, 123, 1112–1116. [Google Scholar] [CrossRef]
- Moïse, J.A.; Han, S.; Gudynaitę-Savitch, L.; Johnson, D.A.; Miki, B.L.A. Seed coats: Structure, development, composition, and biotechnology. In Vitro Cell. Dev. Biol.-Plant 2005, 41, 620–644. [Google Scholar] [CrossRef]
- Wood, J.A.; Knights, E.J.; Choct, M. Morphology of Chickpea Seeds (Cicer arietinum L.): Comparison of desi and kabuli Types. Int. J. Plant Sci. 2011, 172, 632–643. [Google Scholar] [CrossRef]
- Tsamo, A.T.; Mohammed, H.; Mohammed, M.; Papoh Ndibewu, P.; Dapare Dakora, F. Seed coat metabolite profiling of cowpea (Vigna unguiculata L. Walp.) accessions from Ghana using UPLC-PDA-QTOF-MS and chemometrics. Nat. Prod. Res. 2020, 34, 1158–1162. [Google Scholar] [CrossRef] [PubMed]
- Saikumar, P.; Murali, R.; Reddy, E.P. Role of tryptophan repeats and flanking amino acids in Myb-DNA interactions. Proc. Natl. Acad. Sci. USA 1990, 87, 8452–8456. [Google Scholar] [CrossRef]
- Kanei-Ishii, C.; Sarai, A.; Sawazaki, T.; Nakagoshi, H.; He, D.N.; Ogata, K.; Nishimura, Y.; Ishii, S. The tryptophan cluster: A hypothetical structure of the DNA-binding domain of the myb protooncogene product. J. Biol. Chem. 1990, 265, 19990–19995. [Google Scholar] [CrossRef]
- Muravieva, D.A.; Bubenchikova, V.N.; Belikov, V.V. Spectrophotometric determination of the amount of anthocyanins in flowers of blue cornflower. Pharmacy 1984, 36, 28–29. [Google Scholar]
- Dash, S.; Campbell, J.D.; Cannon, E.K.S.; Cleary, A.M.; Huang, W.; Kalberer, S.R.; Karingula, V.; Rice, A.G.; Singh, J.; Umale, P.E.; et al. Legume information system (LegumeInfo.org): A key component of a set of federated data resources for the legume family. Nucleic Acids Res. 2016, 44, D1181–D1188. [Google Scholar] [CrossRef]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Corpet, F. Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res. 1988, 16, 10881–10890. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed]
- Amorim, L.L.B.; Ferreira-Neto, J.R.C.; Bezerra-Neto, J.P.; Pandolfi, V.; de Araújo, F.T.; da Silva Matos, M.K.; Santos, M.G.; Kido, E.A.; Benko-Iseppon, A.M. Cowpea and abiotic stresses: Identification of reference genes for transcriptional profiling by qPCR. Plant Methods 2018, 14, 88. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
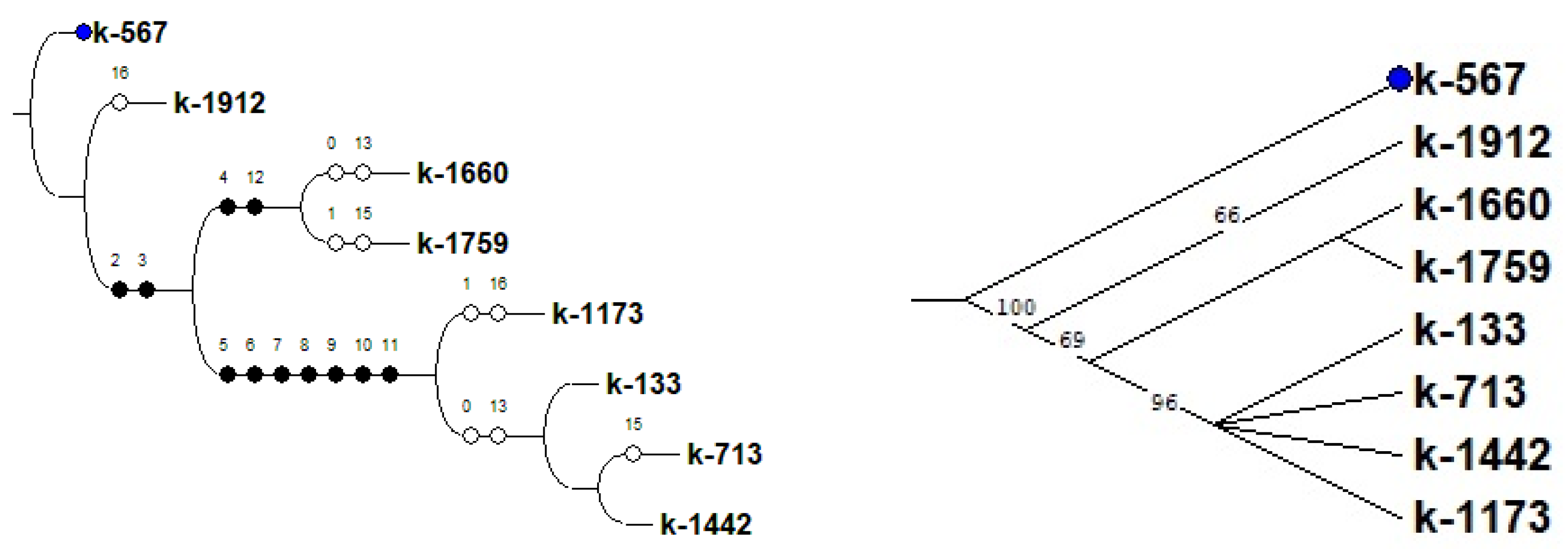{kind=link}
{kind=link}
| Accession | Seed Color | OD510–OD657 * | Criterion, p ** | Anthocyanins (mg/100 g DW) | |
|---|---|---|---|---|---|
| Student’s t-Test | Mann–Whitney U Test | ||||
| k-713 | white | 0.032 ± 0.003 | – | – | – |
| k-1660 | white | 0.032 ± 0.001 | – | – | – |
| k-1442 | light brown | 0.032 ± 0.003 | 0.26 | 0.23 | – |
| k-133 | brown | 0.039 ± 0.007 | 0.37 | 0.52 | – |
| k-1759 | reddish brown | 0.065 ± 0.006 | 0.07 × 10−4 | 0.01 | 2.56 ± 0.05 |
| k-1173 | reddish brown | 0.076 ± 0.005 | 0.01 × 10−5 | 0.05 × 10−1 | 2.94 ± 0.12 |
| k-567 | black | 2.295 ± 0.201 | 0.01 × 10−10 | 0.05 × 10−1 | 90.88 ± 7.87 |
| k-1912 | black | 4.788 ± 0.063 | 0.02 × 10−14 | 0.03 | 175.16 ± 4.61 |
| AtMYB113 | AtMYB114 | AtMYB90 | Vigun05g039300 | Vigun05g039400 | Vigun05g039500 | Vigun05g039700 | Vigun05g039800 | |
|---|---|---|---|---|---|---|---|---|
| AtMYB113 | - | 80.7% | 70.8% | 50.5% | 57.6% | 59.9% | 62.7% | 61.5% |
| AtMYB114 | 80.7% | - | 87.9% | 79.3% | 78.4% | 74.5% | 71.2% | 76.6% |
| AtMYB90 | 70.8% | 87.9% | - | 64.2% | 62.1% | 61.2% | 64.4% | 62.8% |
| Vigun05g039300 | 50.5% | 79.3% | 64.2% | - | 57.6% | 59.9% | 62.7% | 61.5% |
| Vigun05g039400 | 57.6% | 78.4% | 62.1% | 65.2% | - | 71.9% | 66.8% | 64.8% |
| Vigun05g039500 | 59.9% | 74.5% | 61.2% | 69.2% | 71.9% | - | 69.0% | 67.5% |
| Vigun05g039700 | 62.7% | 71.2% | 64.4% | 95.3% | 66.8% | 69.0% | - | 94.8% |
| Vigun05g039800 | 61.5% | 76.6% | 62.8% | 96.1% | 64.8% | 67.5% | 94.8% | - |
| VIR Catalogue Number | Color of Seed Coat | Presence or Absence of PCR Product with Primer Pairs | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 393-300-1 | 393-300-2 | Del-st-F/300-1-R | 394-400-1 | 395-500-1 | 395-500-2 | 395-500-3 | 395-500-4 | 397-700-1 | Del-end-F/ Del-st-R | ||
| К-713 | white | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| К-1660 | white | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 |
| К-1442 | light brown | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| К-133 | brown | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| К-1173 | reddish-brown | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| К-1759 | reddish-brown | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 |
| К-567 | black | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| К-1912 | black | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| VIR Catalogue Number | Seed Coat Color | Gene | |||
|---|---|---|---|---|---|
| Vigun05g039300 | Vigun05g039400 | Vigun05g039500 | Vigun05g039700 | ||
| k-713 | White | ||||
| k-1660 | White | ||||
| k-1442 | Light brown | ||||
| k-133 | Brown | ||||
| k-1173 | Reddish brown | ||||
| k-1759 | Reddish brown | ||||
| k-567 | Black | ||||
| k-1912 | Black | ||||
| No | VIR Catalogue Number | Type of Accessions | Country of Origin | Color of Seed Coat |
|---|---|---|---|---|
| 1 | k-133 | Landrace | Russia | brown |
| 2 | k-567 | Landrace | India | black |
| 3 | k-713 | Landrace | Trinidad and Tobago | white |
| 4 | k-1173 | Landrace | Somalia | reddish brown |
| 5 | k-1442 | Landrace | Nigeria | light brown |
| 6 | k-1660 | Landrace | France | white |
| 7 | k-1759 | Landrace | Kyrgyzstan | reddish brown |
| 8 | k-1912 | Landrace | Bolivia | black |
| Primer | Gene | Sequence (5′-3′) | Reference |
|---|---|---|---|
| 300-1-F | Vigun05g039300 | GGTGGTTTTGACGGTAAGCA | [41] |
| 300-1-R | TGTCCAACTCCACACCTTGA | ||
| 300-2-F * | ATGTGTGCAAGCACTTGTGTG | ||
| 300-2-R * | TAGAGACACCAACTGCGAATAC | ||
| 400-1-F | Vigun05g039400 | AGCATGATGAGACGACCGTA | [41] |
| 400-1-R | CCAAACTCTTCGCCCCAAAA | ||
| 500-1-F | Vigun05g039500 | TCCCTGATTGCAGGAAGACT | [41] |
| 500-1-R | CCAATTCTCCGACCCTGATA | ||
| 500-2-F * | GGGTTGAAGAGATGCAGGAA | ||
| 500-2-R * | ACCTGCATATTCATTCACAAACA | ||
| 500-3-F * | ATCTCATCTAGCGGTGCTTATAC | ||
| 500-3-R * | ATCTACAACTCTTCCTGCATCTC | ||
| 500-4-F * | TTATCAGGGTCGGAGAATTGG | ||
| 500-4-R * | GTAATCTGGAGGAGGCAGAAT | ||
| 700-1-F | Vigun05g039700 | ACAGGGTTATGCATTCTGAGC | [41] |
| 700-1-R | GGGATGAGAGAGATGGCGAA | ||
| Del-st-F | AGGGAAAGATGAGTGCAGGC | ||
| Del-st-R | AGGTCGCCCAACTTCACATT | ||
| Del-end-F | AGCCTCTGAGAATTGGTGGG | ||
| Del-end-R | Vigun05g039700 | GGGATGAGAGAGATGGCGAA | |
| Act-F | Vigun04g203000 | TCAGGTGTCCAGAGGTGTTGTA | [97] |
| Act-R | ATGGTTGTGCCTCCTGAAAGTA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krylova, E.А.; Mikhailova, A.S.; Zinchenko, Y.N.; Perchuk, I.N.; Razgonova, M.P.; Khlestkina, E.K.; Burlyaeva, M.O. The Content of Anthocyanins in Cowpea (Vigna unguiculata (L.) Walp.) Seeds and Contribution of the MYB Gene Cluster to Their Coloration Pattern. Plants 2023, 12, 3624. https://doi.org/10.3390/plants12203624
Krylova EА, Mikhailova AS, Zinchenko YN, Perchuk IN, Razgonova MP, Khlestkina EK, Burlyaeva MO. The Content of Anthocyanins in Cowpea (Vigna unguiculata (L.) Walp.) Seeds and Contribution of the MYB Gene Cluster to Their Coloration Pattern. Plants. 2023; 12(20):3624. https://doi.org/10.3390/plants12203624
Chicago/Turabian StyleKrylova, Ekaterina А., Aleksandra S. Mikhailova, Yulia N. Zinchenko, Irina N. Perchuk, Mayya P. Razgonova, Elena K. Khlestkina, and Marina O. Burlyaeva. 2023. "The Content of Anthocyanins in Cowpea (Vigna unguiculata (L.) Walp.) Seeds and Contribution of the MYB Gene Cluster to Their Coloration Pattern" Plants 12, no. 20: 3624. https://doi.org/10.3390/plants12203624
APA StyleKrylova, E. А., Mikhailova, A. S., Zinchenko, Y. N., Perchuk, I. N., Razgonova, M. P., Khlestkina, E. K., & Burlyaeva, M. O. (2023). The Content of Anthocyanins in Cowpea (Vigna unguiculata (L.) Walp.) Seeds and Contribution of the MYB Gene Cluster to Their Coloration Pattern. Plants, 12(20), 3624. https://doi.org/10.3390/plants12203624