

Multispecies Bacterial Bio-Input: Tracking and Plant-Growth-Promoting Effect on Lettuce var. sagess

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Tracking of Bacterial Strains
2.1.1. Effectiveness of Plate Colony-Counting Technique
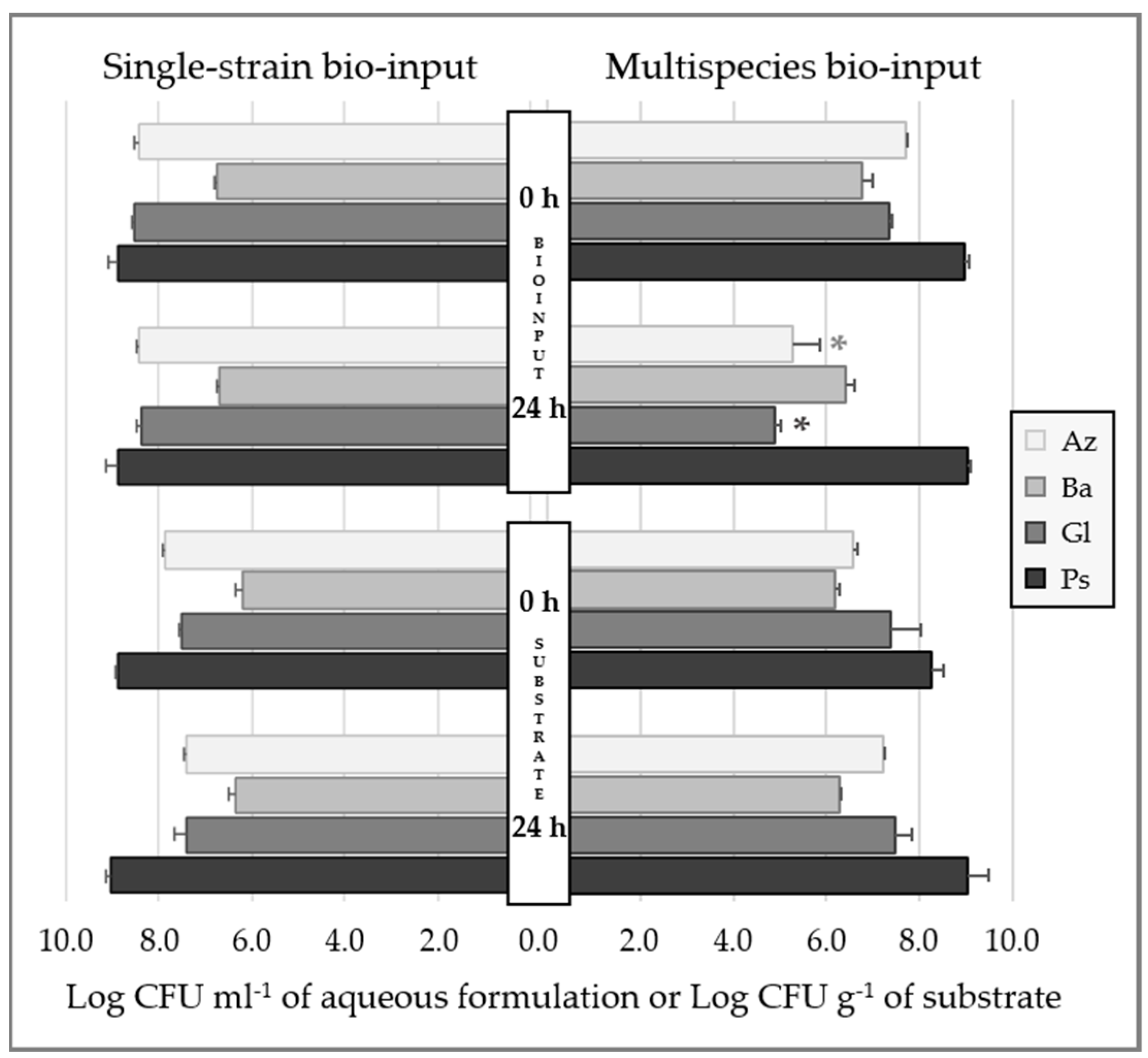
2.1.2. Bacterial Survival in Aqueous Formulation
2.1.3. Bacterial Survival in Inoculated Substrate
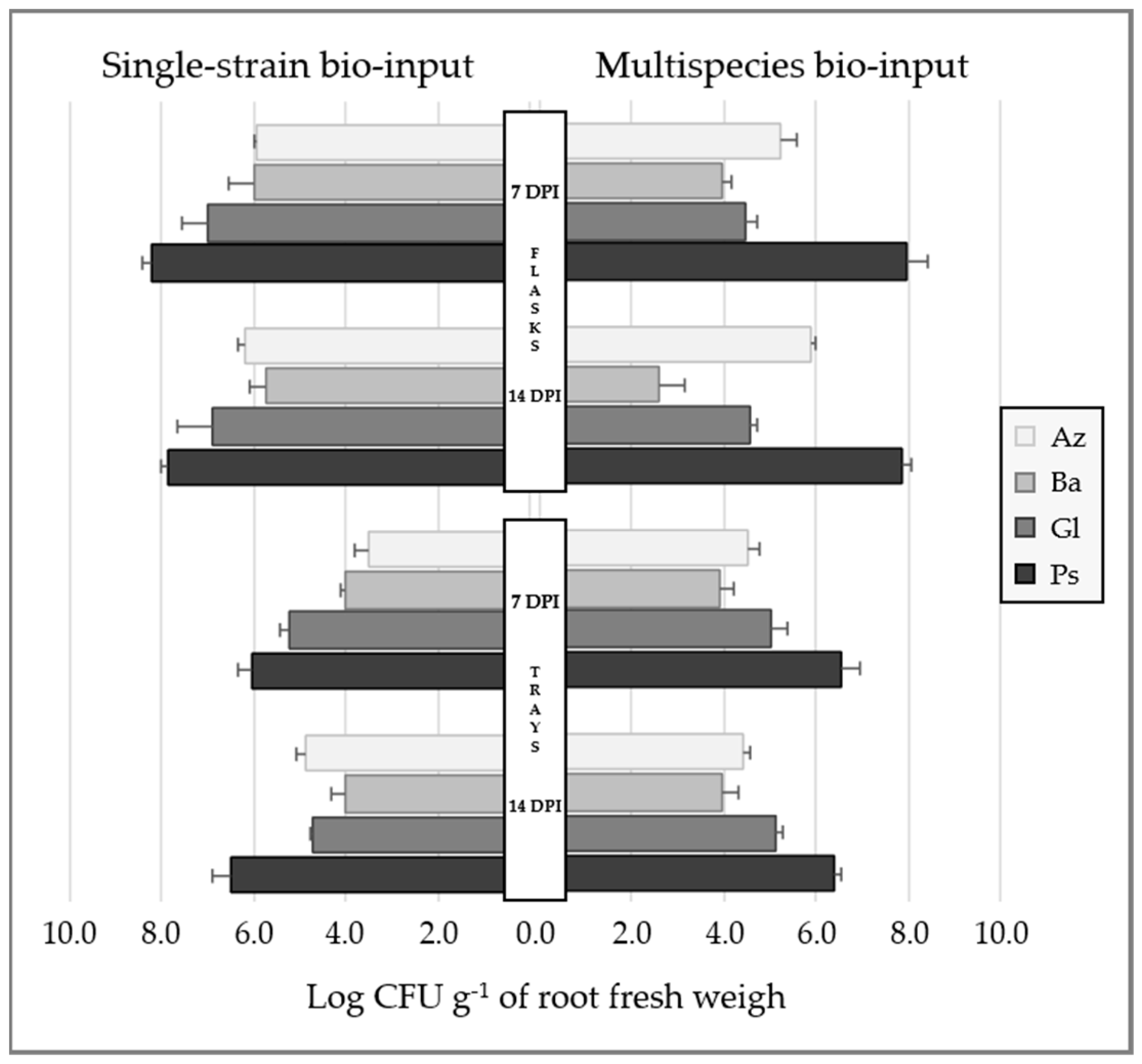
2.1.4. Bacterial Root Colonization in Inoculated Lettuce Seedlings
2.2. Effect of the Experimental Bio-Inputs on Plant Growth of Lettuce
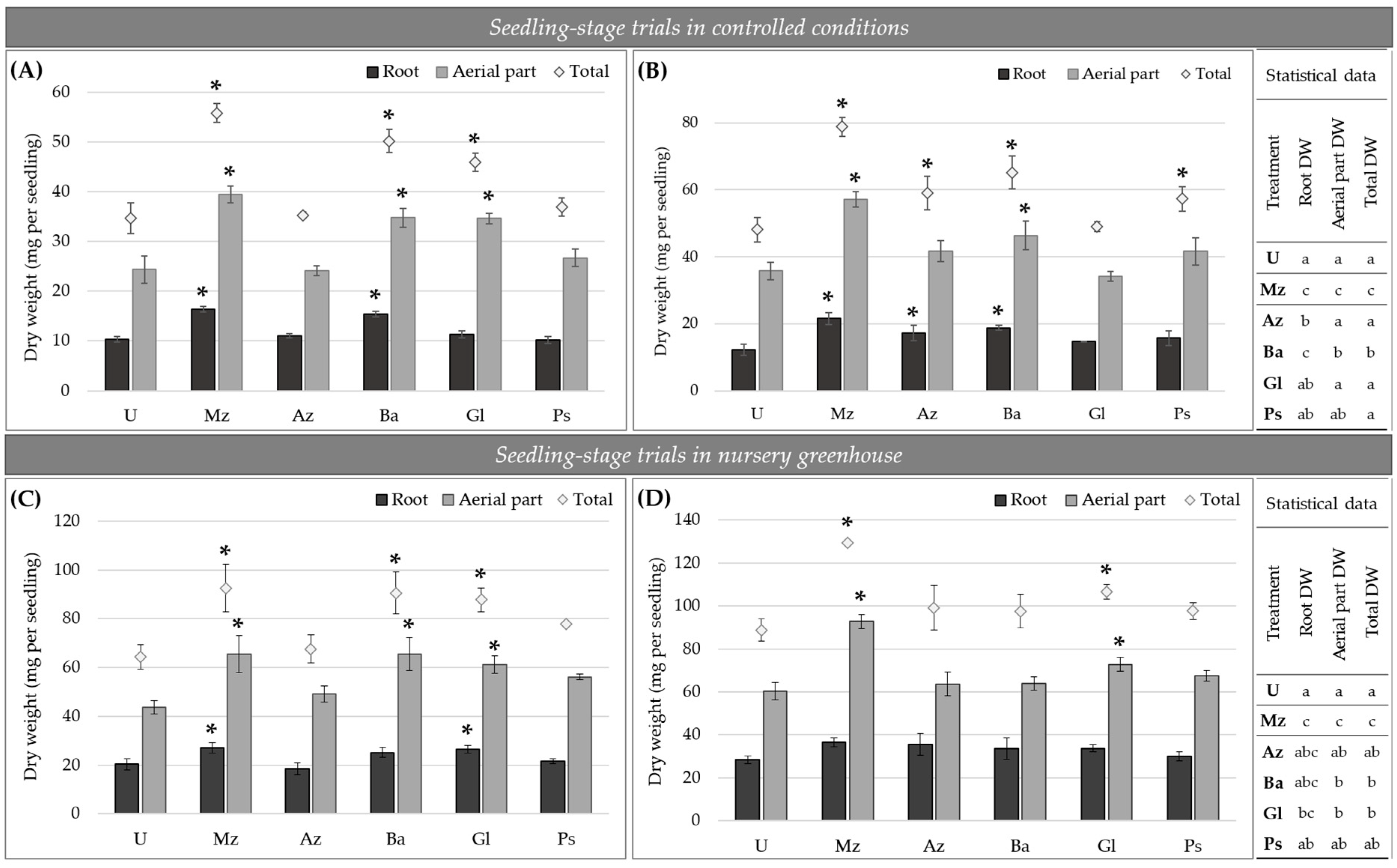
2.2.1. Plant-Growth-Promoting Effect at Seedling Stage
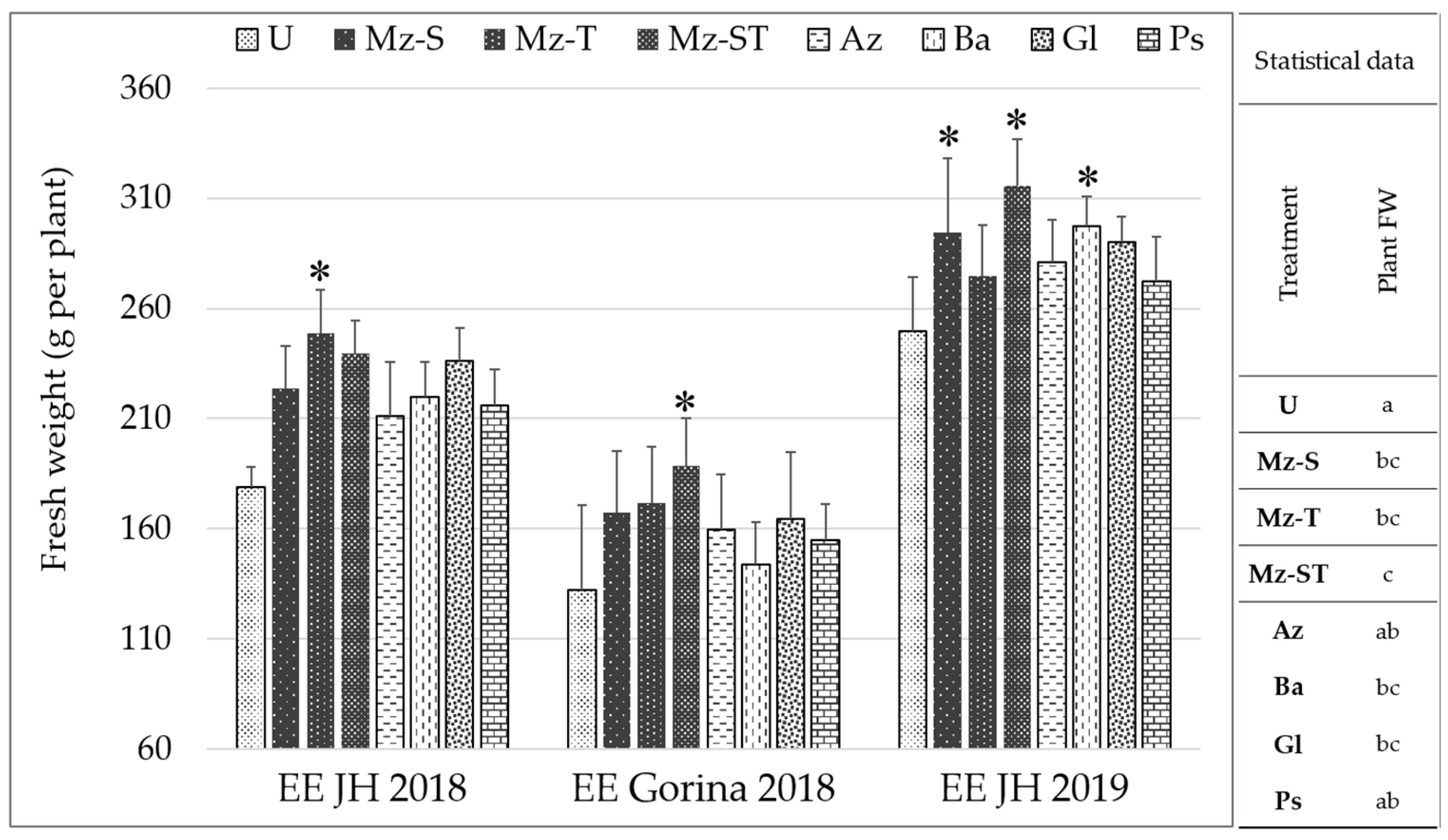
2.2.2. Plant-Growth-Promoting Effect at Productive Stage
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Maintenance
4.2. Bacterial Cultures and Formulation of Experimental Bio-Inputs
4.3. Bacterial Survival Assay
4.3.1. Bacterial Survival in Aqueous Formulations
4.3.2. Bacterial Survival in Inoculated Substrate
4.4. Plant Growth Conditions and Inoculation
4.4.1. Lettuce Plants at Seedling Stage
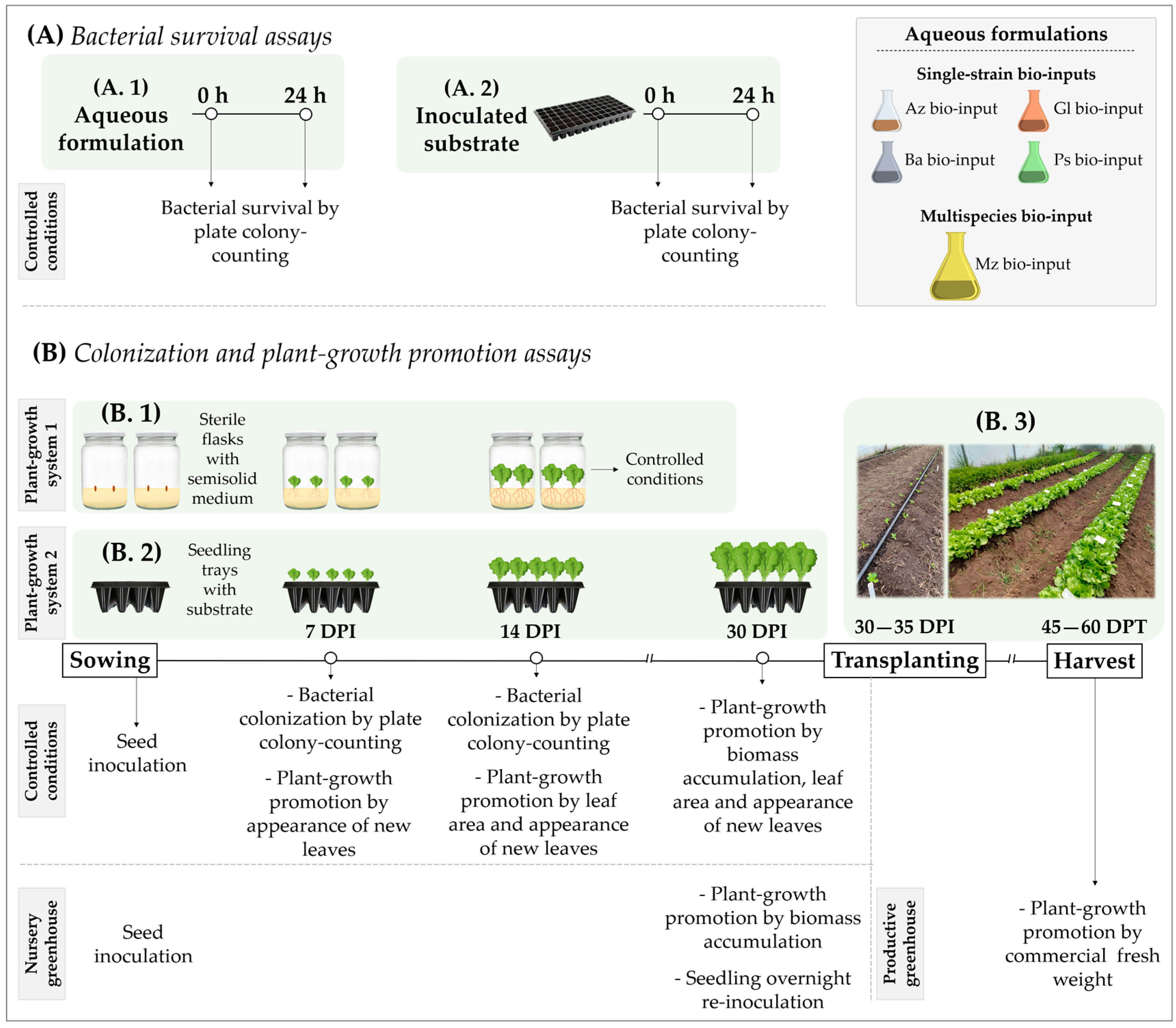
- Plant-growth system 1: flasks with sterile semisolid agar-containing medium and seeds, disinfected to diminish the presence of indigenous microorganisms, were used in controlled conditions (plant-growth chamber at 26 °C with photoperiod light/dark cycle of 16 and 8 h, respectively), a suitable environment to study plant-bacteria interactions. Lettuce seeds were surface-disinfected with 20% v/v sodium hypochlorite (46 g L−1 NaClO) for 10 min and washed three times with PBS [21,55]. Disinfected seeds were placed between wet sterile filter papers in the dark and incubated for ~48 h at 28 °C for germination. Sprouted seeds were immersed in each experimental bio-input (Az, Ba, Gl, Ps and Mz bio-input) for 20 min without agitation and placed on flasks containing 100 mL of sterile semisolid Fåhraeus agar-containing medium (0.5% w/v) at pH 6.00 [56]. Distilled sterile water was used instead of experimental bio-inputs as the uninoculated control (U treatment). Seedlings were grown in controlled conditions throughout the assay. Two independent assays were performed using a completely randomized design with six treatments and three biological replicates each: uninoculated, single-inoculated with Az, Ba, Gl or Ps bio-inputs, and co-inoculated with Mz bio-input (U, Az, Ba, Gl, Ps and Mz treatments, respectively). Samples were taken on different days to evaluate root colonization as described below (Figure 1(B.1)).
- Plant-growth system 2: trays with sterile/nonsterile substrate and disinfected/nondisinfected seeds were used to study plant–bacteria interactions in controlled conditions (growth chamber) and in nursery greenhouse conditions (a commercial plant nursery) (Figure 1(B.2)).
- -
- Lettuce seedlings grown in controlled conditions: surface-disinfected seeds were placed in seedling trays filled with an autoclaved 1:1 mixture of organic compost and amended soil (pH 6.4 and 6.5, respectively; Rincón Verde, Biofertyl–SRL) as substrate. Experimental Az, Ba, Gl, Ps and Mz bio-inputs were added on seeds (immediately after formulating the bio-inputs) at a ratio of 1 mL per cell (10 mL of substrate), after which seeds were covered with a thin layer of substrate. Sterile distilled water was used instead of experimental bio-inputs as the uninoculated control (U treatment). Seedling trays were grown in controlled conditions throughout the test. Plants were kept with irrigation on demand and modified Hoagland’s solution with KNO3 as the nitrogen source (0.14 g L−1 KH2PO4; 0.50 g L−1 KNO3; 1.18 g L−1 Ca2(NO3)2·4H2O; 0.49 g L−1 MgSO4·7H2O; 2.86 mg L−1 H3BO3; 1.81 mg L−1 MnCl·4H2O; 0.22 mg L−1 ZnSO4·7H2O; 0.08 mg L−1 CuSO4·5H2O; 2 mg L−1 Na2Mo4·2H2O; 3 mg L−1 FeSO4·7H2O; 3.73 mg L−1 EDTANa2) [57] which was added once a week after 14 DPI. Two independent assays were performed using a completely randomized design with six treatments and three biological replicates each: uninoculated, inoculated with Az, Ba, Gl or Ps bio-inputs, and inoculated with Mz bio-input (U, Az, Ba, Gl, Ps and Mz treatments, respectively). Samples were taken on different days to evaluate root colonization, biomass accumulation and leaf area as described below.
- -
- Lettuce seedlings grown in nursery greenhouse conditions: Following the workflow usually carried out by the commercial plant nursery, Baby Plant S.A. (−34.980211, −58.031761, La Plata, Argentina), for cropping lettuce seedlings, nondisinfected lettuce seeds were sowed with a pneumatic sowing machine in seedling trays (volume of 10 mL per cell) with nonsterile peat-based substrate (Klasmann Ts1). Inoculation was performed as previously described for seedlings grown in trays in controlled conditions. Seedling trays were kept in nursery greenhouse conditions and irrigated on demand throughout the test. Two independent assays (October 2018 and September 2019) were carried out using a completely randomized design with six treatments and three biological replicates each: uninoculated, inoculated with Az, Ba, Gl or Ps bio-inputs, and inoculated with Mz bio-input (U, Az, Ba, Gl, Ps and Mz treatments, respectively). Biomass accumulation was evaluated as described below when seedlings were ready for transplanting (30–35 DPI and 4 to 6 developed leaves).
4.4.2. Lettuce Plants at Productive Stage
4.5. Plant-Root Colonization Assessment
4.6. Plant-Growth Promotion Assessment
4.7. Statistical Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dong, C.J.; Wang, L.L.; Li, Q.; Shang, Q.M. Bacterial communities in the rhizosphere, phyllosphere and endosphere of tomato plants. PLoS ONE 2019, 14, e0223847. [Google Scholar] [CrossRef] [PubMed]
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Bashan, Y.; de-Bashan, L.E.; Prabhu, S.R.; Hernandez, J.P. Advances in plant growth-promoting bacterial inoculant technology: Formulations and practical perspectives (1998–2013). Plant Soil 2014, 378, 1–33. [Google Scholar] [CrossRef]
- Compant, S.; Clément, C.; Sessitsch, A. Plant growth promoting bacteria in the rhizo and endosphere of plants: Their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Biochem. 2010, 42, 669–678. [Google Scholar] [CrossRef]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef]
- Supratim, B.; Rabara, R.; Negi, S. Towards a better greener future—An alternative strategy using biofertilizers. I: Plant growth promoting bacteria. Plant Gene 2017, 12, 43–49. [Google Scholar] [CrossRef]
- Starobinsky, G.; Monzón, J.; Di Marzo Broggi, E.; Braude, E. Bioinsumos para la Agricultura que Demandan Esfuerzos de Investigación y Desarrollo. Capacidades Existentes y Estrategia de Política Pública para Impulsar su Desarrollo en Argentina. Documentos de Trabajo del CCE N° 17. 2021, pp. 1–137. Available online: https://www.argentina.gob.ar/sites/default/files/2021/03/dt_17_-_bioinsumos.pdf (accessed on 12 December 2022).
- Molina-Romero, D.; Baez, A.; Quintero-Hernández, V.; Castañeda-Lucio, M.; Fuentes-Ramírez, L.E.; Bustillos-Cristales, M.d.R.; Rodríguez-Andrade, O.; Morales-García, Y.E.; Munive, A.; Muñoz-Rojas, J.; et al. Compatible bacterial mixture, tolerant to desiccation, improves maize plant growth. PLoS ONE 2017, 12, e0187913. [Google Scholar] [CrossRef]
- Thomloudi, E.E.; Tsalgatidou, P.C.; Douka, D.; Spantidos, T.N.; Dimou, M.; Venieraki, A.; Katinakis, P. Multistrain versus Single-Strain Plant Growth Promoting Microbial Inoculants-The Compatibility Issue. Hell. Plant Prot. J. 2019, 12, 61–77. [Google Scholar] [CrossRef]
- Vivanco-Calixto, R.; Molina-Romero, D.; Morales-García, Y.E.; Quintero-Hernández, V.; Munive-Hernández, J.A.; Baez-Rogelio, A.; Muñoz-Rojas, J. Reto agrobiotecnológico: Inoculantes bacterianos de segunda generación. Alianzas Tend. BUAP 2016, 1, 1–10. Available online: https://www.aytbuap.mx/publicaciones#h.26a62fnd2t88 (accessed on 14 December 2022).
- Ansari, F.A.; Ahmad, I. Fluorescent Pseudomonas -FAP2 and Bacillus licheniformis interact positively in biofilm mode enhancing plant growth and photosynthetic attributes. Sci. Rep. 2019, 9, 4547. [Google Scholar] [CrossRef]
- Romita, D.A.; Sharmab, G.D.; Majumdarc, P.B.; Pandey, P. A multispecies consortium of bacteria having plant growth promotion and antifungal activities, for the management of Fusarium wilt complex disease in potato (Solanum tuberosum L.). Biocatal. Agric. Biotechnol. 2018, 16, 614–624. [Google Scholar] [CrossRef]
- Gómez-Godínez, L.J.; Fernandez-Valverde, S.L.; Martinez Romero, J.C.; Martínez-Romero, E. Metatranscriptomics and nitrogen fixation from the rhizoplane of maize plantlets inoculated with a group of PGPRs. Syst. Appl. Microbiol. 2019, 42, 517–525. [Google Scholar] [CrossRef]
- Hett, J.; Neuhoff, D.; Döring, T.F.; Masoero, G.; Ercole, E.; Bevivino, A. Effects of multi-species microbial inoculants on early wheat growth and litterbag microbial activity. Agronomy 2022, 12, 899. [Google Scholar] [CrossRef]
- Iqbal, A.; Iqbal, M.A.; Iqbal, A.; Aslam, Z.; Maqsood, M.; Ahmad, Z.; Akbar, N.; Khan, H.Z.; Abbas, R.N.; Khan, R.D.; et al. Boosting forage yield and quality of maize (Zea mays L.) with multi-species bacterial inoculation in Pakistan. Phyton-Int. J. Exp. Bot. 2017, 86, 84–88. [Google Scholar] [CrossRef]
- Tabacchioni, S.; Passato, S.; Ambrosino, P.; Huang, L.; Caldara, M.; Cantale, C.; Hett, J.; Del Fiore, A.; Fiore, A.; Schlüter, A.; et al. Identification of beneficial microbial consortia and bioactive compounds with potential as plant biostimulants for a sustainable agriculture. Microorganisms 2021, 9, 426. [Google Scholar] [CrossRef]
- Emmanuel, O.C.; Babalola, O.O. Productivity and quality of horticultural crops through co-inoculation of arbuscular mycorrhizal fungi and plant growth promoting bacteria. Microbiol. Res. 2020, 239, 126569. [Google Scholar] [CrossRef]
- Abdullah, K.R.; Mehdi, Z.; Angelika, A.; Rostislav, S.; Hassan, E. Chapter 1: Effects of microbial inoculants on growth, yield, and fruit quality under stress conditions. In Sustainable Horticulture: Microbial Inoculants and Stress Interaction, 1st ed.; Seymen, M., Kurtar, E.S., Erdinc, C., Kumar, A., Eds.; Elsevier: Amsterdan, The Netherland, 2022; pp. 1–38. [Google Scholar] [CrossRef]
- Kim, M.J.; Moon, Y.; Tou, J.C.; Mou, B.; Waterland, N.L. Nutritional value, bioactive compounds and health benefits of lettuce (Lactuca sativa L.). J. Food Compos. Anal. 2016, 49, 19–34. [Google Scholar] [CrossRef]
- Mercado Interno y Externo de Lechuga 2018. MAGyP, Presidencia de la Nación. Available online: https://www.magyp.gob.ar/sitio/areas/ss_mercados_agropecuarios/publicaciones/_archivos/000101_Perfiles/999984_Perfil%20del%20Mercadode%20la%20Lechuga%202018.pdf (accessed on 6 December 2022).
- Vio, S.A.; Bernabeu, P.R.; García, S.S.; Galar, M.L.; Luna, M.F. Tracking and plant growth-promoting effect of Paraburkholderia tropica MTo-293 applied to Solanum lycopersicum. J. Basic Microbiol. 2022, 62, 875–886. [Google Scholar] [CrossRef]
- Mitidieri, M.S.; Corbino, G.B. Manual de Horticultura Periurbana, 1ra Edición. Ediciones INTA 2012, San Pedro, Buenos Aires. Available online: https://inta.gob.ar/sites/default/files/script-tmp-manual_de_horticultura_urbana_y_periurbana.pdf (accessed on 25 November 2022).
- Bonaldi, M.; Chen, X.; Kunova, A.; Pizzatti, C.; Saracchi, M.; Cortesi, P. Colonization of lettuce rhizosphere and roots by tagged Streptomyces. Front. Microbiol. 2015, 6, 25. [Google Scholar] [CrossRef]
- Cipriano, M.A.P.; Lupatini, M.; Lopes-Santos, L.; da Silva, M.J.; Roesch, L.f.W.; Destéfano, S.A.L.; Freitas, S.S.; Kuramae, E.E. Lettuce and rhizosphere microbiome responses to growth promoting Pseudomonas species under field conditions. FEMS Microbiol. Ecol. 2016, 92, fiw197. [Google Scholar]
- Chamangasht, S.; Ardakani, M.R.; Khavazi, K.; Abbaszadeh, B.; Mafakheri, S. Improving lettuce (Lactuca sativa l.) growth and yield by the application of biofertilizers. Ann. Biol. Res. 2012, 3, 1876–1879. Available online: https://www.scholarsresearchlibrary.com/articles/improving-lettuce-lactuca-sativa-l-growth-and-yield-by-the-application-of-biofertilizers.pdf (accessed on 6 December 2022).
- Henriques dos Santos Moraes, M.C.; Valente de Medeiros, E.; da Silva de Andrade, D.; Dias de Lima, L.; da Silva Santos, I.C.; Martins Filho, A.P. Microbial biomass and enzymatic activities in sandy soil cultivated with lettuce inoculated with plant growth promoters. Rev. Caatinga 2018, 31, 860–870. [Google Scholar] [CrossRef]
- Szczech, M.; Szafirowska, A.; Kowalczyk, W.; Szwejda-Grzybowska, J.; Włodarek, A.; Maciorowski, R. The effect of plant growth promoting bacteria on transplants growth and lettuce yield in organic production. J. Hortic. Res. 2016, 24, 101–107. [Google Scholar] [CrossRef]
- Martins Filho, A.P.; Valente de Medeiros, E.; Gomes Barbosa, J.; Peniche Barbosa, J.M.; Kuklinsky-Sobral, J.; Souza-Motta, C. Combined effect of pseudomonas sp. and trichoderma aureoviride on lettuce growth promotion. Biosci. J. 2019, 35, 419–430. [Google Scholar] [CrossRef]
- Mamani de Marchese, A.; Filippone, M.P. Bio-products: Key components of sustainable agriculture. Rev. Agron. Noroeste Argent. 2018, 38, 9–21. Available online: http://www.scielo.org.ar/scielo.php?script=sci_abstract&pid=S2314-369X2018000100001&lng=pt&nrm=iso&tlng=es (accessed on 10 December 2022).
- Barassi, C.A.; Ayrault, G.; Creus, C.M.; Sueldo, R.J.; Sobrero, M.T. Seed inoculation with Azospirillum mitigates NaCl effects on lettuce. Sci. Hortic. 2006, 109, 8–14. [Google Scholar] [CrossRef]
- Fasciglione, G.; Casanovas, E.M.; Yommi, A.; Sueldo, R.J.; Barassi, C.A. Azospirillum improves lettuce growth and transplant under saline conditions. J. Sci. Food Agric. 2012, 92, 2518–2523. [Google Scholar] [CrossRef]
- Fasciglione, G.; Casanovas, E.M.; Quillehauquy, V.; Yommi, A.K.; Goñi, M.G.; Roura, S.I.; Barassi, C.A. Azospirillum inoculation effects on growth, product quality and storage life of lettuce plants grown under salt stress. Sci. Hortic. 2015, 195, 154–162. [Google Scholar] [CrossRef]
- Garbi, M.; Carletti, S.; Sillon, C.; Vita, F. Respuesta de plántulas de lechuga mantecosa (Lactuca sativa L.) a la inoculación con una formulación compuesta por tres cepas de Azospirillum brasilense. Hortic. Argent. 2016, 35, 19–28. Available online: https://www.horticulturaar.com.ar/es/articulos/respuesta-de-plantulas-de-lechuga-mantecosa-a-la-inoculacion-con-una-formulacion-compuesta-por-tres-cepas-de-azospirillum-brasilense.html (accessed on 30 November 2022).
- Maroniche, G.A.; Diaz, P.R.; Borrajo, M.P.; Valverde, C.F.; Creus, C.M. Friends or foes in the rhizosphere: Traits of fluorescent Pseudomonas that hinder Azospirillum brasilense growth and root colonization. FEMS Microbiol. Ecol. 2018, 94, fiy202. [Google Scholar] [CrossRef]
- Romano, I.; Ventorino, V.; Pepe, O. Effectiveness of plant beneficial microbes: Overview of the methodological approaches for the assessment of root colonization and persistence. Front. Plant Sci. 2020, 11, 6. [Google Scholar] [CrossRef]
- Cassán, F.; Diaz-Zorita, M. Azospirillum sp. in current agriculture: From the laboratory to the field. Soil Biol. Biochem. 2016, 103, 117–130. [Google Scholar] [CrossRef]
- Dwivedi, M. Chapter 25: Gluconacetobacter. In Beneficial Microbes in Agro-Ecology: Bacteria and Fungi, 1st ed.; Amaresan, N., Senthil Kumar, M., Annapurna, K., Kumar, K., Sankaranarayanan, A., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 521–544. [Google Scholar] [CrossRef]
- Krishnashis, D.; Shrutica, A.; Renu, V.; Harika, A.; Nalini, K.; Senthilkumar, M. Chapter 8: Pseudomonas. In Beneficial Microbes in Agro-Ecology: Bacteria and Fungi, 1st ed.; Amaresan, N., Senthil Kumar, M., Annapurna, K., Kumar, K., Sankaranarayanan, A., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 133–148. [Google Scholar] [CrossRef]
- Rainer, B. Chapter 8: Bacillus. In Beneficial Microbes in Agro-Ecology: Bacteria and Fungi, 1st ed.; Amaresan, N., Senthil Kumar, M., Annapurna, K., Kumar, K., Sankaranarayanan, A., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 107–132. [Google Scholar] [CrossRef]
- Prasad, A.A.; Babu, S. Compatibility of Azospirillum brasilense and Pseudomonas fluorescens in growth promotion of groundnut (Arachis hypogea L.). Acad. Bras. Cienc. 2017, 89, 1027–1040. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Bassel, G.W. Seed vigour and crop establishment: Extending performance beyond adaptation. J. Exp. Bot. 2016, 67, 567–591. [Google Scholar] [CrossRef]
- Rodríguez Cáceres, E. Improved medium for isolation of Azospirillum spp. Appl. Environ. Microbiol. 1982, 44, 990–991. [Google Scholar] [CrossRef]
- dos Santos Ferreira, N.; Coniglio, A.; Puente, M.; Hayashi Sant’Anna, F.; Maroniche, G.; García, J.; Molina, R.; Nievas, S.; Gazolla Volpiano, C.; Ambrosini, A.; et al. Genome-based reclassification of Azospirillum brasilense Az39 as the type strain of Azospirillum argentinense sp. nov. Int. J. Syst. Evol. Microbiol. 2022, 72, 005475. [Google Scholar] [CrossRef]
- Howell, C.R.; Stipanovic, R.D. Control of Rhizoctonia solani on cotton seedlings with Pseudomonas fluorescens and with an antibiotic produced by the bacterium. Phytopathology 1979, 69, 480–482. [Google Scholar] [CrossRef]
- Ramette, A.; Frapolli, M.; Fischer-Le Saux, M.; Gruffaz, C.; Meyer, J.M.; Défago, G.; Sutra, L.; Moënne-Loccoz, Y. Pseudomonas protegens sp. nov., widespread plant-protecting bacteria producing the biocontrol compounds 2,4-diacetylphloroglucinol and pyoluteorin. Syst. Appl. Microbiol. 2011, 34, 180–188. [Google Scholar] [CrossRef]
- Cavalcante, V.A.; Döbereiner, J. A new acid tolerant nitrogen-fixing bacterium associated with sugar cane. Plant Soil 1988, 108, 23–31. [Google Scholar] [CrossRef]
- Yamada, Y.; Hoshino, K.; Ishikawa, T. The phylogeny of acetic acid bacteria based on the partial sequences of 16S ribosomal RNA: The elevation of the subgenus Gluconoacetobacter to the generic level. Biosci. Biotechnol. Biochem. 1997, 61, 1244–1251. [Google Scholar] [CrossRef]
- Toledo, A.V.; López, S.M.Y.; Aulacino, M.; Marino, A.M.; Balatti, P. Antagonism of entomopathogenic fungi by Bacillus spp. associated with the integument of cicadellids and delphacids. Int. Microbiol. 2015, 18, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Stephan, M.P.; Oliveira, M.; Teixeira, K.R.S.; Martinez-Drets, G.; Döbereiner, J. Physiology and dinitrogen fixation of Acetobacter diazotrophicus. FEMS Microbiol. Lett. 1991, 77, 67–72. [Google Scholar] [CrossRef]
- Mazza, P.; Zani, F.; Martelli, P. Studies on the antibiotic resistance of Bacillus subtilis strains used in oral bacteriotherapy. Boll. Chim. Farm. 1992, 131. [Google Scholar]
- Luna, M.F.; Aprea, J.; Crespo, J.M.; Boiardi, J.L. Colonization and yield promotion of tomato by Gluconacetobacter diazotrophicus. Appl Soil Ecol. 2012, 61, 225–229. [Google Scholar] [CrossRef]
- Rai, R.; Prasad, V.; Shukla, I.C. Interaction between finger millet (Eleusine coracana) genotypes and drug-resistant mutants of Azospirillum brasilense in calcareous soil. J. Agric. Sci. 1984, 102, 521. [Google Scholar] [CrossRef]
- Janssen, P.H.; Yates, P.S.; Grinton, B.E.; Taylor, P.M.; Sait, M. Improved culturability of soil bacteria and isolation in pure culture of novel members of the divisions acidobacteria, actinobacteria, proteobacteria, and verrucomicrobia. Appl. Environ. Microbiol. 2002, 68, 2391–2396. [Google Scholar] [CrossRef]
- García, S.S.; Bernabeu, P.R.; Vio, S.A.; Cattelan, N.; García, J.E.; Puente, M.L.; Galar, M.L.; Prieto, C.I.; Luna, M.F. Paraburkholderia tropica as a plant- growth–promoting bacterium in barley: Characterization of tissues colonization by culture-dependent and-independent techniques for use as an agronomic bioinput. Plant Soil 2019, 451, 89–106. [Google Scholar] [CrossRef]
- Fähraeus, G. The infection of clover root hairs by nodule bacteria studied by a simple glass slide technique. Microbiology 1957, 16, 374–381. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Calif. Agric. Exp. Stn. 1950, 347, 1–32. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Bremner, J.M. Determination of nitrogen in soil by the Kjeldahl method. J. Agric. Sci. 1960, 55, 11–33. [Google Scholar] [CrossRef]
- Bernabeu, P.R.; Pistorio, M.; Torres-Tejerizo, G.; Estrada-de Los Santos, P.; Galar, M.L.; Boiardi, J.L.; Luna, M.F. Colonization and plant growth-promotion of tomato by Burkholderia tropica. Sci. Hortic. 2015, 191, 113–120. [Google Scholar] [CrossRef]
- Rasband, W.S. ImageJ 1997–2016, U.S. National Institutes of Health, Bethesda, Maryland, USA. Available online: https://imagej.nih.gov/ij (accessed on 15 November 2022).
- di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C. Infostat versión 2018. Grupo Infostat, FCA. Córdoba, Argentina: Universidad Nacional de Córdoba, Argentina. Available online: https://www.infostat.com.ar/ (accessed on 10 November 2022).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System | Time | Single-Strain Bio-Input 1 | Mz Bio-Input 2 | Uninoculated | |||
|---|---|---|---|---|---|---|---|
| Az | Ba | Gl | Ps | ||||
| Aqueous formulation | 0 h | 7.01 | 8.30 | 6.16 | 7.58 | 7.67 | -- |
| 24 h | 6.91 | 8.22 | 6.26 | 7.54 | 7.48 | ||
| Substrate | 0 h | 6.45 | 6.47 | 6.50 | 6.50 | 6.74 | 6.27 |
| 24 h | 6.75 | 6.73 | 6.60 | 6.40 | 6.76 | 6.26 | |
| Strain | Plant-Growth Promoting Traits [36,37,38,39] | Antibiotic Resistance 1 | Culture Media 2 | pH Initial/Final | Concentration 3 |
|---|---|---|---|---|---|
| Azospirillum argentinense Az39 | -Phytohormone production -Nitrogen fixation | Kanamycin | CR | 6.8/7.0 | ~109 |
| Bacillus sp. Dm-B10 | -Biocontrol activity | Rifampicin | LB | 7.0/8.3 | ~108 |
| Gluconacetobacter diazotrophicus PAL5 | -Nitrogen fixation -Phosphate solubilization | Nalidixin | LGI | 6.0/6.16 | ~109 |
| Pseudomonas protegens Pf-5 | -Phosphate solubilization -Biocontrol activity | Streptomycin | LB | 7.0/7.6 | ~109 |
| Analysis 1 | Units | EE JH | EE Gorina |
|---|---|---|---|
| pH | 6.81 | 7.49 | |
| EC | [dS.m−1] | 1.28 | 1.25 |
| C | [%] | 2.54 | 1.39 |
| OM | [%] | 4.38 | 2.40 |
| Nt | [%] | 0.242 | 0.134 |
| C/N | 10 | 10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vio, S.A.; Galar, M.L.; Gortari, M.C.; Balatti, P.; Garbi, M.; Lodeiro, A.R.; Luna, M.F. Multispecies Bacterial Bio-Input: Tracking and Plant-Growth-Promoting Effect on Lettuce var. sagess. Plants 2023, 12, 736. https://doi.org/10.3390/plants12040736
Vio SA, Galar ML, Gortari MC, Balatti P, Garbi M, Lodeiro AR, Luna MF. Multispecies Bacterial Bio-Input: Tracking and Plant-Growth-Promoting Effect on Lettuce var. sagess. Plants. 2023; 12(4):736. https://doi.org/10.3390/plants12040736
Chicago/Turabian StyleVio, Santiago A., María Lina Galar, María Cecilia Gortari, Pedro Balatti, Mariana Garbi, Aníbal Roberto Lodeiro, and María Flavia Luna. 2023. "Multispecies Bacterial Bio-Input: Tracking and Plant-Growth-Promoting Effect on Lettuce var. sagess" Plants 12, no. 4: 736. https://doi.org/10.3390/plants12040736
APA StyleVio, S. A., Galar, M. L., Gortari, M. C., Balatti, P., Garbi, M., Lodeiro, A. R., & Luna, M. F. (2023). Multispecies Bacterial Bio-Input: Tracking and Plant-Growth-Promoting Effect on Lettuce var. sagess. Plants, 12(4), 736. https://doi.org/10.3390/plants12040736