

Seed Germination of Three Milk Thistle (Silybum marianum (L.) Gaertn.) Populations of Greek Origin: Temperature, Duration, and Storage Conditions Effects




Abstract
:1. Introduction
2. Results
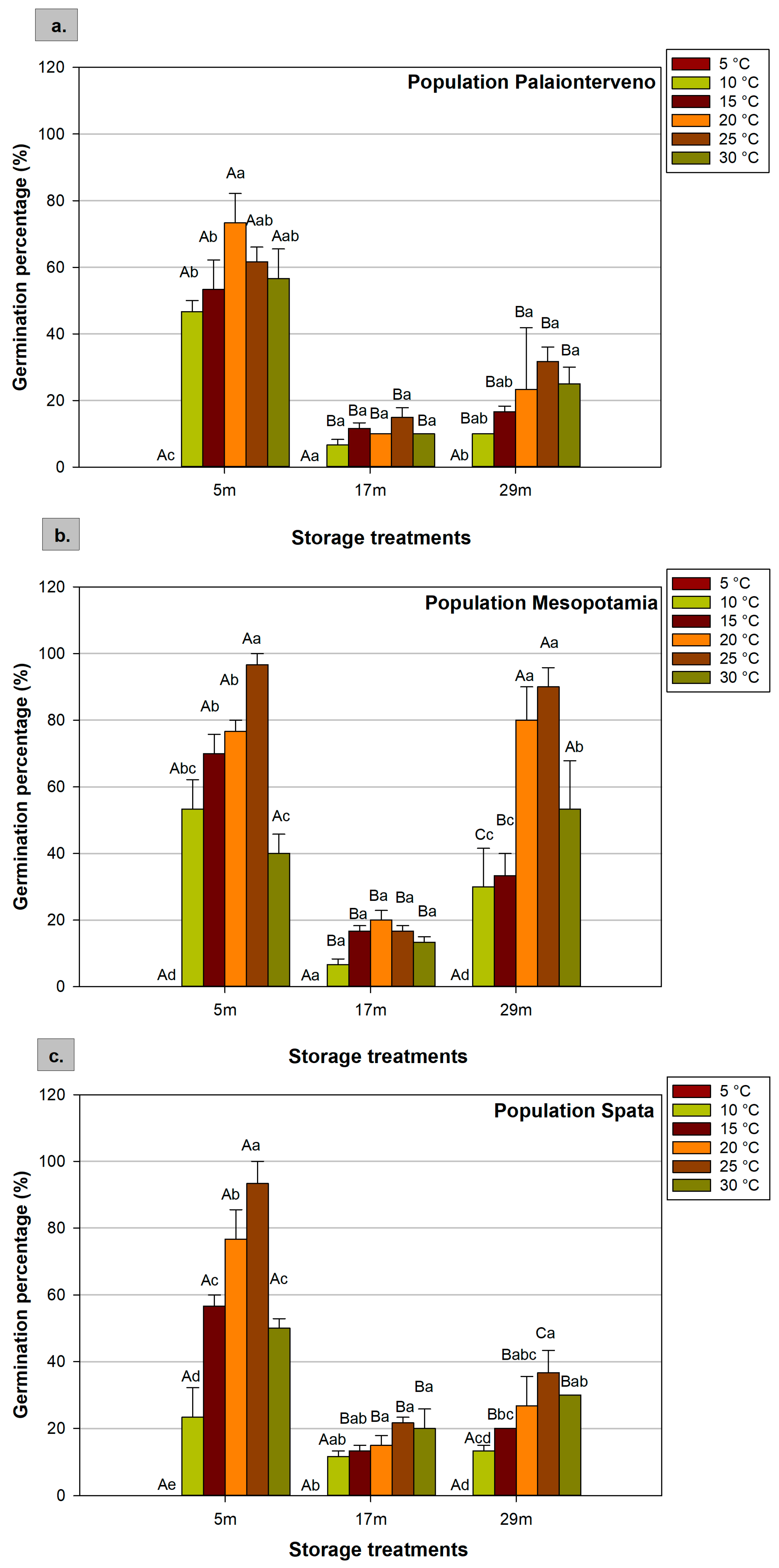
2.1. Germination Percentage
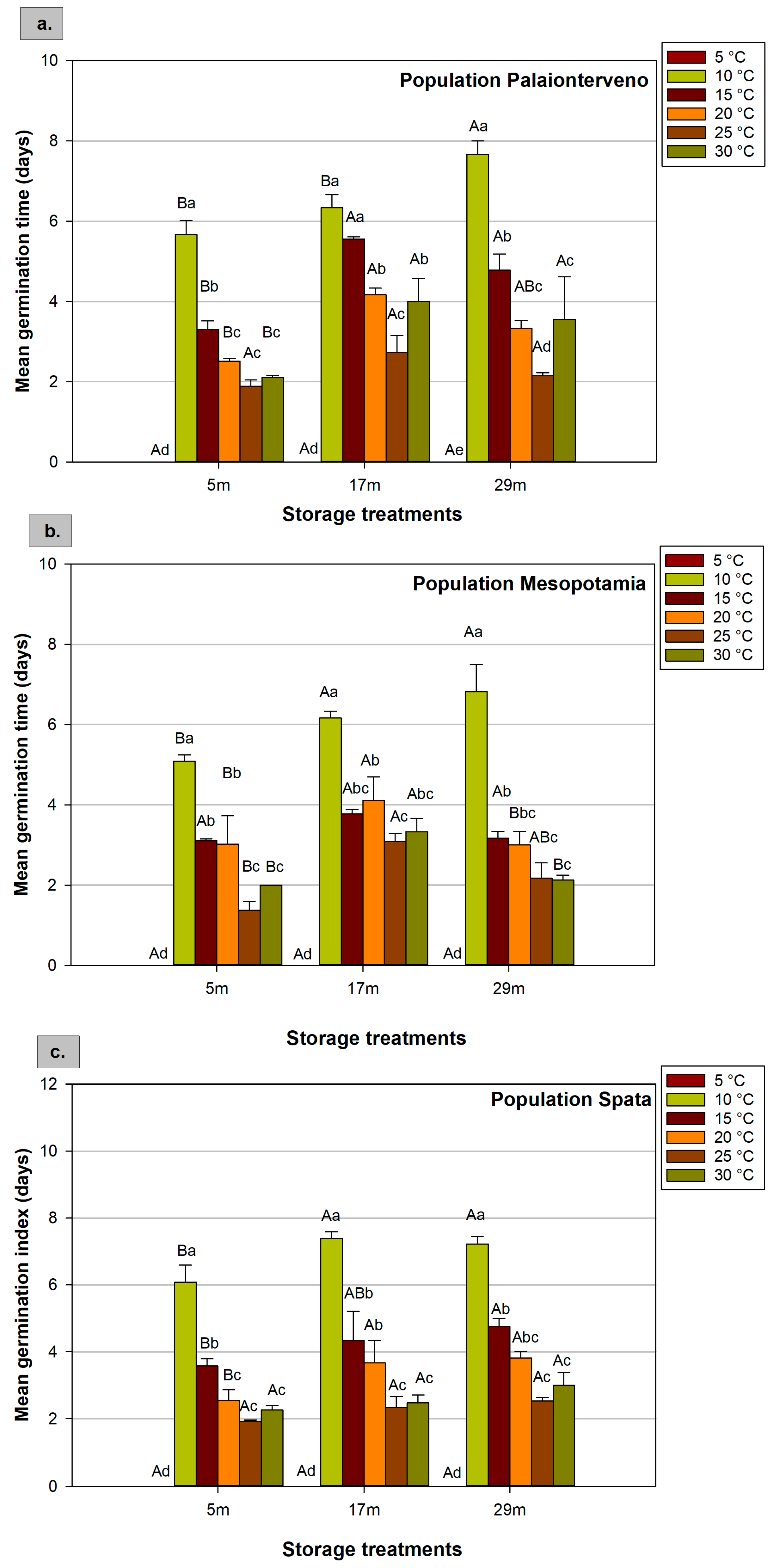
2.2. Mean Germination Time
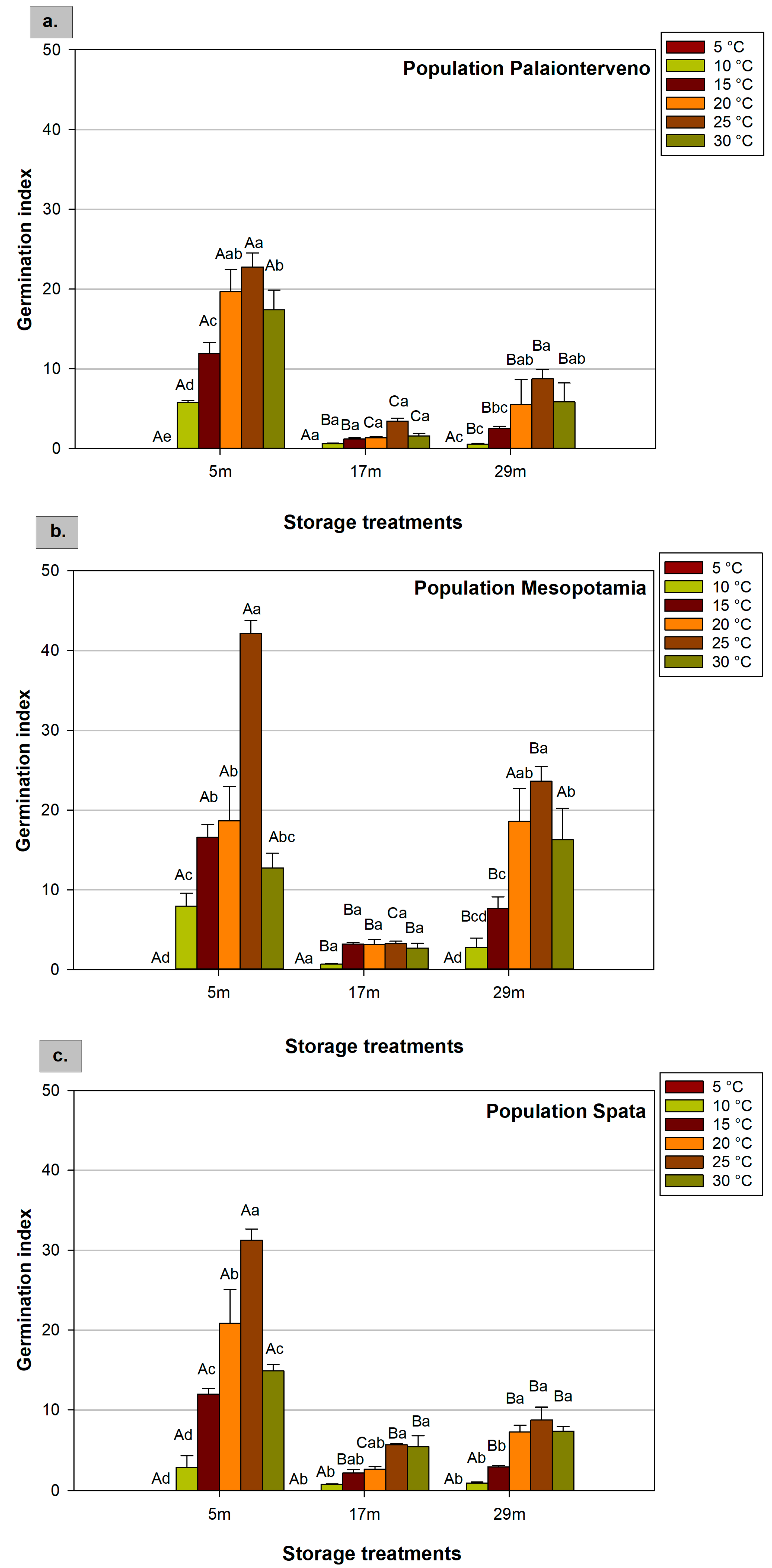
2.3. Germination Index
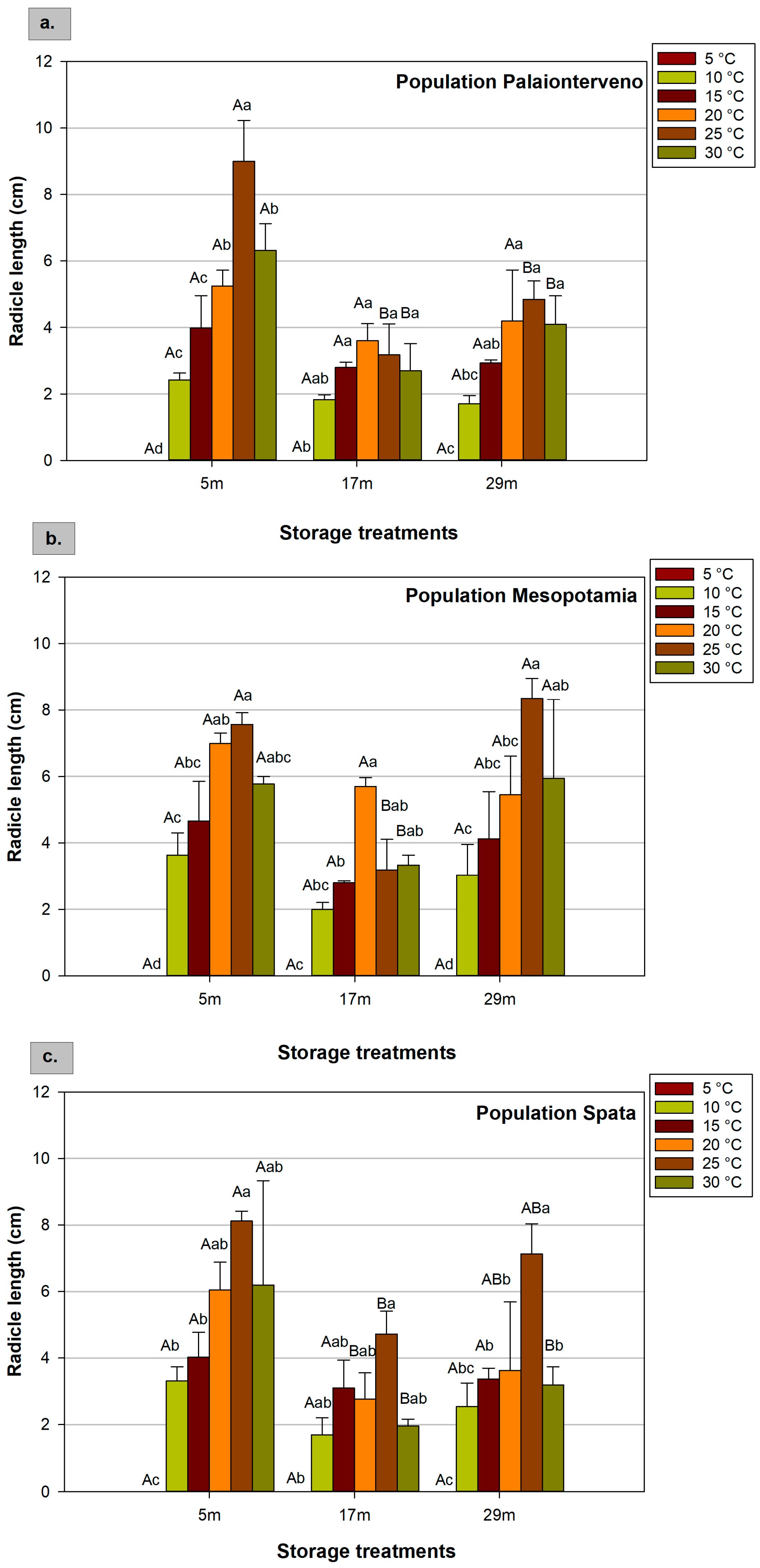
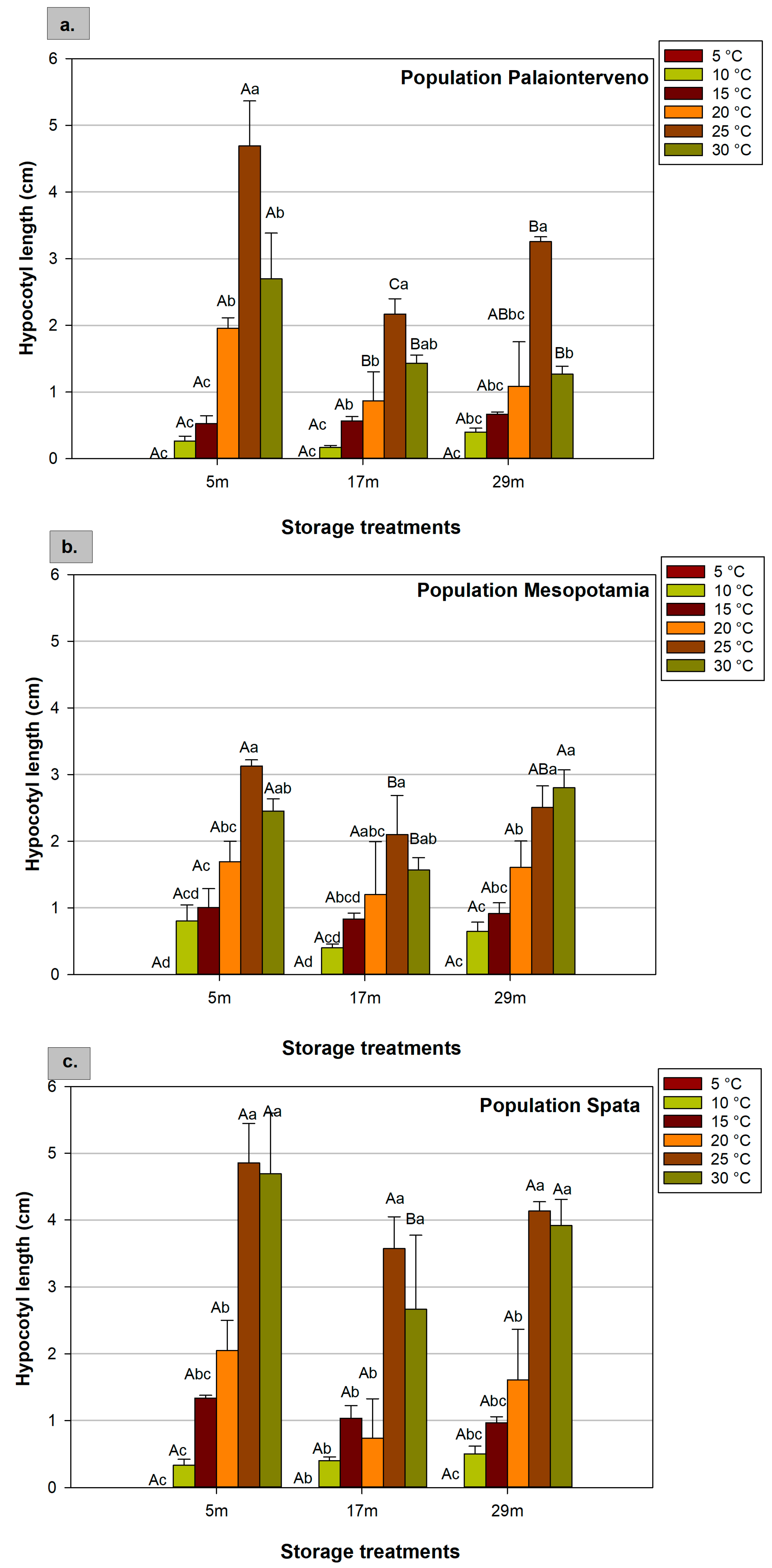
2.4. Radicle and Hypocotyl Length
3. Discussion
4. Materials and Methods
4.1. Seed Sampling, Storage, Germination Test, and Temperature Treatments
4.2. Measurements
4.2.1. Germination Percentage
4.2.2. Mean Germination Time
4.2.3. Germination Index
4.2.4. Radicle and Hypocotyl Length
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fathi-Achachlouei, B.; Azadmard-Damirchi, S.; Zahedi, Y.; Shaddel, R. Microwave Pretreatment as a Promising Strategy for Increment of Nutraceutical Content and Extraction Yield of Oil from Milk Thistle Seed. Ind. Crops Prod. 2019, 128, 527–533. [Google Scholar] [CrossRef]
- Rasic, S.; Stefanic, E.; Stefanic, I. Plants for Bees Silybum marianum. J. Apic. Res. 2009, 48, 298–299. [Google Scholar] [CrossRef]
- Karkanis, A.; Angou, A.; Athanasiadou, D.; Giannoulis, K.D.; Askianaki, R.; Kousi, N.; Sarridis, A.; Souipas, S.; Karamoutis, C. Using Post-emergence Herbicides in Combination with the Sowing Date to Suppress Sinapis arvensis and Silybum marianum in Durum Wheat. Agronomy 2022, 12, 2583. [Google Scholar] [CrossRef]
- Liava, V.; Karkanis, A.; Danalatos, N.; Tsiropoulos, N. Effects of Two Varieties and Fertilization Regimes on Growth, Fruit, and Silymarin Yield of Milk Thistle Crop. Agronomy 2022, 12, 105. [Google Scholar] [CrossRef]
- Montemurro, P.; Fracchiolla, M.; Lonigro, A. Effects of Some Environmental Factors on Seed Germination and Spreading Potentials of Silybum marianum Gaertner. Ital. J. Agron. 2007, 23, 315–320. [Google Scholar] [CrossRef]
- Ahmad, I.; Gul, B.; Khan, H.; Fawad, M. Influence of Various Densities of Wild Oat and Holy Thistle on the Productivity of Wheat (Triticum aestivum). Gesunde Pflanz. 2022, 74, 839–852. [Google Scholar] [CrossRef]
- Zhang, M.; Chen, K.; Hu, Z.; Shen, Q.; Wang, H. PRiME Pass-through Purification of Lignans in Silybum marianum and UPLC−MS/MS Analysis. J. Chromatogr. B 2018, 1093–1094, 128–133. [Google Scholar] [CrossRef]
- Ferraz, A.C.; Almeida, L.T.; da Silva Caetano, C.C.; da Silva Menegatto, M.B.; Souza Lima, R.L.; de Senna, J.P.N.; de Oliveira Cardoso, J.M.; Perucci, L.O.; Talvani, A.; Geraldo de Lima, W.; et al. Hepatoprotective, Antioxidant, Anti-inflammatory, and Antiviral Activities of Silymarin Against Mayaro Virus Infection. Antivir. Res. 2021, 194, 105168. [Google Scholar] [CrossRef]
- Mengesha, T.; Gnanasekaran, N.; Mehare, T. Hepatoprotective Effect of Silymarin on Fructose Induced Nonalcoholic Fatty Liver Disease in Male Albino Wistar Rats. BMC Complement. Med. Ther. 2021, 21, 104. [Google Scholar] [CrossRef]
- Mirjalili, M.; Mirzaei, E.; Vazin, A. Pharmacological Agents for the Prevention of Colistin-induced Nephrotoxicity. Eur. J. Med. Res. 2022, 27, 64. [Google Scholar] [CrossRef]
- Ralli, T.; Tripathi, T.; Kalaiselvan, V.; Tiwari, R.; Aeri, V.; Kohli, K. Silymarin as a Phyto-pharmaceutical: Isolation, Simultaneous Quantification of four Biomarkers and in-silico Anti-inflammatory Activity. Chin. J. Anal. Chem. 2023, 51, 100174. [Google Scholar] [CrossRef]
- Zutic, I.; Dudai, N. Factor Affecting Germination of Dalmatian Sage (Salvia officinalis). Acta Hortic. 2008, 782, 121–126. [Google Scholar] [CrossRef]
- Carrera-Castaño, G.; Calleja-Cabrera, J.; Pernas, M.; Gómez, L.; Oñate-Sánchez, L. An Updated Overview on the Regulation of Seed Germination. Plants 2020, 9, 703. [Google Scholar] [CrossRef]
- Buijs, G.A. Perspective on Secondary Seed Dormancy in Arabidopsis thaliana. Plants 2020, 9, 749. [Google Scholar] [CrossRef] [PubMed]
- Yan, A.; Chen, Z. The Control of Seed Dormancy and Germination by Temperature, Light and Nitrate. Bot. Rev. 2020, 86, 39–75. [Google Scholar] [CrossRef]
- Monemizadeh, Z.; Ghaderi-Far, F.; Sadeghipour, H.R.; Siahmarguee, A.; Soltani, E.; Torabi, B.; Baskin, C.C. Variation in Seed Dormancy and Germination among Populations of Silybum marianum (Asteraceae). Plant Species Biol. 2021, 36, 421–424. [Google Scholar] [CrossRef]
- Bhatt, A.; Caron, M.M.; Chen, X.; Yu, D.; Niu, Y. Effect of Temperature, Light and Storage on Seed Germination of Salvia plebeia R.Br., Leonurus japonicus Houtt., Mosla scabra (Thunb.) C.Y.Wu & H.W.Li and Perilla frutescens (L.) Britton. J. Appl. Res. Med. Aromat. Plants 2022, 31, 100402. [Google Scholar] [CrossRef]
- Malek, M.; Ghaderi-Far, F.; Torabi, B.; Sadeghipour, H.R. Dynamics of Seed Dormancy and Germination at High Temperature Stress is Affected by Priming and Phytohormones in Rapeseed (Brassica napus L.). J. Plant Physiol. 2022, 269, 153614. [Google Scholar] [CrossRef]
- Parmoon, G.; Moosavi, S.A.; Akbari, H.; Ebadi, A. Quantifying Cardinal Temperatures and Thermal Time Required for Germination of Silybum marianum seed. Crop. J. 2015, 3, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Sohrabi, S.; Ghanbari, A.; Mohassel, M.; Gherekhloo, J.; Vidal, R. Effects of Environmental Factors on Cucumis melo L. subsp. agrestis var. agrestis (Naudin) Pangalo Seed Germination and Seedling Emergence. S. Afr. J. Bot. 2016, 105, 1–8. [Google Scholar] [CrossRef]
- Rezvani, M.; Nadimi, S.; Zaefarian, F.; Chauhan, B. Environmental Factors Affecting Seed Germination and Seedling Emergence of three Phalaris Species. Crop. Prot. 2021, 148, 105743. [Google Scholar] [CrossRef]
- Dowsett, C.A.; Buddenhagen, C.; James, T.K.; McGill, C.R. Yellow Bristle Grass (Setaria pumila) Germination Biology. N. Z. Plant Prot. 2018, 71, 72–80. Available online: https://journal.nzpps.org/index.php/nzpp/article/view/154 (accessed on 18 January 2023). [CrossRef]
- Khan, A.M.; Mobli, A.; Werth, J.A.; Chauhan, B.S. Germination and Seed Persistence of Amaranthus retroflexus and Amaranthus viridis: Two Emerging Weeds in Australian Cotton and Other Summer Crops. PLoS ONE 2022, 17, e0263798. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, J.R.; Debaeke, P.; Steinberg, C.; You, M.P.; Barbetti, M.J.; Aubertot, J. Abiotic and Biotic Factors Affecting Crop Seed Germination and Seedling Emergence: A Conceptual Framework. Plant Soil 2018, 432, 1–28. [Google Scholar] [CrossRef]
- Kibar, H.; Yücesan, B. Effects of Storage Durations at Different Temperatures on Various Physiological Parameters of Einkorn Seed (Triticum monucocum L.) Germination. J. Stored Prod. Res. 2021, 93, 101851. [Google Scholar] [CrossRef]
- Koskosidis, A.; Khah, E.M.; Pavli, O.I.; Vlachostergios, D.N. Effect of Storage Conditions on Seed Quality of Soybean (Glycine max L.) Germplasm. AIMS Agric. Food 2022, 7, 387–402. [Google Scholar] [CrossRef]
- Islam, M.M.; Rengel, Z.; Storer, P.; Siddique, K.H.M.; Solaiman, Z.M. Industrial Hemp (Cannabis sativa L.) Varieties and Seed Pre-treatments Affect Seed Germination and Early Growth of Seedlings. Agronomy 2022, 12, 6. [Google Scholar] [CrossRef]
- Li, R.; Chen, L.; Wu, Y.; Zhang, R.; Baskin, C.C.; Baskin, J.M.; Hu, X. Effects of Cultivar and Maternal Environment on Seed Quality in Vicia sativa. Front. Plant Sci. 2017, 8, 1411. [Google Scholar] [CrossRef] [Green Version]
- Roman, A.M.; Truta, A.M.; Viman, O.; Morar, I.M.; Spalevic, V.; Dan, C.; Sestras, R.E.; Holonec, L.; Sestras, A.F. Seed Germination and Seedling Growth of Robinia pseudoacacia Depending on the Origin of Different Geographic Provenances. Diversity 2022, 14, 34. [Google Scholar] [CrossRef]
- Kissing Kucek, L.; Azevedo, M.D.; Eagen, S.S.; Ehlke, N.J.; Hayes, R.J.; Mirsky, S.B.; Reberg-Horton, C.; Ryan, M.R.; Wayman, S.; Wiering, N.P.; et al. Seed Dormancy in Hairy Vetch (Vicia villosa Roth) Is Influenced by Genotype and Environment. Agronomy 2020, 10, 1804. [Google Scholar] [CrossRef]
- Fernández, R.; Chantre, G.R.; Renzi, J.P. Seed dormancy of Lolium perenne L. Related to the Maternal Environment During Seed Filling. Seed Sci. Res. 2021, 31, 217–223. [Google Scholar] [CrossRef]
- Murthy, U.M.N.; Kumar, P.P.; Sun, W.Q. Mechanisms of seed ageing under different storage conditions for Vigna radiata (L.) Wilczek: Lipid peroxidation, sugar hydrolysis, Maillard reactions and their relationship to glass state transition. J. Exp. Bot. 2003, 54, 1057–1067. [Google Scholar] [CrossRef] [PubMed]
- Kurek, K.; Plitta-Michalak, B.; Ratajczak, E. Reactive Oxygen Species as Potential Drivers of the Seed Aging Process. Plants 2019, 8, 174. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Niu, Y.; Zheng, Y.; Wang, Z. Advances in the Understanding of Reactive Oxygen Species-Dependent Regulation on Seed Dormancy, Germination, and Deterioration in Crops. Front. Plant Sci. 2022, 13, 826809. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, R.; Singh, S.T. Seed Aging, Storage and Deterioration: An Irresistible Physiological Phenomenon. Agric. Rev. 2019, 40, 234–238. [Google Scholar] [CrossRef] [Green Version]
- Padilha, M.S.; Coelho, C.M.M.; Ehrhardt-Brocardo, N.C.M. Viability and Performance of Wheat Seedlings after Artificial Seed Aging. J. Seed Sci. 2022, 44, e202244037. [Google Scholar] [CrossRef]
- Cokkizgin, A. Salinity Stress in Common Bean (Phaseolus vulgaris L.) Seed Germination. Not. Bot. Horti Agrobot. 2012, 40, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Boakye Boadu, K.; Siaw, D. The Effect of Storage Period on the Viability of Triplochiton scleroxylon K. Schum. Seeds. Sci. Afr. 2019, 5, e00127. [Google Scholar] [CrossRef]
- Martin, R.J.; Deo, B.; Douglas, J.A. Effect of Time of Sowing on Reproductive Development of Variegated Thistle. Agron N. Z. 2000, 30, 1–5. [Google Scholar]
- Bürger, J.; Malyshev, A.V.; Colbach, N. Populations of Arable Weed Species Show Intra-Specific Variability in Germination Base Temperature but not in Early Growth Rate. PLoS ONE 2020, 15, e0240538. [Google Scholar] [CrossRef]
- Guillemin, J.; Gardarin, A.; Granger, S.; Reibel, C.; Munier-Jolain, N.; Colbach, N. Assessing Potential Germination Period of Weeds with Base Temperatures and Base Water Potentials. Weed Res. 2012, 53, 76–87. [Google Scholar] [CrossRef]
- De Roo, A.; Tozzi, E.; Benaragama, D.I.; Willenborg, C.J. Recruitment Biology of Cleavers (Galium spp.) Populations in Western Canada. Weed Sci. 2022, 70, 669–679. [Google Scholar] [CrossRef]
- Koocheki, A.; Nassiri, M.; Alimoradi, L.; Ghorbani, R. Effect of Cropping Systems and Crop Rotations on Weeds. Agron. Sustain. Dev. 2009, 29, 401–408. [Google Scholar] [CrossRef] [Green Version]
- Manalil, S.; Chauhan, B.S. Seedbank Persistence and Emergence Pattern of Argemone mexicana, Rapistrum rugosum and Sonchus oleraceus in the Eastern Grain Region of Australia. Sci. Rep. 2021, 11, 18095. [Google Scholar] [CrossRef]
- Long, R.L.; Gorecki, M.J.; Renton, M.; Scott, J.K.; Colville, L.; Goggin, D.E.; Commander, L.E.; Westcott, D.A.; Cherry, H.; Finch-Savage, W.E. The Ecophysiology of Seed Persistence: A Mechanistic View of the Journey to Germination or Demise. Biol. Rev. 2014, 90, 31–59. [Google Scholar] [CrossRef]
- Týr, Š. Weed Infestation in the Stand of Milk Thistle and Infestation in Sustainable Crop Rotation. Acta Fytotech. Zootech. 2015, 18, 90–94. [Google Scholar] [CrossRef] [Green Version]
- Arampatzis, D.; Karkanis, A.; Tsiropoulos, N. Silymarin Content and Antioxidant Activity of Seeds of Wild Silybum marianum populations growing in Greece. Ann. Appl. Biol. 2019, 174, 61–73. [Google Scholar] [CrossRef] [Green Version]
- Arampatzis, D.A.; Karkanis, A.C.; Tsiropoulos, N.G. Impact of Plant Density and Mepiquat Chloride on Growth, Yield, and Silymarin Content of Silybum marianum Grown under Mediterranean Semi-Arid Conditions. Agronomy 2019, 9, 669. [Google Scholar] [CrossRef] [Green Version]
- Martinelli, T.; Andrzejewska, J.; Salis, M.; Sulas, L. Phenological Growth Stages of Silybum marianum According to the Extended BBCH Scale. Ann. Appl. Biol. 2015, 166, 53–66. [Google Scholar] [CrossRef]
- Roman, A.M.; Truta, A.M.; Morar, I.M.; Viman, O.; Dan, C.; Sestras, A.F.; Holonec, L.; Boscaiu, M.; Sestras, R.E. From Seed to Seedling: Influence of Seed Geographic Provenance and Germination Treatments on Reproductive Material Represented by Seedlings of Robinia pseudoacacia. Sustainability 2022, 14, 5654. [Google Scholar] [CrossRef]
- Deng, Z.; Song, S. Sodium Nitroprusside, Ferricyanide, Nitrite and Nitrate Decrease the Thermo-dormancy of Lettuce Seed Germination in a Nitric Oxide-dependent Manner in Light. S. Afr. J. Bot. 2012, 78, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Włodarczyk, K.; Smolińska, B. The Effect of Nano-ZnO on Seeds Germination Parameters of Different Tomatoes (Solanum lycopersicum L.) Cultivars. Molecules 2022, 27, 4963. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.H.; Roberts, E.H. The Quantification of Ageing and Survival in Orthodox Seeds. Seed Sci. Technol. 1981, 9, 373–409. [Google Scholar]
- AOSA (Association of Official Seed Analysts). Seed Vigor Hand Testing Book: Contribution No. 32 to the Handbook on Seed Testing; The Association: Ithaca, NY, USA, 1983. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | F Values of ANOVA | ||||
|---|---|---|---|---|---|
| GP | MGT | GI | RL | HL | |
| Population (P) | 27.848 *** | 7.007 *** | 30.562 *** | 3.808 * | 13.826 *** |
| Storage (S) | 220.704 *** | 38.445 *** | 273.201 *** | 25.902 *** | 16.958 *** |
| Temperature (T) | 90.281 *** | 351.308 *** | 128.943 *** | 54.827 *** | 113.374 *** |
| P × S | 12.784 *** | 2.504 * | 10.566 *** | 0.987 ns | 0.768 ns |
| P × T | 3.728 *** | 2.366 * | 5.054 *** | 0.831 ns | 5.300 *** |
| S × T | 14.236 *** | 2.964 ** | 30.257 *** | 2.873 ** | 2.782 ** |
| P × S × T | 2.937 *** | 1.306 ns | 3.810 *** | 0.619 ns | 0.696 ns |
| Temperature | GP (%) | MGT (Days) | GI | RL (cm) | HL (cm) |
|---|---|---|---|---|---|
| 5 °C | 0 d | 0 e | 0 e | 0 d | 0 e |
| 10 °C | 41.1 c | 5.61 a | 5.53 d | 3.12 c | 0.47 de |
| 15 °C | 60.0 b | 3.33 b | 13.49 c | 4.22 bc | 0.96 d |
| 20 °C | 75.6 a | 2.69 bc | 19.74 b | 6.09 ab | 1.90 c |
| 25 °C | 83.9 a | 1.73 d | 32.07 a | 8.23 a | 4.23 a |
| 30 °C | 48.9 bc | 2.12 cd | 15.00 bc | 6.10 ab | 3.28 b |
| Populations | GP (%) | MGT (Days) | GI | RL (cm) | HL (cm) |
|---|---|---|---|---|---|
| Palaionterveno | 73.3 a | 2.51 a | 19.70 a | 5.24 a | 1.96 a |
| Mesopotamia | 76.7 a | 3.02 a | 18.66 a | 6.99 a | 1.69 a |
| Spata | 76.7 a | 2.84 a | 20.87 a | 6.05 a | 2.05 a |
| Storage Treatments | GP (%) | MGT (Days) | GI | RL (cm) | HL (cm) |
|---|---|---|---|---|---|
| 5 m | 75.6 a | 2.69 a | 19.74 a | 6.09 a | 1.90 a |
| 17 m | 15.0 c | 3.98 a | 2.39 c | 4.02 a | 0.93 a |
| 29 m | 43.3 b | 3.38 a | 10.47 b | 4.42 a | 1.43 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liava, V.; Ntatsi, G.; Karkanis, A. Seed Germination of Three Milk Thistle (Silybum marianum (L.) Gaertn.) Populations of Greek Origin: Temperature, Duration, and Storage Conditions Effects. Plants 2023, 12, 1025. https://doi.org/10.3390/plants12051025
Liava V, Ntatsi G, Karkanis A. Seed Germination of Three Milk Thistle (Silybum marianum (L.) Gaertn.) Populations of Greek Origin: Temperature, Duration, and Storage Conditions Effects. Plants. 2023; 12(5):1025. https://doi.org/10.3390/plants12051025
Chicago/Turabian StyleLiava, Vasiliki, Georgia Ntatsi, and Anestis Karkanis. 2023. "Seed Germination of Three Milk Thistle (Silybum marianum (L.) Gaertn.) Populations of Greek Origin: Temperature, Duration, and Storage Conditions Effects" Plants 12, no. 5: 1025. https://doi.org/10.3390/plants12051025
APA StyleLiava, V., Ntatsi, G., & Karkanis, A. (2023). Seed Germination of Three Milk Thistle (Silybum marianum (L.) Gaertn.) Populations of Greek Origin: Temperature, Duration, and Storage Conditions Effects. Plants, 12(5), 1025. https://doi.org/10.3390/plants12051025