

Effects of Different Sulfur Compounds on the Distribution Characteristics of Subcellular Lead Content in Arabis alpina L. var. parviflora Franch under Lead Stress
Abstract
:1. Introduction
2. Results
2.1. The Effect of Different Forms of Sulfur on the Biomass of A. alpina
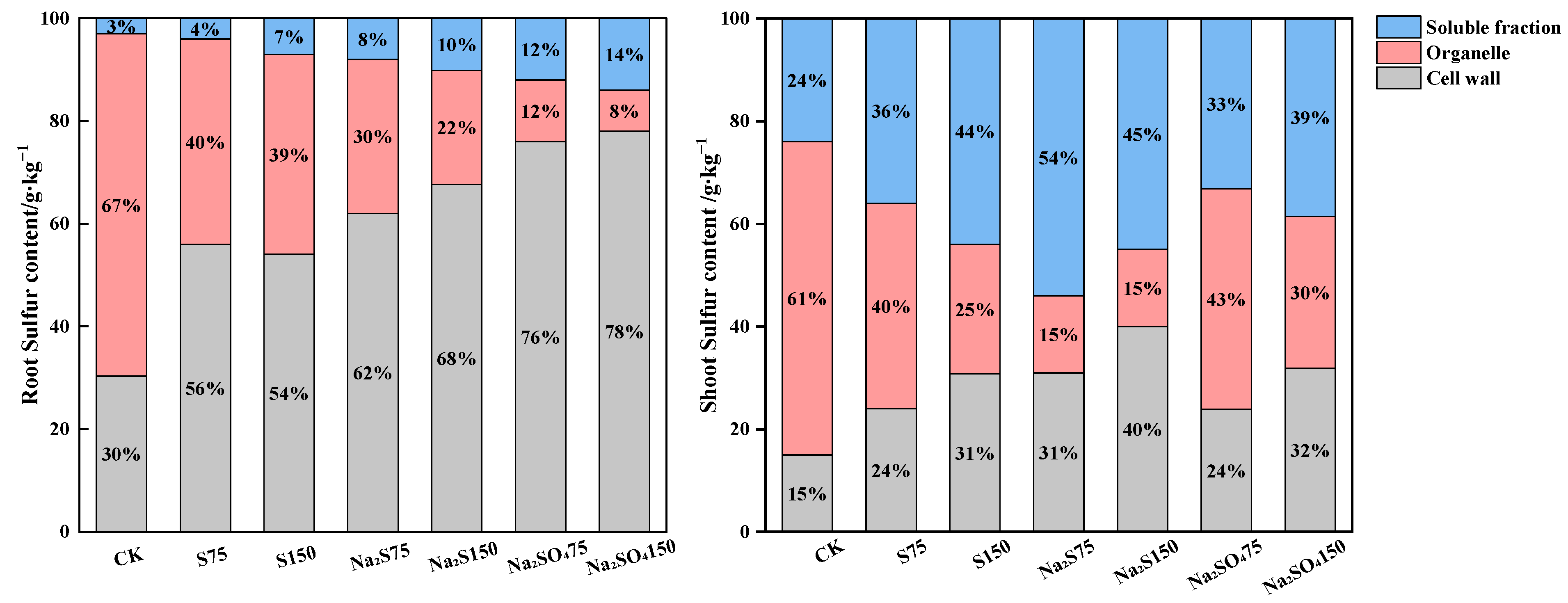
2.2. Effects of Different Forms of Sulfur on the Sulfur Contents in Subcellular Part of A. alpina
2.3. Effects of Different Forms of Sulfur on Lead Content in Subcellular Part of A. alpina
2.4. Correlation of Sulfur Content and Lead Content in Subcellular Parts of A. alpina
3. Discussion
4. Materials and Methods
4.1. Experimental Design and Materials
4.2. Indicator Determination
4.2.1. Soil Determination of Physical and Chemical Properties
4.2.2. Determination of Biomass
4.2.3. Determination of Subcellular Sulfur and Lead Content
4.2.4. Determination of Total Lead Content in the Plant and Soil
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Entwistle, J.A.; Amaibi, P.A.; Dean, J.D.; Deary, M.E.; Medock, D.; Morton, J.; Rodushkin, I.; Bramwell, L. An apple a day? Assessing gardeners’ lead exposure in urban agriculture sites to improve the derivation of soil assessment criteria. Environ. Int. 2019, 122, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Li, X.X.; Lan, X.; Liu, W.; Cui, X.W.; Cui, Z.J. Toxicity, migration and transformation characteristics of lead in soil-plant system: Effect of lead species. J. Hazard. Mater. 2020, 395, 122676. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.L.; Lu, J.Y.; Jia, S.L. Effects of sulfur on ascorbate-glutathione cycle and the content of phytochelatins in the leaves of Pakchoi (Brassica chinensis L.) under cadmium stress. J. Agro-Environ. Sci. 2013, 32, 1294–1301. [Google Scholar] [CrossRef]
- Zakari, S.; Jiang, X.J.; Zhu, X.A.; Liu, W.J.; Allakonon, M.G.B.; Singh, S.K.; Chen, C.F.; Zou, X.; Akponikpè, P.B.I.; Dossa, G.G.O.; et al. Influence of sulfur amendments on heavy metals phytoextraction from agricultural contaminated soils: A meta-analysis. Environ. Pollut. 2021, 288, 117820. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.; Feng, J.B.; Shi, J.Y. Impact of sulfur (S) fertilization in paddy soils on copper (Cu) accumulation in rice (Oryza sativa L.) plants under flooding conditions. Biol. Fertil. Soils 2016, 52, 31–39. [Google Scholar] [CrossRef]
- Wang, Y.P.; Li, Q.B.; Wang, H.; Shi, J.Y. Effect of sulfur on soil Cu/Zn availability and microbial community composition. J. Hazard. Mater. 2008, 159, 385–389. [Google Scholar] [CrossRef]
- Li, Y.; He, J.; Luo, L.; Wang, Y. The Combinations of sulfur and molybdenum fertilization improved antioxidant capacity in grazing Nanjiang brown goat. Biol. Trace Elem. Res. 2022, 200, 600–608. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, S.M.; Wang, J.X.; Swertz, A.C.; Feng, X.B.; Bolan, N.; Rinklebe, J. Enhancing phytoextraction of potentially toxic elements in a polluted floodplain soil using sulfur-impregnated organoclay. Environ. Pollut. 2019, 248, 1059–1066. [Google Scholar] [CrossRef]
- Ren, C.; Qi, Y.B.; Huang, G.Y.; Hu, H.Q. Effects of sulfur on toxicity and bioavailability of Cu for castor (Ricinus communis L.) in Cu-contaminated soil. Environ. Sci. Pollut. Res. 2017, 24, 27476–27483. [Google Scholar] [CrossRef] [PubMed]
- Karimi, A.N.; Souri, Z. Exogenous supplementation of sulfur (S) and reduced glutathione (GSH) alleviates arsenic toxicity in shoots of Isatis cappadocica Desv and Erysimum allionii L. Environ. Sci. Pollut. Res. 2022, 29, 64205–64214. [Google Scholar] [CrossRef]
- Shahbza, M.; Tseng, M.H.; Stuiver, C.E.E.; Koraleaska, A.; Posthumus, J.H.V.; Venema, J.H.; Parmar, S.; Schat, H.; Hawkesford, N.J.; Kok, L.J.D. Copper exposure interferes with the regulation of the uptake, distribution, and metabolism of sulfate in Chinese cabbage. J. Plant Physiol. 2010, 167, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Pu, P.; Li, X.R.; Gong, Y.Z.; An, D.S.; Zhang, L.X.; Lv, J.Y. Sulfur application reduces cadmium uptake in edible parts of pakchoi (Brassica chinensis L.) by cadmium chelation and vacuolar sequestration. Ecotoxicol. Environ. Saf. 2020, 194, 110402. [Google Scholar] [CrossRef]
- Das, U.; Rahman, M.A.; Ela, E.J.; Lee, K.W.; Kabir, A.H. Sulfur triggers glutathione and phytochelatin accumulation causing excess Cd bound to the cell wall of roots in alleviating Cd-toxicity in alfalfa. Chemosphere 2020, 262, 128361. [Google Scholar] [CrossRef] [PubMed]
- Li, D.Q.; Chen, G.K.; Lu, Q.; Li, Y.T.; Wang, J.J.; Li, H.S. Responses of two kidney bean (Phaseolus vulgaris) cultivars to the combined stress of sulfur deficiency and cadmium toxicity. Biometals 2018, 31, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Hao, M.; Liu, Y.H.; Huang, G.Y.; Fu, Q.L.; Zhu, J.; Hu, H.Q. Effects of exogenous sulfur on growth and Cd uptake in Chinese cabbage (Brassica campestris spp. pekinensis) in Cd-contaminated soil. Environ. Sci. Pollut. Res. 2018, 25, 15823–15829. [Google Scholar] [CrossRef]
- Küpper, H.; Lombi, E.; Mcgrath, Z.S.P. Cellular compartmentation of cadmium and zinc in relation to other elements in the hyperaccumulator Arabidopsis helleri. Plant 2000, 212, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Li, H.H.; Yang, X.E. Effects of sulfur on accumulation, subcellular distribution and chemical forms of cadmium in hyperaccumulator-Sedum alfredii Hance. Plant Nutr. Fertil. Sci. 2009, 15, 395–402. [Google Scholar] [CrossRef]
- Lin, H.R.; Shi, J.Y.; Chen, X.C.; Yang, J.J.; Chen, Y.X.; Zhao, Y.D.; Hu, T.D. Sulfate-reducing bacteria existed in the rice rhi-zosphere affected by lead and thiosulfate interaction. Soil Biol. Biochem. 2010, 42, 1038–1044. [Google Scholar] [CrossRef]
- Li, Z.R.; Golinet, G.; Zu, Y.Q.; Wang, J.X.; An, L.Z.; Li, Q.; Niu, X.Y. Species diversity of Arabis alpina L. communities in two Pb/Zn mining areas with different smelting history in Yunnan Province, China. Chemosphere 2019, 05, 228. [Google Scholar] [CrossRef] [PubMed]
- Mou, F.L.; Yang, J.M.; Liu, C.; Liu, M.; Chen, J.J.; Zu, Y.Q.; Wang, J.X. Effects of sulfur on the characteristics of sulfur-containing compounds and Pb accumulation in Arabis alpina L. var. parviflora Franch. J. Agro-Environ. Sci. 2021, 40, 1851–1859. [Google Scholar] [CrossRef]
- Buchner, P.; Takahashi, H.J.; Hawkesford, M. Plant sulphate transporters: Co-ordination of uptake, intracellular and long-distance transport. J. Exp. Bot. 2004, 404, 1765–1773. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.N.; Lai, H.X.; Wen, X.L.; Li, X.P. Study on microflora in the remediation of saline-alkali soil by addition of sulfur. J. Northwest A F Univ. Nat. Sci. Ed. 2010, 38, 153–157. [Google Scholar] [CrossRef]
- Xu, Y.; Mu, J.M.; Zhang, G.Q.; Ma, J.J.; Xu, J.; Li, J.; Liu, F.J.; She, X.D. Effect of sulfur availability on nitrate accumulation and expression of nitrogen and sulfur assimilation related genes in non-heading Chinese cabbage. Sci. Agric. Sin. 2016, 49, 2222–2233. [Google Scholar] [CrossRef]
- Zhang, X.; Lou, X.; Zhang, H.; Ren, W.; Tang, M. Effects of sodium sulfide application on the growth of Robinia pseudoacacia, heavy metal immobilization, and soil microbial activity in Pb-Zn polluted soil. Ecotoxicol. Environ. Saf. 2020, 197, 110563. [Google Scholar] [CrossRef]
- Mikkelsen, R.L.; Wan, H.F. The effect of selenium on sulfur uptake by barley and rice. Plant Soil 1990, 121, 151–153. [Google Scholar] [CrossRef]
- Parrotta, L. Target or barrier? The cell wall of early- and later-diverging plants vs cadmium toxicity: Differences in the response mechanisms. Front. Plant Sci. 2015, 6, 133. [Google Scholar] [CrossRef] [Green Version]
- Xin, J.P.; Zhang, Y.; Tian, R.N. Tolerance mechanism of Triarrhena sacchariflora (Maxim.) Nakai. seedlings to lead and cadmium: Translocation, subcellular distribution, chemical forms and variations in leaf ultrastructure. Ecotoxicol. Environ. Saf. 2018, 165, 611–621. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Zhu, Y.N.; Hu, C. Subcellular distribution of chromium in accumulating plant Leersia hexandra Swartz. Plant Soil 2009, 322, 187–195. [Google Scholar] [CrossRef]
- Krzesłowska, M. The cell wall in plant cell response to trace metals: Polysaccharide remodeling and its role in defense strategy. Acta Physiol. Plant 2011, 33, 35–51. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Cao, X.F.; Li, X.X.; Li, T.; Zhang, H.; Cui, X.W.; Cui, Z.J. Ecological toxicity of Cd, Pb, Zn, Hg and regulation mechanism in Solanum nigrum L. Chemosphere 2023, 313, 137447. [Google Scholar] [CrossRef]
- Liao, Y.M.; Li, Z.R.; Yang, Z.C.; Wang, J.X.; Li, B.; Zu, Y.Q. Response of Cd, Zn Translocation and Distribution to Organic Acids Heterogeneity in Brassica juncea L. Plants 2023, 12, 479. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Yuan, Y.; Wang, X.M.; Wang, L.; Tao, Y.; Zhao, J.; Yu, H.; Dong, M.; Zhang, Y. Stabilization of lead and cadmium in soil by sulfur-iron functionalized biochar: Performance, mechanisms and microbial community evolution. J. Hazard. Mater. 2021, 425, 127876. [Google Scholar] [CrossRef]
- Wang, J.X.; Tai, G.C.; ZU, Y.Q.; Li, Y.; Zhan, F.D. Effects of sulfur application on growth and lead and zinc accumulation contents in Arabis alpinal var. parviflora franch. J. Agro-Environ. Sci. 2011, 30, 1064–1069. [Google Scholar]
- Bao, S.D. Agricultural Soil Analysis, 3rd ed.; China Agricuture Press: Beijing, China, 2000. [Google Scholar]
- Wang, Y.P.; Lin, Q.T.; Xiao, R.B.; Cheng, S.L.; Luo, H.Y.; Wen, X.Q.; Wu, L.B.; Zhong, Q.F. Removal of Cu and Pb from contaminated agricultural soil using mixed chelators of fulvic acid potassium and citric acid. Ecotoxicol. Environ. Saf. 2020, 206, 111179. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Sulfur Forms | Treatment Concentration /mg·kg−1 | Root /g·pot−1 | Shoot /g·pot−1 | Ratio of Root Biomass to Shoot Biomass |
|---|---|---|---|---|
| CK | 0 | 0.17 ± 0.06 d | 0.57 ± 0.10 d | 0.30 |
| S | 75 | 0.20 ± 0.05 d | 0.70 ± 0.06 d | 0.29 |
| 150 | 0.24 ± 0.03 d | 0.58 ± 0.07 cd | 0.41 | |
| Na2S | 75 | 0.32 ± 0.03 c | 0.71 ± 0.02 cd | 0.45 |
| 150 | 0.40 ± 0.05 b | 0.77 ± 0.06 c | 0.53 | |
| Na2SO4 | 75 | 0.52 ± 0.03 a | 0.91 ± 0.04 b | 0.57 |
| 150 | 0.57 ± 0.04 a | 1.16 ± 0.18 a | 0.49 |
| Treatment | Root/g·kg−1 | Shoot/g·kg−1 | |||||
|---|---|---|---|---|---|---|---|
| Sulfur Forms | Concentration /mg·kg−1 | Cell Wall | Organelle | Soluble Fraction | Cell Wall | Organelle | Soluble Fraction |
| CK | 0 | 1.03 ± 0.06 d | 2.28 ± 0.56 a | 0.12 ± 0.13 e | 0.79 ± 0.16 d | 3.34 ± 0.23 a | 1.31 ± 0.07 e |
| S | 75 | 1.74 ± 0.09 cd | 1.24 ± 0.14 b | 0.14 ± 0.08 e | 1.03 ± 0.12 cd | 1.71 ± 0.49 b | 1.54 ± 0.11 de |
| 150 | 1.30 ± 0.05 d | 0.95 ± 0.07 bc | 0.17 ± 0.09 de | 1.07 ± 0.11 cd | 0.86 ± 0.13 c | 1.87 ± 0.09 d | |
| Na2S | 75 | 1.68 ± 0.27 cd | 0.80 ± 0.04 bc | 0.22 ± 0.21 d | 1.39 ± 0.19 c | 0.67 ± 0.07 cd | 2.45 ± 0.35 c |
| 150 | 2.22 ± 0.19 c | 0.74 ± 0.06 bc | 0.34 ± 0.37 c | 1.44 ± 0.09 c | 0.53 ± 0.03 cd | 2.49 ± 0.45 c | |
| Na2SO4 | 75 | 3.04 ± 0.48 b | 0.48 ± 0.08 c | 0.47 ± 0.20 b | 2.41 ± 0.25 b | 0.46 ± 0.02 cd | 3.34 ± 0.23 b |
| 150 | 3.76 ± 0.32 a | 0.39 ± 0.05 c | 0.69 ± 0.85 a | 3.62 ± 0.44 a | 0.36 ± 0.04 d | 4.39 ± 0.42 a | |
| Treatment | Root/mg·kg−1 | Shoot/mg·kg−1 | |||||
|---|---|---|---|---|---|---|---|
| Sulfur Form | Concentration /mg·kg−1 | Cell Wall | Organelle | Soluble Fraction | Cell Wall | Organelle | Soluble Fraction |
| CK | 0 | 87.39 ± 6.6 d | 244.66 ± 33.6 a | 457.65 ± 30.5 a | 297.29 ± 39.3 d | 256.12 ± 27.8 a | 192.05 ± 46.3 f |
| S | 75 | 137.90 ± 21.7 c | 174.89 ± 28.3 b | 406.93 ± 18.5 ab | 381.42 ± 24.2 cd | 260.02 ± 24.2 a | 260.75 ± 33.0 ef |
| 150 | 173.99 ± 27.3 c | 170.61 ± 16.6 b | 349.73 ± 28.3 b | 471.15 ± 82.8 c | 199.28 ± 23.3 b | 334.71 ± 43.3 de | |
| Na2S | 75 | 133.61 ± 17.8 c | 142.72 ± 52.4 bc | 263.92 ± 38.8 c | 634.60 ± 52.0 b | 144.05 ± 13.7 c | 374.52 ± 26.5 d |
| 150 | 175.52 ± 20.6 c | 134.36 ± 14.1 c | 277.19 ± 55.6 c | 706.78 ± 19.4 ab | 104.58 ± 9.8 d | 458.35 ± 34.1 c | |
| Na2SO4 | 75 | 241.33 ± 27.9 b | 134.18 ± 74.3 c | 218.64 ± 31.8 cd | 798.60 ± 67.2 a | 80.17 ± 12.2 de | 539.99 ± 56.0 b |
| 150 | 309.92 ± 38.0 a | 89.85 ± 14.8 d | 185.70 ± 17.5 d | 817.66 ± 101.1 a | 67.80 ± 5.8 e | 664.89 ± 54.9 a | |
| Treatment | Root /mg·kg−1 | Shoot /mg·kg−1 | BCF | TF | |
|---|---|---|---|---|---|
| Sulfur Forms | Concentration /mg·kg−1 | ||||
| CK | 0 | 789.7 | 745.46 | 0.28 | 0.94 |
| S | 75 | 719.72 | 902.19 | 0.34 | 1.25 |
| 150 | 694.33 | 1005.14 | 0.38 | 1.44 | |
| Na2S | 75 | 540.25 | 1153.17 | 0.44 | 2.13 |
| 150 | 587.07 | 1269.71 | 0.48 | 2.16 | |
| Na2SO4 | 75 | 594.15 | 1418.76 | 0.54 | 2.38 |
| 150 | 585.47 | 1550.35 | 0.59 | 2.64 | |
| Treatment | Root | Shoot | |||||
|---|---|---|---|---|---|---|---|
| Sulfur Forms | Concentration /mg·kg−1 | Cell Wall | Organelle | Soluble Fraction | Cell Wall | Organelle | Soluble Fraction |
| CK | 0 | 0.751 | 0.692 | 0.633 | 0.512 | 0.044 | 0.479 |
| S | 75 | 0.968 * | 0.971 * | 0.994 * | 0.281 | 0.455 | 0.822 |
| 150 | 0.997 * | 0.997 * | 0.982 * | 0.929 | 0.436 | 0.651 | |
| Na2S | 75 | 0.671 | 0.835 | 0.967 * | 0.422 | 0.536 | 0.178 |
| 150 | 0.996 * | 0.956 * | 0.864 | 0.649 | 0.195 | 0.112 | |
| Na2SO4 | 75 | 0.975 * | 0.984 * | 0.989 * | 0.999 * | 0.517 | 0.495 |
| 150 | 0.991 * | 0.973 * | 0.962 * | 0.964 * | 0.197 | 0.728 | |
| pH | Available Sulfur /mg·kg−1 | Pb /mg·kg−1 | SOM /g·kg1 | AP /mg·kg−1 | AN /mg·kg−1 | AK /mg·kg−1 | CEC /cmol·kg−1 |
|---|---|---|---|---|---|---|---|
| 6.25 | 10.67 | 2618.74 | 16.87 | 12.08 | 116.34 | 28.77 | 16.45 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, C.; Qin, L.; Li, Y.; Zu, Y.; Wang, J. Effects of Different Sulfur Compounds on the Distribution Characteristics of Subcellular Lead Content in Arabis alpina L. var. parviflora Franch under Lead Stress. Plants 2023, 12, 874. https://doi.org/10.3390/plants12040874
Xu C, Qin L, Li Y, Zu Y, Wang J. Effects of Different Sulfur Compounds on the Distribution Characteristics of Subcellular Lead Content in Arabis alpina L. var. parviflora Franch under Lead Stress. Plants. 2023; 12(4):874. https://doi.org/10.3390/plants12040874
Chicago/Turabian StyleXu, Cui, Li Qin, Yuan Li, Yanqun Zu, and Jixiu Wang. 2023. "Effects of Different Sulfur Compounds on the Distribution Characteristics of Subcellular Lead Content in Arabis alpina L. var. parviflora Franch under Lead Stress" Plants 12, no. 4: 874. https://doi.org/10.3390/plants12040874
APA StyleXu, C., Qin, L., Li, Y., Zu, Y., & Wang, J. (2023). Effects of Different Sulfur Compounds on the Distribution Characteristics of Subcellular Lead Content in Arabis alpina L. var. parviflora Franch under Lead Stress. Plants, 12(4), 874. https://doi.org/10.3390/plants12040874