

Physiological and Biochemical Responses of Commercial Strawberry Cultivars under Optimal and Drought Stress Conditions
 and
and 
Abstract
:1. Introduction
2. Results
2.1. Photosynthetic Pigments, Electrolyte Leakage (EL), and Relative Water Content (RWC)
2.2. Antioxidant Enzymes Activity
2.3. Oxidative Markers and Osmolytes
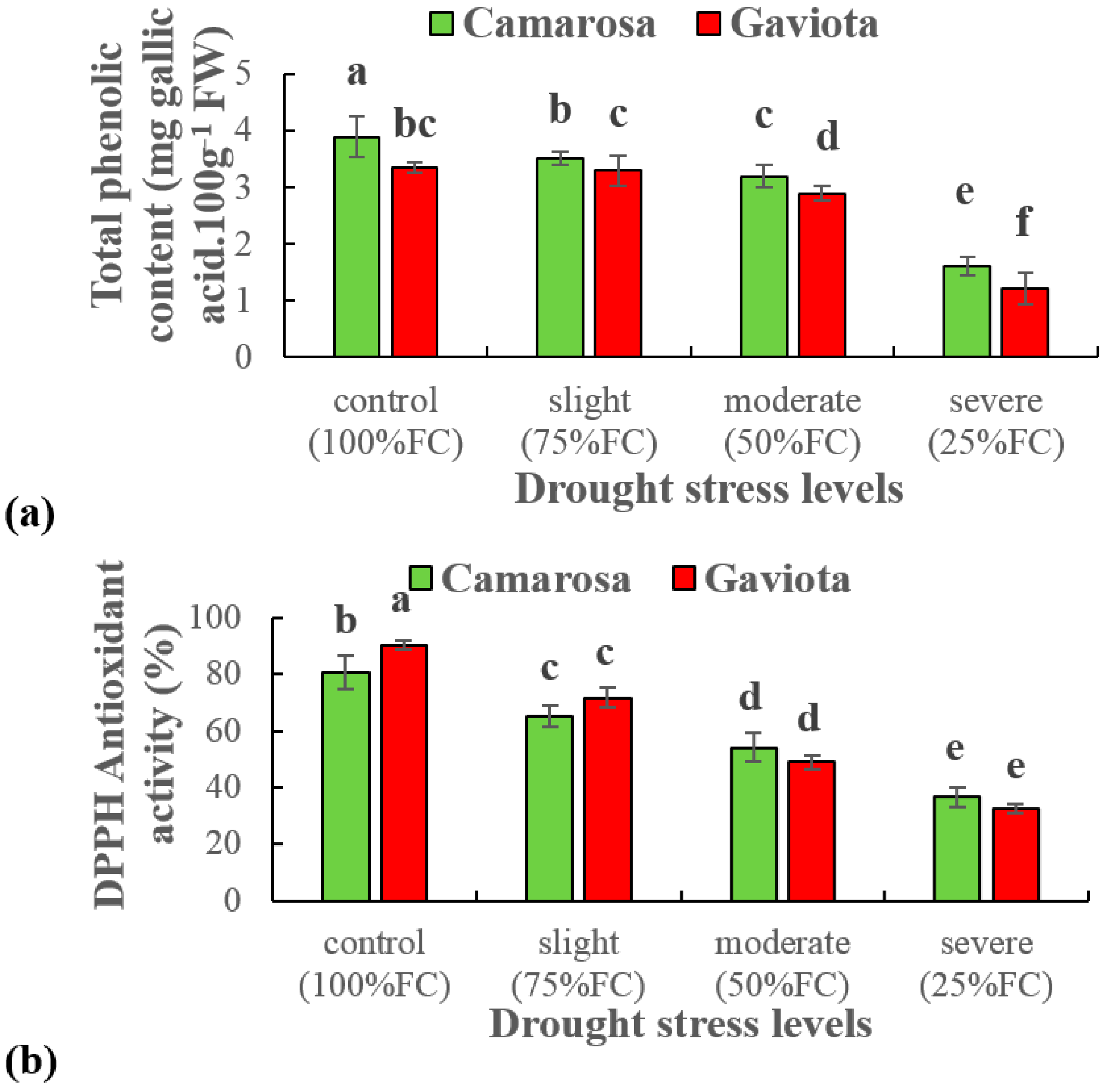
2.4. Phenolic Content and Antioxidant Activity (DPPH Assay)
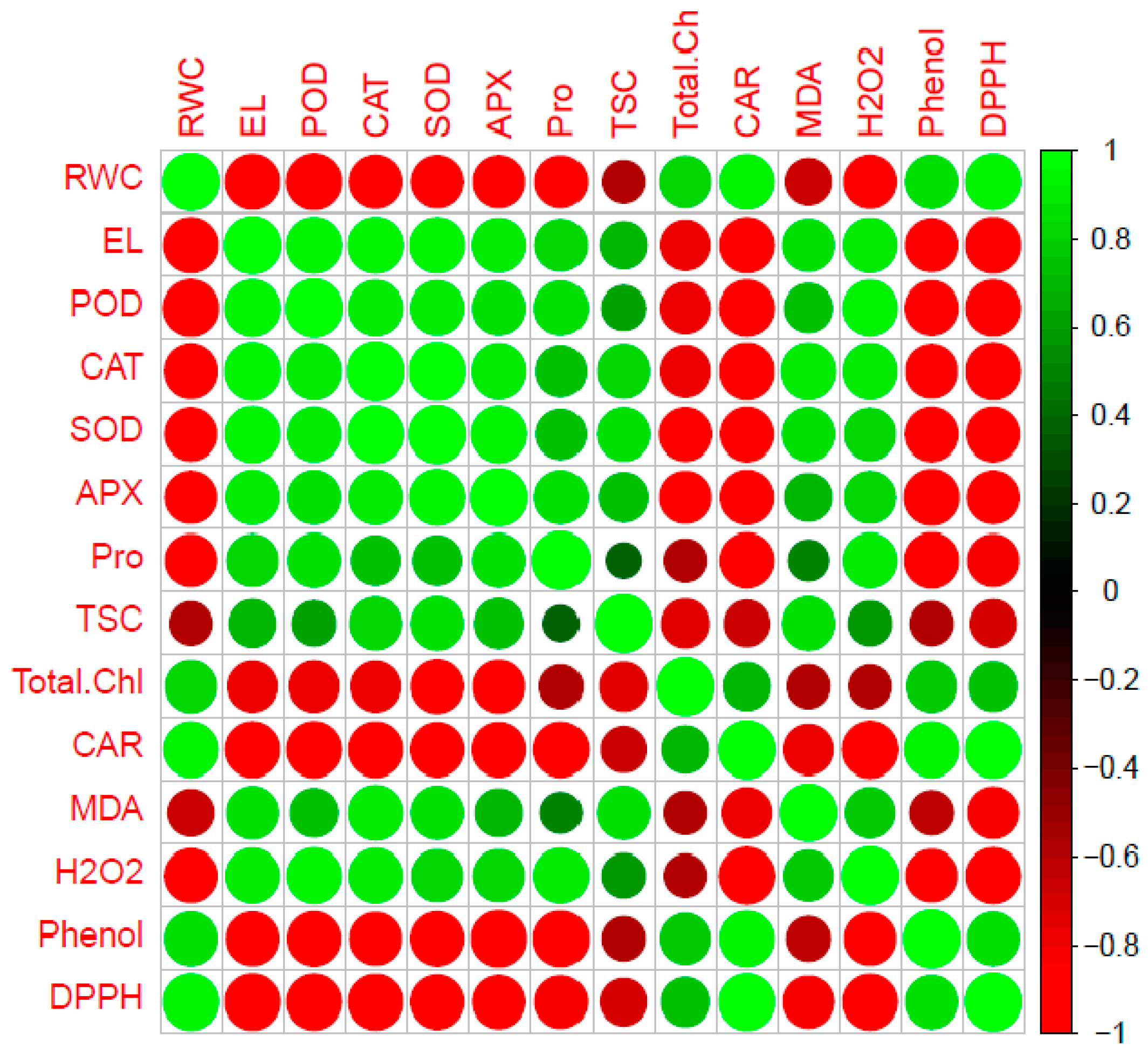
2.5. Heat Map Analysis
3. Discussion
4. Material and Methods
4.1. Plant Material and Treatment
4.2. Photosynthetic Pigments, EL, and RWC
4.3. Antioxidant Enzymes Activity
4.4. Oxidative Markers
4.5. Osmolytes (Proline and Soluble Carbohydrates)
4.6. Phenolic Content and Antioxidant Activity
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Afrin, S.; Gasparrini, M.; Forbes-Hernandez, T.Y.; Reboredo-Rodriguez, P.; Mezzetti, B.; Varela-López, A.; Giampieri, F.; Battino, M. Promising health benefits of the strawberry: A focus on clinical studies. J. Agric. Food Chem. 2016, 64, 4435–4449. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations Statistics. 2021. Available online: https://faostat3.fao.org (accessed on 1 February 2021).
- Gholami, R.; Fahadi Hoveizeh, N.; Zahedi, S.M.; Gholami, H.; Carillo, O. Effect of three water-regimes on morpho-physiological, biochemical and yield responses of local and foreign olive cultivars under field conditions. BMC Plant Biol. 2022, 22, 477. [Google Scholar] [CrossRef] [PubMed]
- Mozafari, A.; Havas, F.; Ghaderi, N. Application of iron nanoparticles and salicylic acid in in vitro culture of strawberries (Fragaria× ananassa Duch.) to cope with drought stress. Plant Cell Tissue Organ Cult. 2018, 132, 511–523. [Google Scholar] [CrossRef]
- Adak, N.; Gubbuk, H.; Tetik, N. Yield, quality and biochemical properties of various strawberry cultivars under water stress. J. Sci. Food Agric. 2018, 98, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, M.S.; Samsampour, D.; Zahedi, S.M.; Zamanian, K.; Rahman, M.M.; Mostofa, M.G.; Tran, L.S.P. Melatonin alleviates drought impact on growth and essential oil yield of lemon verbena by enhancing antioxidant responses, mineral balance, and abscisic acid content. Physiol. Plant. 2021, 172, 1363–1375. [Google Scholar] [CrossRef] [PubMed]
- Elkeilsh, A.; Awad, Y.M.; Soliman, M.H.; Abu-Elsaoud, A.; Abdelhamid, M.T.; El-Metwally, I.M. Exogenous application of β-sitosterol mediated growth and yield improvement in water stressed wheat (Triticum aestivum) involves up-regulated antioxidant system. J. Plant Res. 2019, 132, 881–901. [Google Scholar] [CrossRef]
- Li, J.; Liu, J.; Zhu, T.; Zhao, C.; Li, L.; Chen, M. The Role of melatonin in salt stress responses. Int. J. Mol. Sci. 2019, 20, 1735. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Qiu, L.; Guo, H.; Wang, Y.; Yuan, H.; Yan, D.; Zheng, B. Spermidine induces physiological and biochemical changes in southern highbush blueberry under drought stress. Braz. J. Bot. 2017, 40, 841–851. [Google Scholar] [CrossRef]
- Faaek, M.F.F.; Pirlak, L. Morphological and physiological effects of drought stress on some strawberry cultivars. Selcuk J. Agric. Food Sci. 2021, 35, 194–201. [Google Scholar]
- Gulen, H.; Kesici, M.; Cetinkaya, C.; Ergin, S. Proline and antioxidant enzyme activities in some strawberry cultivars under drought and recovery. Not. Bot. Horti Agrobot. Cluj-Napoca 2018, 46, 570–578. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.H.; Nulit, R.; Sakimin, S.Z. Influence of drought tress on growth biochemical changes and leaf gas exchange of strawberry (Fragaria ananassa Duch.) in Indonesia. AIMS Agric. Food 2022, 7, 37–60. [Google Scholar]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Zhang, S.; Zhang, K.; Li, Y.; Xu, Q.; Liao, C.; et al. Interactive effects of drought and heat stresses on morpho-physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9, 3890. [Google Scholar] [CrossRef] [PubMed]
- Ben Ahmed, C.; Ben Rouina, B.; Sensoy, S.; Boukhris, M.; Ben Abdallah, F. Changes in gas exchange, proline accumulation and antioxidative enzyme activities in three olive cultivars under contrasting water availability regimes. Environ. Exp. Bot. 2009, 67, 345–352. [Google Scholar] [CrossRef]
- Ahmed, A.H.H.; Darwish, E.; Hamoda, S.A.F.; Alobaidy, M.G. Effect of putrescine and humic acid on growth, yield and chemical composition of cotton plants grown under saline soil conditions. Am.-Eurasian J. Agric. Environ. Sci. 2013, 13, 479–497. [Google Scholar]
- Mohammadi, M.; Ghassemi-Golezani, K.; Zehtab-Salmasi, S.; Nasrollahzade, S. Assessment of some physiological traits in spring safflower (Carthamus tinctorius L.) cultivars under water stress. Int. J. Life Sci. 2016, 10, 58–64. [Google Scholar] [CrossRef] [Green Version]
- Rehman, S.; Bilal, M.A.; Rana, R.M.; Tahir, N.; Nawaz Shah, M.K.; Ayalew, H.; Yan, G. Cell membrane stability and chlorophyll content variation in wheat genotypes under conditions of heat and drought. Crop Pasture Sci. 2016, 67, 712–718. [Google Scholar] [CrossRef]
- Kapur, B.; Çeliktopuz, E.; Sarıdaş, M.A.; Kargı, S.P. Irrigation regimes and bio-stimulant application effects on yield and morpho-physiological responses of strawberry. Hortic. Sci. Technol. 2018, 36, 313–325. [Google Scholar]
- Ariza, M.T.; Miranda, L.; Gómez-Mora, J.A.; Medina, J.J.; Lozano, D.; Gavilán, P.; Soria, C.; Martínez-Ferri, E. Yield and fruit quality of strawberry cultivars under different irrigation regimes. Agronomy 2021, 11, 261. [Google Scholar] [CrossRef]
- Kirschbaum, D.S.; Correa, M.; Borquez, A.M.; Larson, K.D.; DeJong, T.M. Water requirements and water use efficiency of fresh and waiting bed strawberry plants. Acta Hortic. 2004, 664, 347–352. [Google Scholar] [CrossRef]
- Adiba, A.; Hssaini, L.; Haddioui, A.; Hamdani, A.; Charafi, J.; Iraqui, S.E.; Razouk, R. Pomegranate plasticity to water stress: Attempt to understand interactions between cultivar, year and stress level. Heliyon 2021, 7, e07403. [Google Scholar] [CrossRef]
- Li, C.; Tan, D.X.; Liang, D.; Chang, C.; Jia, D.; Ma, F. Melatonin mediates the regulation of ABA metabolism, free-radical scavenging, and stomatal behavior in two Malus species under drought stress. J. Exp. Bot. 2015, 66, 669–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klamkowski, K.; Treder, W. Response to drought stress of three strawberry cultivars grown under greenhouse conditions. J. Fruit Ornam. Plant Res. 2008, 16, 179–188. [Google Scholar]
- Grant, O.M.; Johnson, A.W.; Davies, M.J.; James, C.M.; Simpson, D.W. Physiological and morphological diversity of cultivated strawberry (Fragaria × ananassa) in response to water deficit. Environ. Exp. Bot. 2010, 68, 264–272. [Google Scholar] [CrossRef]
- Ghaderi, N.; Siosemardeh, A. Response to drought stress of two strawberry cultivars (cv. Kurdistan and Selva). Hortic. Environ. Biotechnol. 2011, 52, 6–12. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, J.; Burgess, P.; Rossi, S.; Huang, B. Interactive effects of melatonin and cytokinin on alleviating drought-induced leaf senescence in creeping bentgrass (Agrostis stolonifera). Environ. Exp. Bot. 2018, 145, 1–11. [Google Scholar] [CrossRef]
- Basu, S.; Ramegowda, V.; Kumar, A.; Pereira, A. Plant adaptation to drought stress. F1000Research 2016, 5, 1554. [Google Scholar] [CrossRef]
- Kostopoulou, Z.; Therios, I.; Roumeliotis, E.; Kanellis, A.K.; Molassiotis, A. Melatonin combined with ascorbic acid provides salt adaptation in Citrus aurantium L. seedlings. Plant Physiol. Biochem. 2015, 86, 155–165. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Akram, N.A.; Ashraf, M. Osmoprotection in plants under abiotic stresses: New insights into a classical phenomenon. Planta 2019, 251, 3. [Google Scholar] [CrossRef] [Green Version]
- Ahmadipour, S.; Arji, I.; Ebadi, A.; Abdossi, V. Physiological and biochemical responses of some olive cultivars (Olea europaea L.) to water stress. Cell. Mol. Biol. 2018, 64, 20–29. [Google Scholar] [CrossRef]
- Naeem, M.; Naeem, M.S.; Ahmad, R.; Ahmad, R.; Ashraf, M.Y.; Ihsan, M.Z.; Nawaz, F.; Athar, H.U.R.R.; Ashraf, M.; Abbas, H.T.; et al. Improving drought tolerance in maize by foliar application of boron: Water status, antioxidative defense and photosynthetic capacity. Arch. Agron. Soil Sci. 2018, 64, 626–639. [Google Scholar] [CrossRef]
- Jiang, M.; Zhang, J. Water stress-induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulates the activities of antioxidant enzymes in maize leaves. J. Exp. Bot. 2002, 53, 2401–2410. [Google Scholar] [CrossRef] [Green Version]
- Jamshidi Goharrizi, K.; Amirmahani, F.; Salehi, F. Assessment of changes in physiological and biochemical traits in four pistachio rootstocks under drought, salinity and drought + salinity stresses. Physiol. Plant. 2020, 168, 973–989. [Google Scholar] [CrossRef]
- Dawood, M.G.; El-Awadi, M.E. Alleviation of salinity stress on Vicia faba L. plants via seed priming with melatonin. Acta Biol. Colomb. 2015, 20, 223–235. [Google Scholar] [CrossRef]
- Shinde, S.; Villamor, J.G.; Lin, W.; Sharma, S.; Verslues, P.E. Proline coordination with fatty acid synthesis and redox metabolism of chloroplast and mitochondria. Plant Physiol. 2016, 172, 1074–1088. [Google Scholar] [CrossRef] [Green Version]
- Szabados, L.; Savoure, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Hussein, E.A.; El-Kerdany, A.Y.; Afifi, M.K. Effect of drought and salinity stresses on two strawberry cultivars during their regeneration in vitro. Int. J. Innov. Sci. Eng. Technol. 2017, 4, 83–93. [Google Scholar]
- Gao, H.; Lu, Z.; Yang, Y.; Wang, D.; Yang, Y.; Cao, M.; Cao, W. Melatonin treatment reduces chilling injury in peach fruit through its regulation of membrane fatty acid contents and phenolic metabolism. Food Chem. 2018, 245, 659–666. [Google Scholar] [CrossRef]
- Peres, F.; Martins, L.L.; Mourato, M.; Vitorino, C.; Antunes, P.; Ferreira-Dias, S. Phenolic compounds of “Galega Vulgar” and “Cobrançosa” olive oils along early ripening stages. Food Chem. 2016, 211, 51–58. [Google Scholar] [CrossRef]
- Kargi, S.P.; Kafkas, E.; Baktemur, G. Yield and quality characteristics of some strawberry cultivars in Adana, Turkey. Acta Hortic. 2012, 926, 163–167. [Google Scholar] [CrossRef]
- Souza, C.C.; Oliveira, F.A.; Silva, I.F.; AmorimNeto, M.S. Evaluation of methods of available water determination and irrigation management in “terra roxa” under cotton crop. Rev. Bras. Eng. Agric. Ambient. 2000, 4, 338–342. [Google Scholar] [CrossRef] [Green Version]
- Arnon, D.I. Copper enzymes in isolated chloroplasts polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. NaCl-induced senescence in leaves of rice (Oryza sativa L.) cultivars differing in salinity resistance. Ann. Bot. 1996, 78, 389–398. [Google Scholar] [CrossRef]
- Gucci, R.; Lombardini, L.; Tattini, M. Analysis of leaf water relations in leaves of two olive (Olea europaea) cultivars differing in tolerance to salinity. Tree Physiol. 1997, 17, 13–21. [Google Scholar] [CrossRef]
- Hemeda, H.M.; Klein, B. Effects of naturally occurring antioxidants on peroxidase activity of vegetable extracts. J. Food Sci. 1990, 55, 184–185. [Google Scholar] [CrossRef]
- Hossain, M.A.; Hasanuzzaman, M.; Fujita, M. Up-regulation of antioxidant and glyoxalase systems by exogenous glycinebetaine and proline in mung bean confer tolerance to cadmium stress. Physiol. Mol. Biol. Plants 2010, 26, 259–272. [Google Scholar] [CrossRef] [Green Version]
- Mostofa, M.G.; Hossain, M.A.; Fujita, M. Trehalose pretreatment induces salt tolerance in rice (Oryza sativa L.) seedlings: Oxidative damage and co-induction of antioxidant defense and glyoxalase systems. Protoplasma 2015, 252, 461–475. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen-peroxide is scavenged by ascorbate-specific peroxidase in spinach-chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plant extracts by anthrone. Biochemistry 1954, 57, 508–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Brand-Williams, W.B.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Lebensm.-Wiss. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Cultivar | Drought Stress Level | Total Chl | CAR | EL (%) | RWC (%) |
|---|---|---|---|---|---|
| (mg g−1 FW) | |||||
| Camarosa | Control 1 | 3.69 ± 0.08 a | 1.34 ± 0.16 ab | 17.32 ± 0.93 f | 87.31 ± 1.35 a |
| Slight 2 | 3.21 ± 0.07 b | 1.24 ± 0.24 bc | 21.52 ± 0.61 e | 78.38 ± 2.23 b | |
| Moderate 3 | 2.35 ± 0.07 e | 1.13 ± 0.26 cd | 33.52 ± 0.66 c | 66.52 ± 1.04 d | |
| Severe 4 | 1.07 ± 0.11 f | 0.83 ± 0.21 e | 39.26 ± 0.71 b | 59.24 ± 3.21 ef | |
| Gaviota | Control 1 | 2.85 ± 0.09 c | 1.44 ± 0.13 a | 14.60 ± 1.19 g | 83.55 ± 1.75 a |
| Slight 2 | 2.61 ± 0.11 d | 1.35 ± 0.25 ab | 22.60 ± 1.06 e | 74.26 ± 2.72 c | |
| Moderate 3 | 2.49 ± 0.10 de | 1.02 ± 0.08 d | 28.57 ± 0.70 d | 61.66 ± 3.68 e | |
| Severe 4 | 2.37 ± 0.10 e | 0.68 ± 0.18 e | 41.26 ± 0.85 a | 55.65 ± 3.64 f | |
| df | Mean Square | ||||
| Cultivar | 1 | 0.0024 ns | 0.0008 ns | 7.878 ns | 82.770 ** |
| Drought | 3 | 2.831 ** | 0.486 ** | 675.909 ** | 975.481 ** |
| Cultivar × Drought | 3 | 1.294 ** | 0.028 * | 15.869 ** | 1.993 * |
| Cultivar | Drought Stress Level | POD | CAT | SOD | APX |
|---|---|---|---|---|---|
| (μmol min−1 mg−1 Protein) | (μmol min−1 mg−1 Protein) | (U min−1 mg−1 Protein) | (μmol min−1 mg−1 Protein) | ||
| Camarosa | Control 1 | 4.04 ± 0.15 g | 0.05 ± 0.01 de | 14.79 ± 0.93 e | 1.39 ± 0.10 e |
| Slight 2 | 4.93 ± 0.30 ef | 0.07 ± 0.06 d | 15.56 ± 0.89 de | 1.53 ± 0.25 de | |
| Moderate 3 | 7.79 ± 0.19 d | 0.11 ± 0.01 b | 17.81 ± 0.84 c | 1.74 ± 0.11 cd | |
| Severe 4 | 9.59 ± 0.21 b | 0.16 ± 0.01 a | 21.76 ± 1.18 a | 3.12 ± 0.09 a | |
| Gaviota | Control 1 | 4.65 ± 0.27 f | 0.03 ± 0.01 e | 14.36 ± 0.10 e | 1.45 ± 0.18 e |
| Slight 2 | 5.16 ± 0.19 e | 0.04 ± 0.01 e | 15.34 ± 1.11 d e | 1.72 ± 0.23 cd | |
| Moderate 3 | 8.48 ± 0.43 c | 0.09 ± 0.01 c | 16.35 ± 0.95 d | 1.87 ± 0.12 c | |
| Severe 4 | 10.28 ± 0.37 a | 0.12 ± 0.03 b | 19.83 ± 0.62 b | 2.80 ± 0.14 b | |
| df | Mean Square | ||||
| Cultivar | 1 | 1.859 ** | 0.003 ** | 6.101 * | 0.008 ns |
| Drought | 3 | 41.372 ** | 0.014 ** | 45.414 ** | 0.032 ** |
| Cultivar × Drought | 3 | 0.072 * | 0.00003 * | 1.004 * | 0.054 * |
| Cultivar | Drought Stress Level | H2O2 | MDA | Proline | Total Soluble Carbohydrates |
|---|---|---|---|---|---|
| (nmol g−1 FW) | (µmol g−1 FW) | (mg g−1 FW) | |||
| Camarosa | Control 1 | 4.85 ± 0.19 d | 4.23 ± 0.16 c | 1.18 ± 0.11 d | 16.12 ± 0.15 d |
| Slight 2 | 5.02 ± 0.29 d | 4.37 ± 0.11 c | 1.26 ± 0.14 cd | 16.60 ± 0.11 c | |
| Moderate 3 | 5.54 ± 0.17 c | 5.04 ± 0.08 b | 1.35 ± 0.08 cd | 17.18 ± 0.26 b | |
| Severe 4 | 6.58 ± 0.11 b | 5.43 ± 0.12 a | 2.02 ± 0.17 b | 23.18 ± 0.17 a | |
| Gaviota | Control 1 | 3.89 ± 0.12 f | 3.09 ± 0.13 e | 1.24 ± 0.13 cd | 13.50 ± 0.33 f |
| Slight 2 | 4.28 ± 0.24 e | 3.53 ± 0.10 d | 1.42 ± 0.17 c | 13.94 ± 0.27 e | |
| Moderate 3 | 6.37 ± 0.10 b | 4.14 ± 0.19 c | 1.85 ± 0.18 b | 15.85 ± 0.21 d | |
| Severe 4 | 7.56 ± 0.11 a | 4.93 ± 0.15 b | 2.82 ± 0.06 a | 16.72 ± 0.20 c | |
| df | Mean Square | ||||
| Cultivar | 1 | 0.007 ns | 4.293 ** | 0.870 ** | 63.342 ** |
| Drought | 3 | 9.419 ** | 2.748 ** | 1.774 ** | 98.916 ** |
| Cultivar × Drought | 3 | 1.589 ** | 0.104 ** | 0.173 ** | 21.279 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zahedi, S.M.; Hosseini, M.S.; Fahadi Hoveizeh, N.; Kadkhodaei, S.; Vaculík, M. Physiological and Biochemical Responses of Commercial Strawberry Cultivars under Optimal and Drought Stress Conditions. Plants 2023, 12, 496. https://doi.org/10.3390/plants12030496
Zahedi SM, Hosseini MS, Fahadi Hoveizeh N, Kadkhodaei S, Vaculík M. Physiological and Biochemical Responses of Commercial Strawberry Cultivars under Optimal and Drought Stress Conditions. Plants. 2023; 12(3):496. https://doi.org/10.3390/plants12030496
Chicago/Turabian StyleZahedi, Seyed Morteza, Marjan Sadat Hosseini, Narjes Fahadi Hoveizeh, Saeid Kadkhodaei, and Marek Vaculík. 2023. "Physiological and Biochemical Responses of Commercial Strawberry Cultivars under Optimal and Drought Stress Conditions" Plants 12, no. 3: 496. https://doi.org/10.3390/plants12030496
APA StyleZahedi, S. M., Hosseini, M. S., Fahadi Hoveizeh, N., Kadkhodaei, S., & Vaculík, M. (2023). Physiological and Biochemical Responses of Commercial Strawberry Cultivars under Optimal and Drought Stress Conditions. Plants, 12(3), 496. https://doi.org/10.3390/plants12030496