


Isolation and Characterization of Erianthus arundinaceus Phosphate Transporter 1 (PHT1) Gene Promoter and 5′ Deletion Analysis of Transcriptional Regulation Regions under Phosphate Stress in Transgenic Tobacco

 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
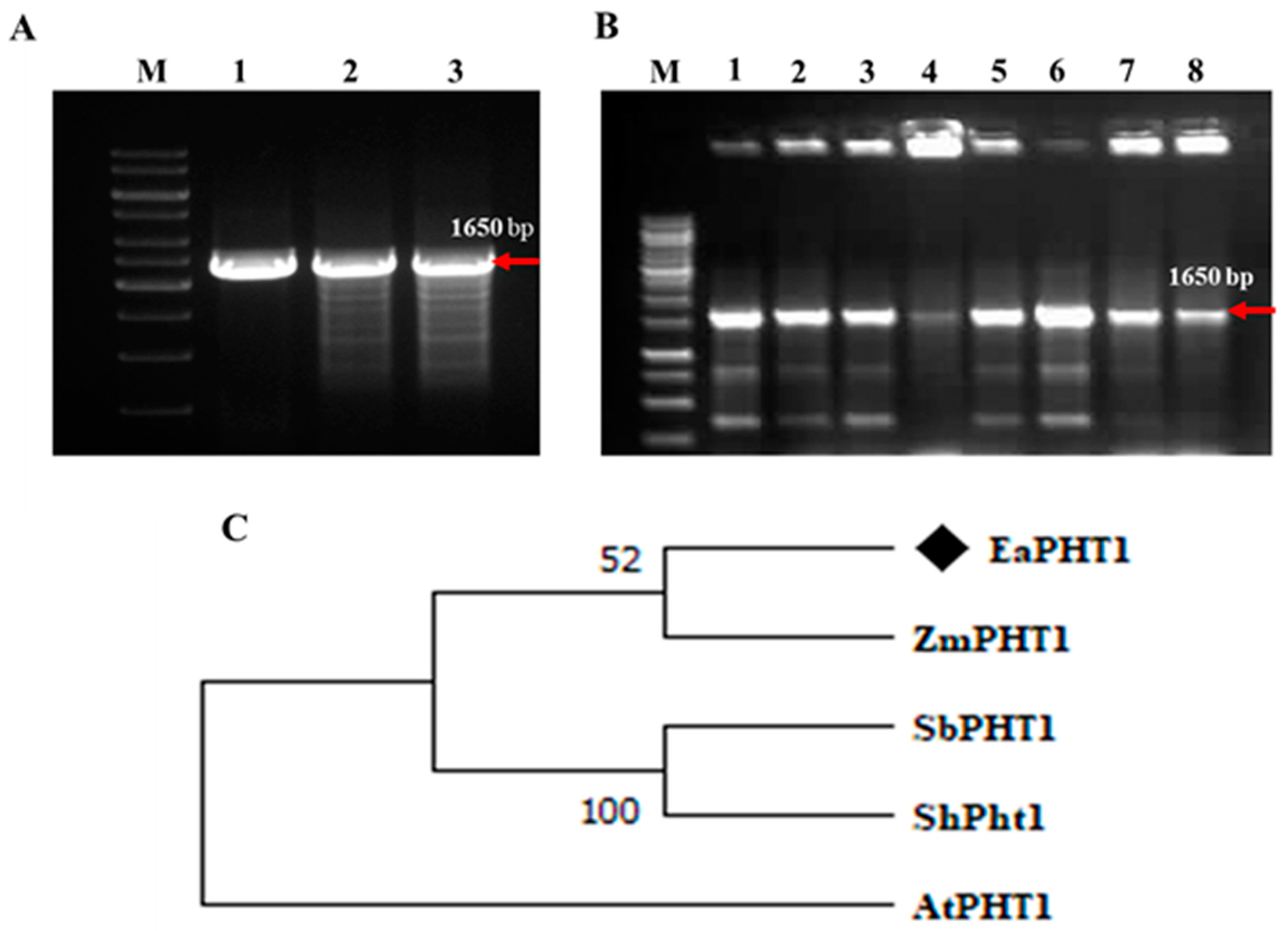
2.1. Isolation and Sequence Analysis of EaPHT1;2 Gene from E. arundinaceus
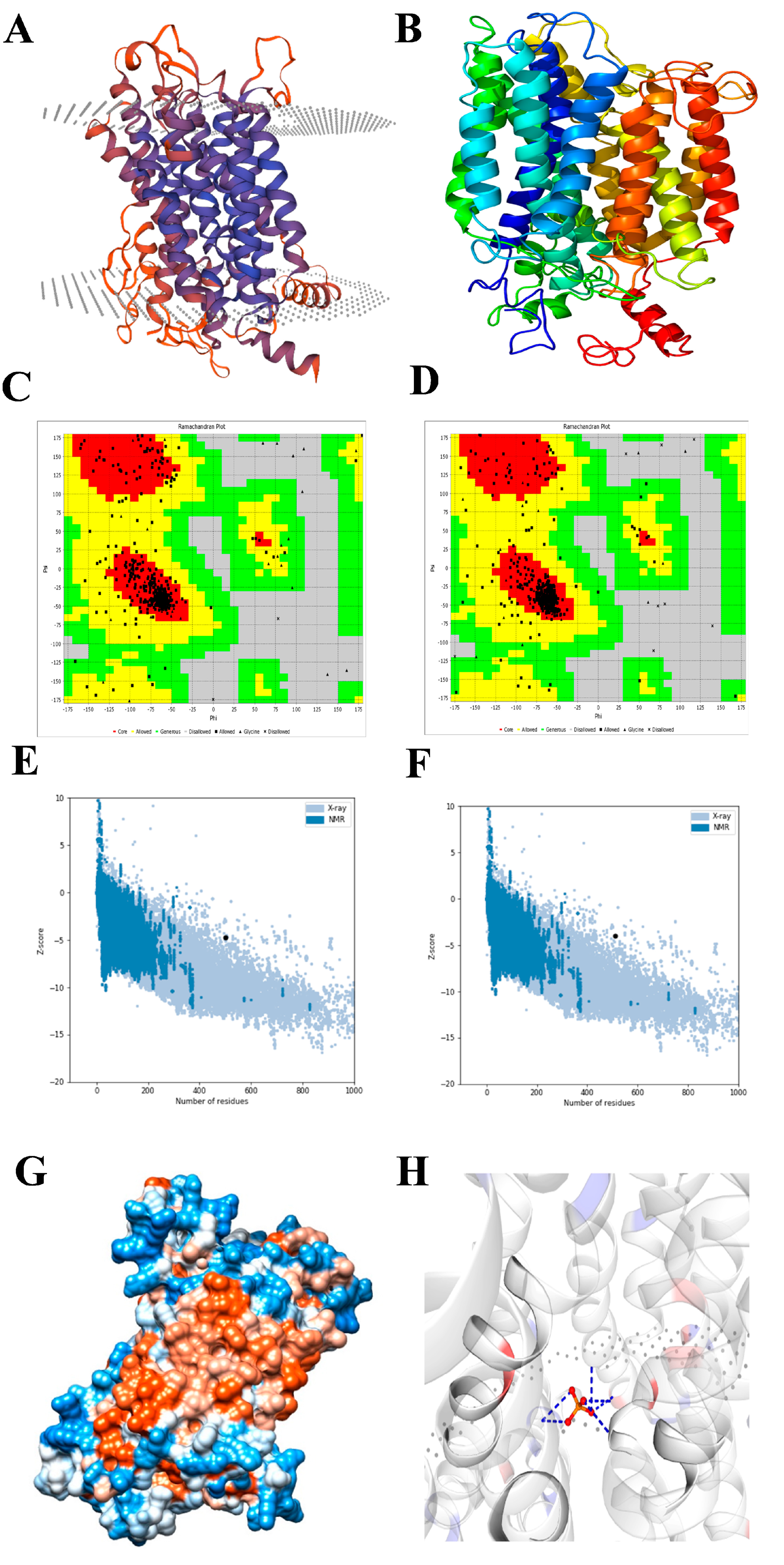
2.2. Structural Prediction of the EaPHT1;2 Protein
2.3. Protein Association Network and Subcellular Localization of Analysis
2.4. Isolation of PHT1 Promoter Region Using a Genome Walking Method
2.5. In Silico Analysis of Promoter
2.6. Expression Analysis of EaPHT1;2 Promoters in Tobacco Transgenic Events
2.7. EaPHT1;2 Promoter Activity in Pi Stress
2.8. Relative GUS Expression Analysis of the EaPHT1;2 Promoter Using RT-PCR
3. Discussion
4. Materials and Methods
4.1. Bacteria and Plant Materials
4.2. EaPHT1;2 Gene Isolation and Cloning
4.3. Bioinformatics Characterization of the EaPHT1;2 Gene
4.4. Isolation of EaPHT1;2 Promoter Region Using a Genome Walking Method
4.5. Bioinformatics Analysis of the Promoter Regions
4.6. Vector Construction and Tobacco Transformation
4.7. Low- and High-Pi Treatments
4.8. Histochemical and Fluorometric β-Glucuronidase (GUS) Assays
4.9. qRT-PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gu, M.; Chen, A.; Sun, S.; Xu, G. Complex Regulation of Plant Phosphate Transporters and the Gap between Molecular Mechanisms and Practical Application: What Is Missing? Mol. Plant 2016, 9, 396–416. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Upadhyay, M.K.; Srivastava, A.K.; Abdelrahman, M.; Suprasanna, P.; Tran, L.-S.P. Cellular and Subcellular Phosphate Transport Machinery in Plants. Int. J. Mol. Sci. 2018, 19, 1914. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, F.; Lu, H.; Liu, Y.; Mao, C. Phosphate Uptake and Transport in Plants: An Elaborate Regulatory System. Plant Cell Physiol. 2021, 62, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Crombez, H.; Motte, H.; Beeckman, T. Tackling Plant Phosphate Starvation by the Roots. Dev. Cell 2019, 48, 599–615. [Google Scholar] [CrossRef] [PubMed]
- Vilaça De Vasconcelos, M.J.; Fontes Figueiredo, J.E.; De Oliveira, M.F.; Schaffert, R.E.; Raghothama, K.G. Plant Phosphorus Use Efficiency in Acid Tropical Soil. RBMS 2022, 21, e1259. [Google Scholar] [CrossRef]
- Balemi, T.; Negisho, K. Management of Soil Phosphorus and Plant Adaptation Mechanisms to Phosphorus Stress for Sustainable Crop Production: A Review. J. Soil Sci. Plant Nutr. 2012, 12, 547–562. [Google Scholar] [CrossRef]
- Akpinar, B.A.; Avsar, B.; Lucas, S.J.; Budak, H. Plant Abiotic Stress Signaling. Plant Signal Behav. 2012, 7, 1450–1455. [Google Scholar] [CrossRef]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic Stress and Reactive Oxygen Species: Generation, Signaling, and Defense Mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [CrossRef]
- Satheesh, V.; Tahir, A.; Li, J.; Lei, M. Plant Phosphate Nutrition: Sensing the Stress. Stress. Biol. 2022, 2, 16. [Google Scholar] [CrossRef]
- Yu, X.; Keitel, C.; Dijkstra, F.A. Global Analysis of Phosphorus Fertilizer Use Efficiency in Cereal Crops. Glob. Food Secur. 2021, 29, 100545. [Google Scholar] [CrossRef]
- Elbasiouny, H.; El-Ramady, H.; Elbehiry, F.; Rajput, V.D.; Minkina, T.; Mandzhieva, S. Plant Nutrition under Climate Change and Soil Carbon Sequestration. Sustainability 2022, 14, 914. [Google Scholar] [CrossRef]
- Bechtaoui, N.; Rabiu, M.K.; Raklami, A.; Oufdou, K.; Hafidi, M.; Jemo, M. Phosphate-Dependent Regulation of Growth and Stresses Management in Plants. Front. Plant Sci. 2021, 12, 679916. [Google Scholar] [CrossRef] [PubMed]
- Nussaume, L.; Kanno, S.; Javot, H.; Marin, E.; Nakanishi, T.M.; Thibaud, M.-C. Phosphate Import in Plants: Focus on the PHT1 Transporters. Front. Plant Sci. 2011, 2, 83. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hu, L.; Yu, D.; Xu, K.; Zhang, J.; Li, X.; Wang, P.; Chen, G.; Liu, Z.; Peng, C.; et al. Integrative Analysis of the Wheat PHT1 Gene Family Reveals A Novel Member Involved in Arbuscular Mycorrhizal Phosphate Transport and Immunity. Cells 2019, 8, 490. [Google Scholar] [CrossRef] [PubMed]
- Rae, A.L.; Cybinski, D.H.; Jarmey, J.M.; Smith, F.W. Characterization of Two Phosphate Transporters from Barley; Evidence for Diverse Function and Kinetic Properties among Members of the Pht1 Family. Plant Mol. Biol. 2003, 53, 27–36. [Google Scholar] [CrossRef]
- Liu, F.; Chang, X.-J.; Ye, Y.; Xie, W.-B.; Wu, P.; Lian, X.-M. Comprehensive Sequence and Whole-Life-Cycle Expression Profile Analysis of the Phosphate Transporter Gene Family in Rice. Mol. Plant 2011, 4, 1105–1122. [Google Scholar] [CrossRef]
- Fan, C.; Wang, X.; Hu, R.; Wang, Y.; Xiao, C.; Jiang, Y.; Zhang, X.; Zheng, C.; Fu, Y.-F. The Pattern of Phosphate transporter 1 Genes Evolutionary Divergence in Glycine maxL. BMC Plant Biol. 2013, 13, 48. [Google Scholar] [CrossRef]
- Ceasar, S.A.; Hodge, A.; Baker, A.; Baldwin, S.A. Phosphate Concentration and Arbuscular Mycorrhizal Colonisation Influence the Growth, Yield and Expression of Twelve PHT1 Family Phosphate Transporters in Foxtail Millet (Setaria italica). PLoS ONE 2014, 9, e108459. [Google Scholar] [CrossRef]
- Chen, A.; Chen, X.; Wang, H.; Liao, D.; Gu, M.; Qu, H.; Sun, S.; Xu, G. Genome-Wide Investigation and Expression Analysis Suggest Diverse Roles and Genetic Redundancy of Pht1 Family Genes in Response to Pi Deficiency in Tomato. BMC Plant Biol. 2014, 14, 61. [Google Scholar] [CrossRef]
- Zhang, C.; Meng, S.; Li, M.; Zhao, Z. Genomic Identification and Expression Analysis of the Phosphate Transporter Gene Family in Poplar. Front. Plant Sci. 2016, 7, 1398. [Google Scholar] [CrossRef]
- Sun, T.; Li, M.; Shao, Y.; Yu, L.; Ma, F. Comprehensive Genomic Identification and Expression Analysis of the Phosphate Transporter (PHT) Gene Family in Apple. Front. Plant Sci. 2017, 8, 426. [Google Scholar] [CrossRef] [PubMed]
- Teng, W.; Zhao, Y.-Y.; Zhao, X.-Q.; He, X.; Ma, W.-Y.; Deng, Y.; Chen, X.-P.; Tong, Y.-P. Genome-Wide Identification, Characterization, and Expression Analysis of PHT1 Phosphate Transporters in Wheat. Front. Plant Sci. 2017, 8, 543. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yang, Y.; Liao, L.; Xu, J.; Liang, X.; Liu, W. Genome-Wide Identification and Functional Characterization of the Phosphate Transporter Gene Family in Sorghum. Biomolecules 2019, 9, 670. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Lu, M.; Zhang, C.; Qu, X.; Liu, Y.; Yang, J.; Yuan, J. Isolation and Functional Characterisation of the PHT1 Gene Encoding a High-Affinity Phosphate Transporter in Camellia oleifera. J. Hortic. Sci. Biotechnol. 2020, 95, 553–564. [Google Scholar] [CrossRef]
- Lhamo, D.; Shao, Q.; Tang, R.; Luan, S. Genome-Wide Analysis of the Five Phosphate Transporter Families in Camelina sativa and Their Expressions in Response to Low-P. Int. J. Mol. Sci. 2020, 21, 8365. [Google Scholar] [CrossRef]
- Cao, Y.; Liu, J.; Li, Y.; Zhang, J.; Li, S.; An, Y.; Hu, T.; Yang, P. Functional Analysis of the Phosphate Transporter Gene MtPT6 From Medicago truncatula. Front. Plant Sci. 2021, 11, 620377. [Google Scholar] [CrossRef]
- Rubio, V.; Linhares, F.; Solano, R.; Martín, A.C.; Iglesias, J.; Leyva, A.; Paz-Ares, J. A Conserved MYB Transcription Factor Involved in Phosphate Starvation Signaling Both in Vascular Plants and in Unicellular Algae. Genes Dev. 2001, 15, 2122–2133. [Google Scholar] [CrossRef]
- Zhou, J.; Jiao, F.; Wu, Z.; Li, Y.; Wang, X.; He, X.; Zhong, W.; Wu, P. OsPHR2 Is Involved in Phosphate-Starvation Signaling and Excessive Phosphate Accumulation in Shoots of Plants. Plant Physiol. 2008, 146, 1673–1686. [Google Scholar] [CrossRef]
- Bustos, R.; Castrillo, G.; Linhares, F.; Puga, M.I.; Rubio, V.; Pérez-Pérez, J.; Solano, R.; Leyva, A.; Paz-Ares, J. A Central Regulatory System Largely Controls Transcriptional Activation and Repression Responses to Phosphate Starvation in Arabidopsis. PLoS Genet. 2010, 6, e1001102. [Google Scholar] [CrossRef]
- Sega, P.; Pacak, A. Plant PHR Transcription Factors: Put on A Map. Genes 2019, 10, 1018. [Google Scholar] [CrossRef]
- Sega, P.; Kruszka, K.; Bielewicz, D.; Karlowski, W.; Nuc, P.; Szweykowska-Kulinska, Z.; Pacak, A. Pi-Starvation Induced Transcriptional Changes in Barley Revealed by a Comprehensive RNA-Seq and Degradome Analyses. BMC Genom. 2021, 22, 165. [Google Scholar] [CrossRef]
- Yi, K.; Wu, Z.; Zhou, J.; Du, L.; Guo, L.; Wu, Y.; Wu, P. OsPTF1, a Novel Transcription Factor Involved in Tolerance to Phosphate Starvation in Rice. Plant Physiol. 2005, 138, 2087–2096. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-F.; Li, L.-Q.; Xu, Q.; Kong, Y.-H.; Wang, H.; Wu, W.-H. The WRKY6 Transcription Factor Modulates PHOSPHATE1 Expression in Response to Low Pi Stress in Arabidopsis. Plant Cell 2009, 21, 3554–3566. [Google Scholar] [CrossRef]
- Karthikeyan, A.S.; Varadarajan, D.K.; Mukatira, U.T.; D’Urzo, M.P.; Damsz, B.; Raghothama, K.G. Regulated Expression of Arabidopsis Phosphate Transporters. Plant Physiol. 2002, 130, 221. [Google Scholar] [CrossRef] [PubMed]
- Baek, D.; Chun, H.J.; Yun, D.-J.; Kim, M.C. Cross-Talk between Phosphate Starvation and Other Environmental Stress Signaling Pathways in Plants. Mol. Cells 2017, 40, 697–705. [Google Scholar] [CrossRef]
- Yong-Villalobos, L.; Cervantes-Pérez, S.A.; Gutiérrez-Alanis, D.; Gonzáles-Morales, S.; Martínez, O.; Herrera-Estrella, L. Phosphate Starvation Induces DNA Methylation in the Vicinity of Cis-Acting Elements Known to Regulate the Expression of Phosphate–Responsive Genes. Plant Signal. Behav. 2016, 11, e1173300. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, X.; Zhang, H.; Wang, S.; Ye, X.; Shi, L.; Xu, F.; Ding, G. Molecular Identification of the Phosphate Transporter Family 1 (PHT1) Genes and Their Expression Profiles in Response to Phosphorus Deprivation and Other Abiotic Stresses in Brassica napus. PLoS ONE 2019, 14, e0220374. [Google Scholar] [CrossRef] [PubMed]
- Desalegn, B.; Kebede, E.; Legesse, H.; Fite, T. Sugarcane Productivity and Sugar Yield Improvement: Selecting Variety, Nitrogen Fertilizer Rate, and Bioregulator as a First-Line Treatment. Heliyon 2023, 9, e15520. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, G.C.; Rosa, P.A.L.; Jalal, A.; Oliveira, C.E.d.S.; Galindo, F.S.; Viana, R.d.S.; De Carvalho, P.H.G.; da Silva, E.C.; Nogueira, T.A.R.; Al-Askar, A.A.; et al. Technological Quality of Sugarcane Inoculated with Plant-Growth-Promoting Bacteria and Residual Effect of Phosphorus Rates. Plants 2023, 12, 2699. [Google Scholar] [CrossRef]
- Soltangheisi, A.; Withers, P.J.A.; Pavinato, P.S.; Cherubin, M.R.; Rossetto, R.; Do Carmo, J.B.; da Rocha, G.C.; Martinelli, L.A. Improving Phosphorus Sustainability of Sugarcane Production in Brazil. GCB Bioenergy 2019, 11, 1444–1455. [Google Scholar] [CrossRef]
- Murugan, N.; Palanisamy, V.; Channappa, M.; Ramanathan, V.; Ramaswamy, M.; Govindakurup, H.; Chinnaswamy, A. Genome-Wide In Silico Identification, Structural Analysis, Promoter Analysis, and Expression Profiling of PHT Gene Family in Sugarcane Root under Salinity Stress. Sustainability 2022, 14, 15893. [Google Scholar] [CrossRef]
- Murugan, N.; Kumar, R.; Pandey, S.K.; Dhansu, P.; Chennappa, M.; Nallusamy, S.; Govindakurup, H.; Chinnaswamy, A. In Silico Dissection of Regulatory Regions of PHT Genes from Saccharum Spp. Hybrid and Sorghum Bicolor and Expression Analysis of PHT Promoters under Osmotic Stress Conditions in Tobacco. Sustainability 2023, 15, 1048. [Google Scholar] [CrossRef]
- Ren, F.; Zhao, C.-Z.; Liu, C.-S.; Huang, K.-L.; Guo, Q.-Q.; Chang, L.-L.; Xiong, H.; Li, X.-B. A Brassica Napus PHT1 Phosphate Transporter, BnPht1;4, Promotes Phosphate Uptake and Affects Roots Architecture of Transgenic Arabidopsis. Plant Mol. Biol. 2014, 86, 595–607. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Li, G.; Sun, Z.; Chen, Y.; Guo, W.; Li, Y.; Chen, Y.; Yang, J.; Hou, H. Identification, Structure Analysis, and Transcript Profiling of Phosphate Transporters under Pi Deficiency in Duckweeds. Int. J. Biol. Macromol. 2021, 188, 595–608. [Google Scholar] [CrossRef]
- Karandashov, V.; Bucher, M. Symbiotic Phosphate Transport in Arbuscular Mycorrhizas. Trends Plant Sci. 2005, 10, 22–29. [Google Scholar] [CrossRef]
- Chen, A.; Hu, J.; Sun, S.; Xu, G. Conservation and Divergence of Both Phosphate- and Mycorrhiza-Regulated Physiological Responses and Expression Patterns of Phosphate Transporters in Solanaceous Species. New Phytol. 2007, 173, 817–831. [Google Scholar] [CrossRef]
- Cao, G.-H.; Li, Z.-D.; Wang, X.-F.; Zhang, X.; Zhao, R.-H.; Gu, W.; Chen, D.; Yu, J.; He, S. Phosphate Transporters, PnPht1;1 and PnPht1;2 from Panax Notoginseng Enhance Phosphate and Arsenate Acquisition. BMC Plant Biol. 2020, 20, 124. [Google Scholar] [CrossRef]
- Wu, Z.; Zhao, J.; Gao, R.; Hu, G.; Gai, J.; Xu, G.; Xing, H. Molecular Cloning, Characterization and Expression Analysis of Two Members of the Pht1 Family of Phosphate Transporters in Glycine Max. PLoS ONE 2011, 6, e19752. [Google Scholar] [CrossRef]
- Liu, C.; Su, J.; Stephen, G.K.; Wang, H.; Song, A.; Chen, F.; Zhu, Y.; Chen, S.; Jiang, J. Overexpression of Phosphate Transporter Gene CmPht1;2 Facilitated Pi Uptake and Alternated the Metabolic Profiles of Chrysanthemum Under Phosphate Deficiency. Front. Plant Sci. 2018, 9, 686. [Google Scholar] [CrossRef]
- Pedersen, B.P.; Kumar, H.; Waight, A.B.; Risenmay, A.J.; Roe-Zurz, Z.; Chau, B.H.; Schlessinger, A.; Bonomi, M.; Harries, W.; Sali, A.; et al. Crystal Structure of a Eukaryotic Phosphate Transporter. Nature 2013, 496, 533–536. [Google Scholar] [CrossRef]
- Kaur, A.; Pati, P.K.; Pati, A.M.; Nagpal, A.K. In-Silico Analysis of Cis-Acting Regulatory Elements of Pathogenesis-Related Proteins of Arabidopsis Thaliana and Oryza Sativa. PLoS ONE 2017, 12, e0184523. [Google Scholar] [CrossRef] [PubMed]
- Lata, C.; Prasad, M. Role of DREBs in Regulation of Abiotic Stress Responses in Plants. J. Exp. Bot. 2011, 62, 4731–4748. [Google Scholar] [CrossRef] [PubMed]
- Tittarelli, A.; Milla, L.; Vargas, F.; Morales, A.; Neupert, C.; Meisel, L.A.; Salvo-G, H.; Peñaloza, E.; Muñoz, G.; Corcuera, L.J.; et al. Isolation and Comparative Analysis of the Wheat TaPT2 Promoter: Identification in silico of New Putative Regulatory Motifs Conserved between Monocots and Dicots. J. Exp. Bot. 2007, 58, 2573–2582. [Google Scholar] [CrossRef] [PubMed]
- Sobkowiak, L.; Bielewicz, D.; Malecka, E.; Jakobsen, I.; Albrechtsen, M.; Pacak, A. The Role of the P1BS Element Containing Promoter-Driven Genes in Pi Transport and Homeostasis in Plants. Front. Plant Sci. 2012, 3, 58. [Google Scholar] [CrossRef] [PubMed]
- Schünmann, P.H.D.; Richardson, A.E.; Smith, F.W.; Delhaize, E. Characterization of Promoter Expression Patterns Derived from the Pht1 Phosphate Transporter Genes of Barley (Hordeum vulgare L.). J. Exp. Bot. 2004, 55, 855–865. [Google Scholar] [CrossRef]
- Koyama, T.; Ono, T.; Shimizu, M.; Jinbo, T.; Mizuno, R.; Tomita, K.; Mitsukawa, N.; Kawazu, T.; Kimura, T.; Ohmiya, K.; et al. Promoter of Arabidopsis Thaliana Phosphate Transporter Gene Drives Root-Specific Expression of Transgene in Rice. J. Biosci. Bioeng. 2005, 99, 38–42. [Google Scholar] [CrossRef]
- Chen, A.; Gu, M.; Sun, S.; Zhu, L.; Hong, S.; Xu, G. Identification of Two Conserved Cis-Acting Elements, MYCS and P1BS, Involved in the Regulation of Mycorrhiza-Activated Phosphate Transporters in Eudicot Species. New Phytol. 2011, 189, 1157–1169. [Google Scholar] [CrossRef]
- Li, L.H.; Guo, N.; Wu, Z.Y.; Zhao, J.M.; Sun, J.T.; Wang, X.T.; Xing, H. P1BS, a Conserved Motif Involved in Tolerance to Phosphate Starvation in Soybean. Genet. Mol. Res. 2015, 14, 9384–9394. [Google Scholar] [CrossRef]
- Peñaloza, E.; Santiago, M.; Cabrera, S.; Muñoz, G.; Corcuera, L.J.; Silva, H. Characterization of the High-Affinity Phosphate Transporter PHT1;4 Gene Promoter of Arabidopsis Thaliana in Transgenic Wheat. Biol. Plant. 2017, 61, 453–462. [Google Scholar] [CrossRef]
- Zhao, Q.; Luo, Z.; Chen, J.; Jia, H.; Ai, P.; Chen, A.; Li, Y.; Xu, G. Characterization of Two Cis-Acting Elements, P1BS and W-Box, in the Regulation of OsPT6 Responsive to Phosphors Deficiency. Plant Growth Regul. 2021, 93, 303–310. [Google Scholar] [CrossRef]
- Miao, J.; Sun, J.; Liu, D.; Li, B.; Zhang, A.; Li, Z.; Tong, Y. Characterization of the Promoter of Phosphate Transporter TaPHT1.2 Differentially Expressed in Wheat Varieties. J. Genet. Genom. 2009, 36, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Mudge, S.R.; Smith, F.W.; Richardson, A.E. Root-Specific and Phosphate-Regulated Expression of Phytase under the Control of a Phosphate Transporter Promoter Enables Arabidopsis to Grow on Phytate as a Sole P Source. Plant Sci. 2003, 165, 871–878. [Google Scholar] [CrossRef]
- Hou, J.; Jiang, P.; Qi, S.; Zhang, K.; He, Q.; Xu, C.; Ding, Z.; Zhang, K.; Li, K. Isolation and Functional Validation of Salinity and Osmotic Stress Inducible Promoter from the Maize Type-II H+-Pyrophosphatase Gene by Deletion Analysis in Transgenic Tobacco Plants. PLoS ONE 2016, 11, e0154041. [Google Scholar] [CrossRef]
- Zhang, H.; Jing, R.; Mao, X. Functional Characterization of TaSnRK2.8 Promoter in Response to Abiotic Stresses by Deletion Analysis in Transgenic Arabidopsis. Front. Plant Sci. 2017, 8, 1198. [Google Scholar] [CrossRef] [PubMed]
- Schünmann, P.H.D.; Richardson, A.E.; Vickers, C.E.; Delhaize, E. Promoter Analysis of the Barley Pht1;1 Phosphate Transporter Gene Identifies Regions Controlling Root Expression and Responsiveness to Phosphate Deprivation. Plant Physiol. 2004, 136, 4205–4214. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Liu, B.; Zhao, M.; Zhang, K.; He, Q.; Zhao, X.; Cheng, W.; Ding, Z.; Zhang, K.; Li, K. Isolation and Characterization of a 295-Bp Strong Promoter of Maize High-Affinity Phosphate Transporter Gene ZmPht1; 5 in Transgenic Nicotiana Benthamiana and Zea Mays. Planta 2020, 251, 106. [Google Scholar] [CrossRef]
- Kasirajan, L.; Valiyaparambth, R.; Kamaraj, K.; Sebastiar, S.; Hoang, N.V.; Athiappan, S.; Srinivasavedantham, V.; Subramanian, K. Deep Sequencing of Suppression Subtractive Library Identifies Differentially Expressed Transcripts of Saccharum Spontaneum Exposed to Salinity Stress. Physiol. Plant. 2022, 174, e13645. [Google Scholar] [CrossRef]
- Horsch, R.B.; Fry, J.E.; Hoffmann, N.L.; Wallroth, M.; Eichholtz, D.; Rogers, S.G.; Fraley, R.T. A Simple and General Method for Transferring Genes into Plants. Science 1985, 227, 1229–1231. [Google Scholar] [CrossRef]
- Doyle, J. DNA Protocols for Plants. In Molecular Techniques in Taxonomy; Hewitt, G.M., Johnston, A.W.B., Young, J.P.W., Eds.; NATO ASI Series; Springer: Berlin/Heidelberg, Germany, 1991; pp. 283–293. ISBN 978-3-642-83962-7. [Google Scholar]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Springer Protocols Handbooks; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. ISBN 978-1-59259-890-8. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent Updates, New Developments and Status in 2020. Nucleic Acids Res. 2021, 49, D458–D460. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The Protein Families Database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [PubMed]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L.; et al. InterPro in 2022. Nucleic Acids Res. 2023, 51, D418–D427. [Google Scholar] [CrossRef] [PubMed]
- Hulo, N.; Bairoch, A.; Bulliard, V.; Cerutti, L.; De Castro, E.; Langendijk-Genevaux, P.S.; Pagni, M.; Sigrist, C.J.A. The PROSITE Database. Nucleic Acids Res. 2006, 34, D227–D230. [Google Scholar] [CrossRef] [PubMed]
- Sonnhammer, E.L.; von Heijne, G.; Krogh, A. A Hidden Markov Model for Predicting Transmembrane Helices in Protein Sequences. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1998, 6, 175–182. [Google Scholar]
- Geourjon, C.; Deléage, G. SOPMA: Significant Improvements in Protein Secondary Structure Prediction by Consensus Prediction from Multiple Alignments. Bioinformatics 1995, 11, 681–684. [Google Scholar] [CrossRef]
- McGuffin, L.J.; Bryson, K.; Jones, D.T. The PSIPRED Protein Structure Prediction Server. Bioinformatics 2000, 16, 404–405. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 Web Portal for Protein Modeling, Prediction and Analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef]
- Wiederstein, M.; Sippl, M.J. ProSA-Web: Interactive Web Service for the Recognition of Errors in Three-Dimensional Structures of Proteins. Nucleic Acids Res. 2007, 35, W407–W410. [Google Scholar] [CrossRef]
- Jensen, L.J.; Kuhn, M.; Stark, M.; Chaffron, S.; Creevey, C.; Muller, J.; Doerks, T.; Julien, P.; Roth, A.; Simonovic, M.; et al. STRING 8—A Global View on Proteins and Their Functional Interactions in 630 Organisms. Nucleic Acids Res. 2009, 37, D412–D416. [Google Scholar] [CrossRef] [PubMed]
- Almagro Armenteros, J.J.; Sønderby, C.K.; Sønderby, S.K.; Nielsen, H.; Winther, O. DeepLoc: Prediction of Protein Subcellular Localization Using Deep Learning. Bioinformatics 2017, 33, 3387–3395. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating Signal Peptides from Transmembrane Regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Blom, N.; Sicheritz-Pontén, T.; Gupta, R.; Gammeltoft, S.; Brunak, S. Prediction of Post-Translational Glycosylation and Phosphorylation of Proteins from the Amino Acid Sequence. Proteomics 2004, 4, 1633–1649. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Brunak, S. Prediction of Glycosylation across the Human Proteome and the Correlation to Protein Function. Pac. Symp. Biocomput. 2002, 310–322. [Google Scholar]
- Potenza, C.; Aleman, L.; Sengupta-Gopalan, C. Targeting Transgene Expression in Research, Agricultural, and Environmental Applications: Promoters Used in Plant Transformation. In Vitro Cell. Dev. Biol.-Plant 2004, 40, 1–22. [Google Scholar] [CrossRef]
- Ahmadi, F.; Akmar Abdullah, S.N.; Kadkhodaei, S.; Ijab, S.M.; Hamzah, L.; Aziz, M.A.; Rahman, Z.A.; Rabiah Syed Alwee, S.S. Functional Characterization of the Gene Promoter for an Elaeis Guineensis Phosphate Starvation-Inducible, High Affinity Phosphate Transporter in Both Homologous and Heterologous Model Systems. Plant Physiol. Biochem. 2018, 127, 320–335. [Google Scholar] [CrossRef]
- Mohan, C. Isolation and Characterization of Constitutive and Wound Inducible Promoters and Validation of Designed Synthetic Stem/Root Specific Promoters for Sugarcane Transformation. 2015. Available online: https://www.researchgate.net/profile/Chakravarthi-Mohan/publication/304213264_Isolation_and_characterization_of_constitutive_and_wound_inducible_promoters_and_validation_of_designed_synthetic_stemroot_specific_promoters_for_sugarcane_transformation/links/576974c808ae1a43d23a3461/Isolation-and-characterization-of-constitutive-and-wound-inducible-promoters-and-validation-of-designed-synthetic-stem-root-specific-promoters-for-sugarcane-transformation.pdf (accessed on 12 March 2023).
- Reese, M.G. Application of a Time-Delay Neural Network to Promoter Annotation in the Drosophila Melanogaster Genome. Comput. Chem. 2001, 26, 51–56. [Google Scholar] [CrossRef]
- Shahmuradov, I.A.; Umarov, R.K.; Solovyev, V.V. TSSPlant: A New Tool for Prediction of Plant Pol II Promoters. Nucleic Acids Res. 2017, 45, e65. [Google Scholar] [CrossRef]
- Chow, C.-N.; Lee, T.-Y.; Hung, Y.-C.; Li, G.-Z.; Tseng, K.-C.; Liu, Y.-H.; Kuo, P.-L.; Zheng, H.-Q.; Chang, W.-C. PlantPAN3.0: A New and Updated Resource for Reconstructing Transcriptional Regulatory Networks from ChIP-Seq Experiments in Plants. Nucleic Acids Res. 2019, 47, D1155–D1163. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME Suite: Tools for Motif Discovery and Searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant Cis-Acting Regulatory DNA Elements (PLACE) Database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for in Silico Analysis of Promoter Sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Aquino, S.; Carneiro, F.; Rêgo, E.; Alves, G.; Andrade, A.; Marraccini, P. Functional Analysis of Different Promoter Haplotypes of the Coffee (Coffea Canephora) CcDREB1D Gene through Genetic Transformation of Nicotiana tabacum. Plant Cell Tissue Organ Cult. (PCTOC) 2018, 132, 1–16. [Google Scholar] [CrossRef]
- Remy, E.; Cabrito, T.R.; Batista, R.A.; Teixeira, M.C.; Sá-Correia, I.; Duque, P. The Pht1;9 and Pht1;8 Transporters Mediate Inorganic Phosphate Acquisition by the Arabidopsis Thaliana Root during Phosphorus Starvation. New Phytol. 2012, 195, 356–371. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS Fusions: Beta-Glucuronidase as a Sensitive and Versatile Gene Fusion Marker in Higher Plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. The Single-Step Method of RNA Isolation by Acid Guanidinium Thiocyanate–Phenol–Chloroform Extraction: Twenty-Something Years On. Nat. Protoc. 2006, 1, 581–585. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Selvarajan, D.; Mohan, C.; Dhandapani, V.; Nerkar, G.; Jayanarayanan, A.N.; Vadakkancherry Mohanan, M.; Murugan, N.; Kaur, L.; Chennappa, M.; Kumar, R.; et al. Differential Gene Expression Profiling through Transcriptome Approach of Saccharum spontaneum L. under Low Temperature Stress Reveals Genes Potentially Involved in Cold Acclimation. 3 Biotech 2018, 8, 195. [Google Scholar] [CrossRef]
- Manoj, V.M.; Anunanthini, P.; Swathik, P.C.; Dharshini, S.; Ashwin Narayan, J.; Manickavasagam, M.; Sathishkumar, R.; Suresha, G.S.; Hemaprabha, G.; Ram, B.; et al. Comparative Analysis of Glyoxalase Pathway Genes in Erianthus Arundinaceus and Commercial Sugarcane Hybrid under Salinity and Drought Conditions. BMC Genom. 2019, 19, 986. [Google Scholar] [CrossRef] [PubMed]
- Dharshini, S.; Manoj, V.M.; Suresha, G.S.; Narayan, J.A.; Padmanabhan, T.S.S.; Kumar, R.; Meena, M.R.; Manickavasagam, M.; Ram, B.; Appunu, C. Isolation and Characterization of Nuclear Localized Abiotic Stress Responsive Cold Regulated Gene 413 (SsCor413) from Saccharum Spontaneum. Plant Mol. Biol. Rep. 2020, 38, 628–640. [Google Scholar] [CrossRef]
- Narayan, J.A.; Dharshini, S.; Manoj, V.M.; Padmanabhan, T.S.S.; Kadirvelu, K.; Suresha, G.S.; Subramonian, N.; Ram, B.; Premachandran, M.N.; Appunu, C. Isolation and Characterization of Water-Deficit Stress-Responsive α-Expansin 1 (EXPA1) Gene from Saccharum Complex. 3 Biotech 2019, 9, 186. [Google Scholar] [CrossRef] [PubMed]
- Ashwin Narayan, J.; Chakravarthi, M.; Nerkar, G.; Manoj, V.M.; Dharshini, S.; Subramonian, N.; Premachandran, M.N.; Arun Kumar, R.; Krishna Surendar, K.; Hemaprabha, G.; et al. Overexpression of Expansin EaEXPA1, a Cell Wall Loosening Protein Enhances Drought Tolerance in Sugarcane. Ind. Crops Prod. 2021, 159, 113035. [Google Scholar] [CrossRef]
- Narayan, A.; Manoj, V.M.; Nerkar, G.; Mohan, C.; Dharshini, S.; Subramonian, N.; Premachandran, M.; Valarmathi, R.; Kumar, R.; Gomathi, R.; et al. Transgenic Sugarcane with Higher Levels of BRK1 Showed Improved Drought Tolerance. Plant Cell Rep. 2023, 42, 1–18. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naveenarani, M.; Swamy, H.K.M.; Surya Krishna, S.; Mahadevaiah, C.; Valarmathi, R.; Manickavasagam, M.; Arun, M.; Hemaprabha, G.; Appunu, C. Isolation and Characterization of Erianthus arundinaceus Phosphate Transporter 1 (PHT1) Gene Promoter and 5′ Deletion Analysis of Transcriptional Regulation Regions under Phosphate Stress in Transgenic Tobacco. Plants 2023, 12, 3760. https://doi.org/10.3390/plants12213760
Naveenarani M, Swamy HKM, Surya Krishna S, Mahadevaiah C, Valarmathi R, Manickavasagam M, Arun M, Hemaprabha G, Appunu C. Isolation and Characterization of Erianthus arundinaceus Phosphate Transporter 1 (PHT1) Gene Promoter and 5′ Deletion Analysis of Transcriptional Regulation Regions under Phosphate Stress in Transgenic Tobacco. Plants. 2023; 12(21):3760. https://doi.org/10.3390/plants12213760
Chicago/Turabian StyleNaveenarani, Murugan, Huskur Kumaraswamy Mahadeva Swamy, Sakthivel Surya Krishna, Channappa Mahadevaiah, Ramanathan Valarmathi, Markandan Manickavasagam, Muthukrishnan Arun, Govindakurup Hemaprabha, and Chinnaswamy Appunu. 2023. "Isolation and Characterization of Erianthus arundinaceus Phosphate Transporter 1 (PHT1) Gene Promoter and 5′ Deletion Analysis of Transcriptional Regulation Regions under Phosphate Stress in Transgenic Tobacco" Plants 12, no. 21: 3760. https://doi.org/10.3390/plants12213760
APA StyleNaveenarani, M., Swamy, H. K. M., Surya Krishna, S., Mahadevaiah, C., Valarmathi, R., Manickavasagam, M., Arun, M., Hemaprabha, G., & Appunu, C. (2023). Isolation and Characterization of Erianthus arundinaceus Phosphate Transporter 1 (PHT1) Gene Promoter and 5′ Deletion Analysis of Transcriptional Regulation Regions under Phosphate Stress in Transgenic Tobacco. Plants, 12(21), 3760. https://doi.org/10.3390/plants12213760