
Effects of Vegetation Types and Soil Properties on Regional Soil Carbon and Nitrogen in Salinized Reservoir Wetland, Northeast China

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Spatial Variations in SOC, TN, SMBC, SMBN and Other Soil Properties
2.2. Correlation Analysis between Soil Physicochemical Properties
2.3. Relationships between SOC, TN, SMBC, SMBN and Other Soil Properties
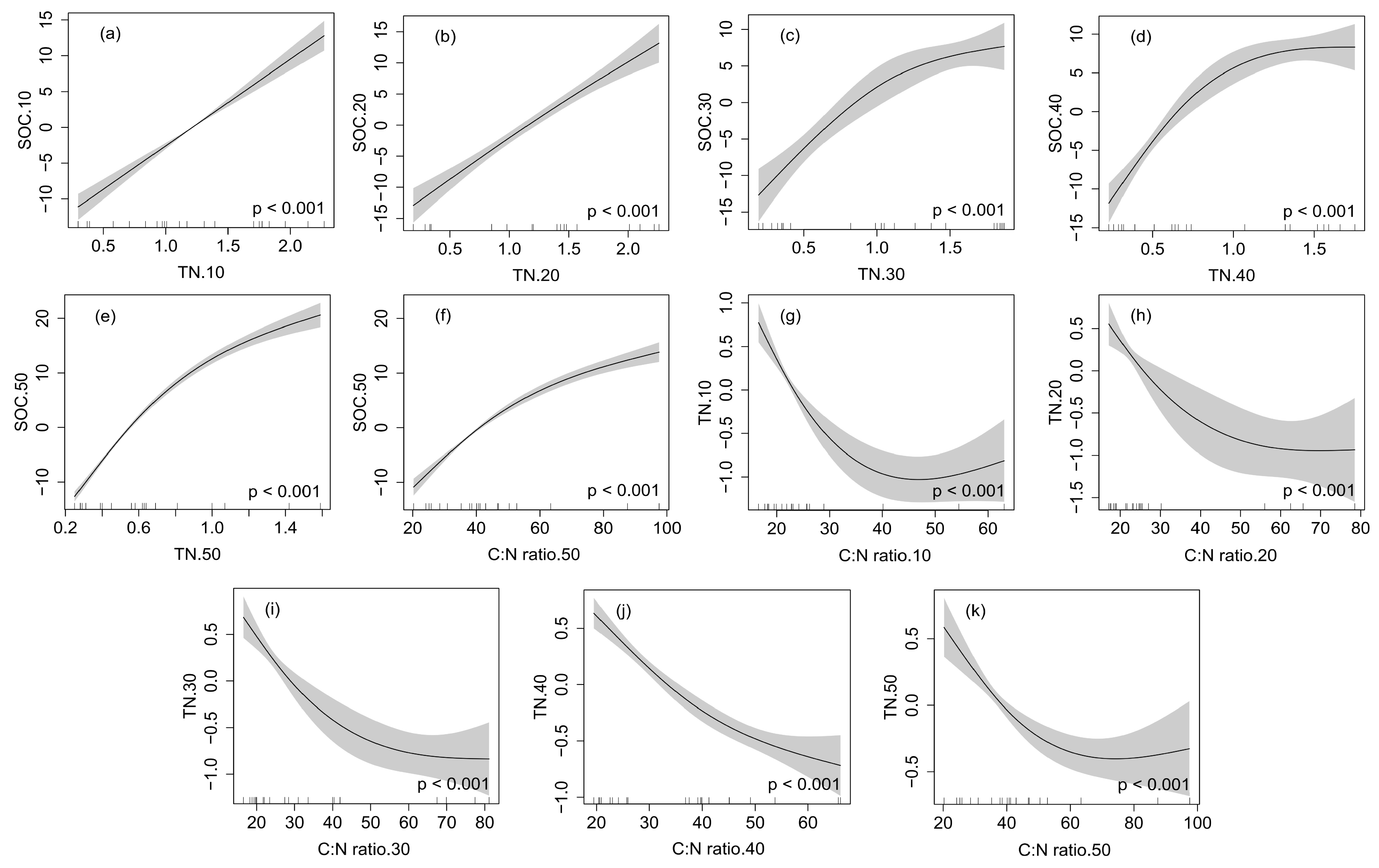
2.4. The GAM for Analyzing Correlations between SOC, TN and Other Soil Physicochemical Properties in Different Soil Depths
2.5. The Effects of Vegetation Type and Soil Depth on SOC, TN, SMBC and SMBN
2.6. Total SOC, TN, SMBC and SMBN Storages
3. Discussion
3.1. The Spatial Variations in Soil Properties in Different Vegetation Type and Soil Depth
3.2. The Relationships between SOC, TN, SMBC, SMBN and Other Soil Properties
4. Materials and Methods
4.1. Study Area
4.2. Soil Sampling and Laboratory Analysis
4.3. Determination of Aboveground Vegetation Biomass
4.4. Calculations of Total Carbon and Nitrogen
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yao, X.; Yu, K.; Deng, Y.; Liu, J.; Lai, Z. Spatial variability of soil organic carbon and total nitrogen in the hilly red soil region of Southern China. J. For. Res. 2020, 31, 2385–2394. [Google Scholar] [CrossRef]
- Yang, M.; Geng, X.; Grace, J.; Lu, C.; Zhu, Y.; Zhou, Y.; Lei, G. Spatial and seasonal CH4 flux in the littoral zone of Miyun Reservoir near Beijing: The effects of water level and its fluctuation. PLoS ONE 2014, 9, e94275. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yang, R.; Bai, Z. Spatial variability and sampling optimization of soil organic carbon and total nitrogen for Minesoils of the Loess Plateau using geostatistics. Ecol. Eng. 2015, 82, 159–164. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Zhang, L.; Anderson, C.J.; Altor, A.E.; Hernández, M.E. Creating riverine wetlands: Ecological succession, nutrient retention, and pulsing effects. Ecol. Eng. 2005, 25, 510–527. [Google Scholar] [CrossRef]
- Reddy, K.R.; DeLaune, R.D. Biogeochemistry of Wetlands: Science and Applications, 1st ed.; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Wang, J.; Bai, J.; Zhao, Q.; Lu, Q.; Xia, Z. Five-year changes in soil organic carbon and total nitrogen in coastal wetlands affected by flow-sediment regulation in a Chinese delta. Sci. Rep. 2016, 6, 21137. [Google Scholar] [CrossRef] [PubMed]
- Bohn, H.L. Estimate of organic carbon in world soils. Soil Sci. Soc. Am. J. 1976, 40, 468–470. [Google Scholar] [CrossRef]
- Wildi, W. Environmental hazards of dams and reservoirs. Near Curric. Nat. Environ. Sci. 2010, 2, 187–197. [Google Scholar]
- Li, S.; Lu, X. Uncertainties of carbon emission from hydroelectric reservoirs. Nat. Hazards 2012, 62, 1343–1345. [Google Scholar] [CrossRef]
- Bridgham, S.D.; Cadillo-Quiroz, H.; Keller, J.K.; Zhuang, Q. Methane emissions from wetlands: Biogeochemical, microbial, and modeling perspectives from local to global scales. Glob. Chang. Biol. 2013, 19, 1325–1346. [Google Scholar] [CrossRef]
- Teng, M.; Zeng, L.; Xiao, W.; Huang, Z.; Zhou, Z.; Yan, Z.; Wang, P. Spatial variability of soil organic carbon in Three Gorges Reservoir area, China. Sci. Total Environ. 2017, 599, 1308–1316. [Google Scholar] [CrossRef]
- Cao, Q.; Wang, H.; Zhang, Y.; Lal, R.; Wang, R.; Ge, X.; Liu, J. Factors affecting distribution patterns of organic carbon in sediments at regional and national scales in China. Sci. Rep. 2017, 7, 5497. [Google Scholar] [CrossRef]
- Li, Y.; Wu, H.; Wang, J.; Cui, L.; Tian, D.; Wang, J.; Zhang, X.; Yan, L.; Yan, Z.; Zhang, K. Plant biomass and soil organic carbon are main factors influencing dry-season ecosystem carbon rates in the coastal zone of the Yellow River Delta. PLoS ONE 2019, 14, e0210768. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Huang, Y.; Sun, F.; Sun, W. A synthesis of soil carbon and nitrogen recovery after wetland restoration and creation in the United States. Sci. Rep. 2017, 7, 7966. [Google Scholar] [CrossRef] [PubMed]
- Craft, C. Freshwater input structures soil properties, vertical accretion, and nutrient accumulation of Georgia and US tidal marshes. Limnol. Oceanogr. 2007, 52, 1220–1230. [Google Scholar] [CrossRef]
- Xu, X.; Thornton, P.E.; Post, W.M. A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems. Glob. Ecol. Biogeogr. 2013, 22, 737–749. [Google Scholar] [CrossRef]
- Bahn, M.; Rodeghiero, M.; Anderson-Dunn, M.; Dore, S.; Gimeno, C.; Drösler, M.; Williams, M.; Ammann, C.; Berninger, F.; Flechard, C. Soil respiration in European grasslands in relation to climate and assimilate supply. Ecosystems 2008, 11, 1352–1367. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, L.; Hu, J.; Zhang, W.; Fu, X.; Le, Y.; Jin, F. Organic carbon accumulation capability of two typical tidal wetland soils in Chongming Dongtan, China. J. Environ. Sci. 2011, 23, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Santín, C.; González-Pérez, M.; Otero, X.; Vidal-Torrado, P.; Macías, F.; Álvarez, M. Characterization of humic substances in salt marsh soils under sea rush (Juncus maritimus). Estuar. Coast. Shelf Sci. 2008, 79, 541–548. [Google Scholar] [CrossRef]
- Xiong, Z.; Li, S.; Yao, L.; Liu, G.; Zhang, Q.; Liu, W. Topography and land use effects on spatial variability of soil denitrification and related soil properties in riparian wetlands. Ecol. Eng. 2015, 83, 437–443. [Google Scholar] [CrossRef]
- Bai, J.; Ouyang, H.; Xiao, R.; Gao, J.; Gao, H.; Cui, B.; Huang, L. Spatial variability of soil carbon, nitrogen, and phosphorus content and storage in an alpine wetland in the Qinghai–Tibet Plateau, China. Soil Res. 2010, 48, 730–736. [Google Scholar] [CrossRef]
- Song, Y.-Q.; Yang, L.-A.; Li, B.; Hu, Y.-M.; Wang, A.-L.; Zhou, W.; Cui, X.-S.; Liu, Y.-L. Spatial prediction of soil organic matter using a hybrid geostatistical model of an extreme learning machine and ordinary kriging. Sustainability 2017, 9, 754. [Google Scholar] [CrossRef]
- Wang, X.; Song, C.; Sun, X.; Wang, J.; Zhang, X.; Mao, R. Soil carbon and nitrogen across wetland types in discontinuous permafrost zone of the Xiao Xing’an Mountains, northeastern China. Catena 2013, 101, 31–37. [Google Scholar] [CrossRef]
- Wang, X.; Xu, L.; Wan, R.; Chen, Y. Seasonal variations of soil microbial biomass within two typical wetland areas along the vegetation gradient of Poyang Lake, China. Catena 2016, 137, 483–493. [Google Scholar] [CrossRef]
- Nair, A.; Ngouajio, M. Soil microbial biomass, functional microbial diversity, and nematode community structure as affected by cover crops and compost in an organic vegetable production system. Appl. Soil Ecol. 2012, 58, 45–55. [Google Scholar] [CrossRef]
- Huang, W.; Chen, Q.; Ren, K.; Chen, K. Vertical distribution and retention mechanism of nitrogen and phosphorus in soils with different macrophytes of a natural river mouth wetland. Environ. Monit. Assess. 2015, 187, 97. [Google Scholar] [CrossRef]
- Brinson, M.M.; Lugo, A.E.; Brown, S. Primary productivity, decomposition and consumer activity in freshwater wetlands. Annu. Rev. Ecol. Syst. 1981, 12, 123–161. [Google Scholar] [CrossRef]
- Partridge, J.W. Persicaria amphibia (L.) Gray (Polygonum amphibium L.). J. Ecol. 2001, 89, 487–501. [Google Scholar] [CrossRef]
- González-Alcaraz, M.; Egea, C.; Jiménez-Cárceles, F.; Párraga, I.; Maria-Cervantes, A.; Delgado, M.; Álvarez-Rogel, J. Storage of organic carbon, nitrogen and phosphorus in the soil–plant system of Phragmites australis stands from a eutrophicated Mediterranean salt marsh. Geoderma 2012, 185, 61–72. [Google Scholar] [CrossRef]
- Zhang, W.-J.; Xiao, H.-A.; Tong, C.-L.; Su, Y.-R.; Xiang, W.-S.; Huang, D.-Y.; Syers, J.K.; Wu, J. Estimating organic carbon storage in temperate wetland profiles in Northeast China. Geoderma 2008, 146, 311–316. [Google Scholar] [CrossRef]
- Gower, S.; Krankina, O.; Olson, R.; Apps, M.; Linder, S.; Wang, C. Net primary production and carbon allocation patterns of boreal forest ecosystems. Ecol. Appl. 2001, 11, 1395–1411. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, G.; Yu, X.; Liu, Y.; Xia, S.; Ya, L.; Hu, B.; Wan, S. Effect of ground water level on the release of carbon, nitrogen and phosphorus during decomposition of Carex. cinerascens Kükenth in the typical seasonal floodplain in dry season. J. Freshw. Ecol. 2019, 34, 305–322. [Google Scholar] [CrossRef]
- Ashraf, M.N.; Hu, C.; Wu, L.; Duan, Y.; Zhang, W.; Aziz, T.; Cai, A.; Abrar, M.M.; Xu, M. Soil and microbial biomass stoichiometry regulate soil organic carbon and nitrogen mineralization in rice-wheat rotation subjected to long-term fertilization. J. Soils Sediments 2020, 20, 3103–3113. [Google Scholar] [CrossRef]
- Xu, Y.; Ding, F.; Gao, X.; Wang, Y.; Li, M.; Wang, J. Mineralization of plant residues and native soil carbon as affected by soil fertility and residue type. J. Soils Sediments 2019, 19, 1407–1415. [Google Scholar] [CrossRef]
- Steenwerth, K.; Drenovsky, R.; Lambert, J.-J.; Kluepfel, D.; Scow, K.; Smart, D. Soil morphology, depth and grapevine root frequency influence microbial communities in a Pinot noir vineyard. Soil Biol. Biochem. 2008, 40, 1330–1340. [Google Scholar] [CrossRef]
- Rangel-Vasconcelos, L.G.T.; Zarin, D.J.; Oliveira, F.D.A.; Vasconcelos, S.S.; Carvalho, C.J.R.D.; Santos, M.M.D.L.S. Effect of water availability on soil microbial biomass in secondary forest in eastern Amazonia. Rev. Bras. Cienc. Solo 2015, 39, 377–384. [Google Scholar] [CrossRef]
- Sturz, A.; Christie, B. Beneficial microbial allelopathies in the root zone: The management of soil quality and plant disease with rhizobacteria. Soil Tillage Res. 2003, 72, 107–123. [Google Scholar] [CrossRef]
- Kyambadde, J.; Kansiime, F.; Gumaelius, L.; Dalhammar, G. A comparative study of Cyperus papyrus and Miscanthidium violaceum-based constructed wetlands for wastewater treatment in a tropical climate. Water Res. 2004, 38, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.S.; Gupta, V.K. Soil microbial biomass: A key soil driver in management of ecosystem functioning. Sci. Total Environ. 2018, 634, 497–500. [Google Scholar] [CrossRef]
- Chaudhari, P.R.; Ahire, D.V.; Ahire, V.D.; Chkravarty, M.; Maity, S. Soil bulk density as related to soil texture, organic matter content and available total nutrients of Coimbatore soil. Int. J. Sci. Res. Publ. 2013, 3, 1–8. [Google Scholar]
- Wang, H.; Piazza, S.C.; Sharp, L.A.; Stagg, C.L.; Couvillion, B.R.; Steyer, G.D.; McGinnis, T.E. Determining the spatial variability of wetland soil bulk density, organic matter, and the conversion factor between organic matter and organic carbon across coastal Louisiana, USA. J. Coast. Res. 2017, 33, 507–517. [Google Scholar] [CrossRef]
- Hatton, R.; DeLaune, R.; Patrick, W., Jr. Sedimentation, accretion, and subsidence in marshes of Barataria Basin, Louisiana. Limnol. Oceanogr. 1983, 28, 494–502. [Google Scholar] [CrossRef]
- Nyman, J.A.; DeLaune, R.D.; Roberts, H.H.; Patrick, W., Jr. Relationship between vegetation and soil formation in a rapidly submerging coastal marsh. Mar. Ecol.-Prog. Ser. 1993, 96, 269–279. [Google Scholar] [CrossRef]
- Bi, X.; Li, B.; Nan, B.; Fan, Y.; Fu, Q.; Zhang, X. Characteristics of soil organic carbon and total nitrogen under various grassland types along a transect in a mountain-basin system in Xinjiang, China. J. Arid Land 2018, 10, 612–627. [Google Scholar] [CrossRef]
- Bai, J.; Deng, W.; Zhang, Y. Spatial distribution of nitrogen and phosphorus in soil of Momoge Wetland. J. Soil Water Conserv. 2001, 15, 79–81. [Google Scholar]
- Baumann, F.; He, J.S.; Schmidt, K.; Kuehn, P.; Scholten, T. Pedogenesis, permafrost, and soil moisture as controlling factors for soil nitrogen and carbon contents across the Tibetan Plateau. Glob. Chang. Biol. 2009, 15, 3001–3017. [Google Scholar] [CrossRef]
- Sun, J.; Wang, H. Soil nitrogen and carbon determine the trade-off of the above-and below-ground biomass across alpine grasslands, Tibetan Plateau. Ecol. Indic. 2016, 60, 1070–1076. [Google Scholar] [CrossRef]
- Zhao, Q.; Bai, J.; Liu, Q.; Lu, Q.; Gao, Z.; Wang, J. Spatial and seasonal variations of soil carbon and nitrogen content and stock in a tidal salt marsh with Tamarix chinensis, China. Wetlands 2016, 36, 145–152. [Google Scholar] [CrossRef]
- Njeru, C.M.; Ekesi, S.; Mohamed, S.; Kinyamario, J.; Kiboi, S.; Maeda, E. Assessing stock and thresholds detection of soil organic carbon and nitrogen along an altitude gradient in an east Africa mountain ecosystem. Geoderma Reg. 2017, 10, 29–38. [Google Scholar] [CrossRef]
- De Neve, S.; Hofman, G. Influence of soil compaction on carbon and nitrogen mineralization of soil organic matter and crop residues. Biol. Fertil. Soils 2000, 30, 544–549. [Google Scholar] [CrossRef]
- Anh, P.T.Q.; Gomi, T.; MacDonald, L.H.; Mizugaki, S.; Van Khoa, P.; Furuichi, T. Linkages among land use, macronutrient levels, and soil erosion in northern Vietnam: A plot-scale study. Geoderma 2014, 232, 352–362. [Google Scholar] [CrossRef]
- Zhang, C.; Nie, S.; Liang, J.; Zeng, G.; Wu, H.; Hua, S.; Liu, J.; Yuan, Y.; Xiao, H.; Deng, L. Effects of heavy metals and soil physicochemical properties on wetland soil microbial biomass and bacterial community structure. Sci. Total Environ. 2016, 557, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Esperschütz, J.; Zimmermann, C.; Dümig, A.; Welzl, G.; Buegger, F.; Elmer, M.; Munch, J.C.; Schloter, M. Dynamics of microbial communities during decomposition of litter from pioneering plants in initial soil ecosystems. Biogeosciences 2013, 10, 5115–5124. [Google Scholar] [CrossRef]
- Dieckow, J.; Mielniczuk, J.; Knicker, H.; Bayer, C.; Dick, D.P.; Kögel-Knabner, I. Comparison of carbon and nitrogen determination methods for samples of a paleudult subjected to no-till cropping systems. Sci. Agric. 2007, 64, 532–540. [Google Scholar] [CrossRef]
- Hedges, J.I.; Stern, J.H. Carbon and nitrogen determinations of carbonate-containing solids. Limnol. Oceanogr. 1984, 29, 657–663. [Google Scholar] [CrossRef]
- Blake, G.R.; Hartge, K. Bulk density. In Methods of Soil Analysis: Part 1 Physical and Mineralogical Methods, 2nd ed.; Klute, A., Ed.; American Society of Agronomy: Madison, WI, USA, 1986; pp. 363–375. [Google Scholar]
- Brookes, P.; Landman, A.; Pruden, G.; Jenkinson, D. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Tate, K.; Ross, D.; Feltham, C. A direct extraction method to estimate soil microbial C: Effects of experimental variables and some different calibration procedures. Soil Biol. Biochem. 1988, 20, 329–335. [Google Scholar] [CrossRef]
- Jenkinson, D.S.; Ladd, J.N. Microbial biomass in soil: Measurement and turnover. In Soil Biochemistry, 1st ed.; Paul, E.A., Ladd, J.N., Eds.; CRC Press: Boca Raton, FL, USA, 1981; pp. 415–471. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Properties | A | B | C | D | E | F | G | p-Value |
|---|---|---|---|---|---|---|---|---|
| SOC (g kg−1) | 21.62 ± 1.73 cd | 17.25 ± 4.31 d | 27.22 ± 2.37 ab | 32.46 ± 4.81 a | 30.54 ± 3.84 ab | 26.02 ± 3.84 bc | 26.73 ± 2.11 bc | 0.000 *** |
| TN (g kg−1) | 0.62 ± 0.13 bc | 0.48 ± 0.37 b | 1.18 ± 0.24 a | 1.40 ± 0.49 a | 1.18 ± 0.48 a | 0.93 ± 0.38 abc | 1.10 ± 0.14 ab | 0.003 ** |
| C:N ratio | 52.38 ± 4.70 a | 43.76 ± 13.20 ab | 26.76 ± 4.29 d | 26.64 ± 7.46 d | 32.90 ± 14.00 bc | 31.21 ± 10.21 bc | 28.07 ± 4.09 d | 0.001 *** |
| SMBC (mg kg−1) | 995.51 ± 195.82 a | 880.71 ± 96.85 a | 859.44 ± 262.87 a | 1156.67 ± 453.39 a | 1123.66 ± 403.70 a | 954.79 ± 220.41 a | 917.41 ± 121.60 a | 0.536 |
| SMBN (mg kg−1) | 12.44 ± 5.78 a | 6.02 ± 2.93 a | 12.35 ± 11.59 a | 46.22 ± 78.82 a | 11.27 ± 3.76 a | 47.97 ± 22.97 a | 36.14 ± 5.89 ab | 0.171 |
| BD (g cm−3) | 1.50 ± 0.07 ab | 1.58 ± 0.17 a | 1.35 ± 0.12 bc | 1.26 ± 0.17 d | 1.23 ± 0.17 d | 1.31 ± 0.15 bc | 1.21 ± 0.07 d | 0.001 *** |
| MC (%) | 25.05 ± 1.92 cd | 21.77 ± 1.75 d | 38.73 ± 5.73 a | 39.11 ± 5.48 a | 36.22 ± 5.98 ab | 30.06 ± 1.55 bc | 27.20 ± 1.62 cd | 0.000 *** |
| pH | 8.37 ± 0.17 ab | 8.35 ± 0.12 ab | 8.39 ± 0.07 ab | 8.27 ± 0.15 b | 8.25 ± 0.12 b | 8.51 ± 0.21 a | 8.35 ± 0.02 ab | 0.101 |
| SOC | TN | SMBC | SMBN | C:N Ratio | MC | BD | |
|---|---|---|---|---|---|---|---|
| SOC | |||||||
| TN | 0.91 ** | ||||||
| SMBC | 0.35 * | 0.46 ** | |||||
| SMBN | 0.25 | 0.36 * | 0.58 ** | ||||
| C:N Ratio | −0.73 ** | −0.86 ** | −0.31 | −0.37 * | |||
| MC | 0.75 ** | 0.74 ** | 0.44 ** | 0.31 | −0.61 ** | ||
| BD | −0.81 ** | −0.86 ** | −0.46 ** | −0.51 ** | 0.80 ** | −0.60 ** | |
| pH | −0.26 | −0.37 * | −0.49 ** | −0.28 | 0.29 | −0.28 | 0.33 |
| Y | Factors | Equations | R2 | p |
|---|---|---|---|---|
| SOC | Correlated | Y = 18.160 + 11.630X2 − 0.004 X3 + 0.133X5 + 0.161X6 − 7.264X7 | 0.864 | 0.000 *** |
| All | Y = 0.706 + 11.035X2 − 0.002X3 − 0.017X4 + 0.119X5 + 0.161X6 − 8.879X7 + 2.344X8 | 0.863 | 0.000 *** | |
| TN | Correlated | Y = 1.840 + 0.040X1 + 0.000X3 − 0.000X4 − 0.013X5 + 0.001X6− 0.194X7 − 0.168X8 | 0.914 | 0.000 *** |
| All | Y = 1.840 + 0.040X1 + 0.000X3 − 0.000X4 − 0.013X5 + 0.001X6− 0.194X7 − 0.168X8 | 0.914 | 0.000 *** | |
| SMBC | Correlated | Y = 5132.780 − 8.166X1 + 161.133X2 + 3.394X4 + 7.281X6 + 62.747X7 − 573.348X8 | 0.397 | 0.002 ** |
| All | Y = 4428.302 − 18.234X1 + 461.896X2 + 3.268X4 + 10.839X5 + 8.099X6 − 173.218X7 − 466.619X8 | 0.441 | 0.001 ** | |
| SMBN | Correlated | Y = 213.706 − 1.959X1 − 18.385X2 + 0.056X3 − 0.467X5 − 117.401X7 | 0.402 | 0.001 ** |
| All | Y = 142.587 − 2.565X1 − 17.529X2 − 0.054X3 − 0.395X5 + 0.598X6 − 124.134X7 + 9.076X8 | 0.368 | 0.005 ** |
| Sub-Plot Types | TSOC (kg C m−2) | TSN (kg N m−2) | TSMBC (g C m−2) | TSMBN (g N m−2) |
|---|---|---|---|---|
| A | 16.29 ± 5.73 | 0.47 ± 0.46 | 743.70 ± 140.11 | 9.26 ± 1.44 |
| B | 12.63 ± 4.14 | 0.35 ± 0.03 | 705.53 ± 186.67 | 4.59 ± 0.71 |
| C | 17.17 ± 1.09 | 0.72 ± 0.12 | 576.20 ± 241.39 | 7.82 ± 1.47 |
| D | 20.17 ± 4.90 | 0.83 ± 0.06 | 708.71 ± 268.74 | 25.72 ± 14.36 |
| E | 18.29 ± 3.43 | 0.69 ± 0.17 | 680.22 ± 215.21 | 6.71 ± 1.63 |
| F | 16.97 ± 4.30 | 0.60 ± 0.16 | 617.62 ± 185.11 | 30.52 ± 8.60 |
| G | 16.10 ± 3.26 | 0.67 ± 0.29 | 650.31 ± 169.52 | 22.43 ± 8.91 |
| Sub-Plots | Dominant Vegetation Species | Hydrology | Biomass (kg m−2) |
|---|---|---|---|
| A | Poa annua L., Imperata cylindrica, Polygonum | Dry | 0.38 ± 0.21 |
| B | Phragmites australis, Mongolian wormwood | Inundated | 0.71 ± 0.03 |
| C | Tamarix chinensis Lour., Polygonum amphibium L. | Inundated | 3.74 ± 1.73 |
| D | Scirpus fluviatilis, Polygonum amphibium L., Carex tristachya | Inundated | 0.55 ± 0.19 |
| E | Phragmites australis | Inundated | 0.91 ± 0.08 |
| F | Leymus chinensis | Dry | 0.64 ± 0.17 |
| G | Poa annua L. | Dry | 0.58 ± 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Bao, H.; Kavana, D.J.; Li, Y.; Li, X.; Yan, L.; Xu, W.; Yu, B. Effects of Vegetation Types and Soil Properties on Regional Soil Carbon and Nitrogen in Salinized Reservoir Wetland, Northeast China. Plants 2023, 12, 3767. https://doi.org/10.3390/plants12213767
Wang Y, Bao H, Kavana DJ, Li Y, Li X, Yan L, Xu W, Yu B. Effects of Vegetation Types and Soil Properties on Regional Soil Carbon and Nitrogen in Salinized Reservoir Wetland, Northeast China. Plants. 2023; 12(21):3767. https://doi.org/10.3390/plants12213767
Chicago/Turabian StyleWang, Yuchen, Heng Bao, David J. Kavana, Yuncong Li, Xiaoyu Li, Linlu Yan, Wenjing Xu, and Bing Yu. 2023. "Effects of Vegetation Types and Soil Properties on Regional Soil Carbon and Nitrogen in Salinized Reservoir Wetland, Northeast China" Plants 12, no. 21: 3767. https://doi.org/10.3390/plants12213767
APA StyleWang, Y., Bao, H., Kavana, D. J., Li, Y., Li, X., Yan, L., Xu, W., & Yu, B. (2023). Effects of Vegetation Types and Soil Properties on Regional Soil Carbon and Nitrogen in Salinized Reservoir Wetland, Northeast China. Plants, 12(21), 3767. https://doi.org/10.3390/plants12213767