


Effects of Coumarin on Rhizosphere Microbiome and Metabolome of Lolium multiflorum


Abstract
:1. Introduction
2. Results
2.1. Diversity in the Bacterial Community of the Rhizosphere
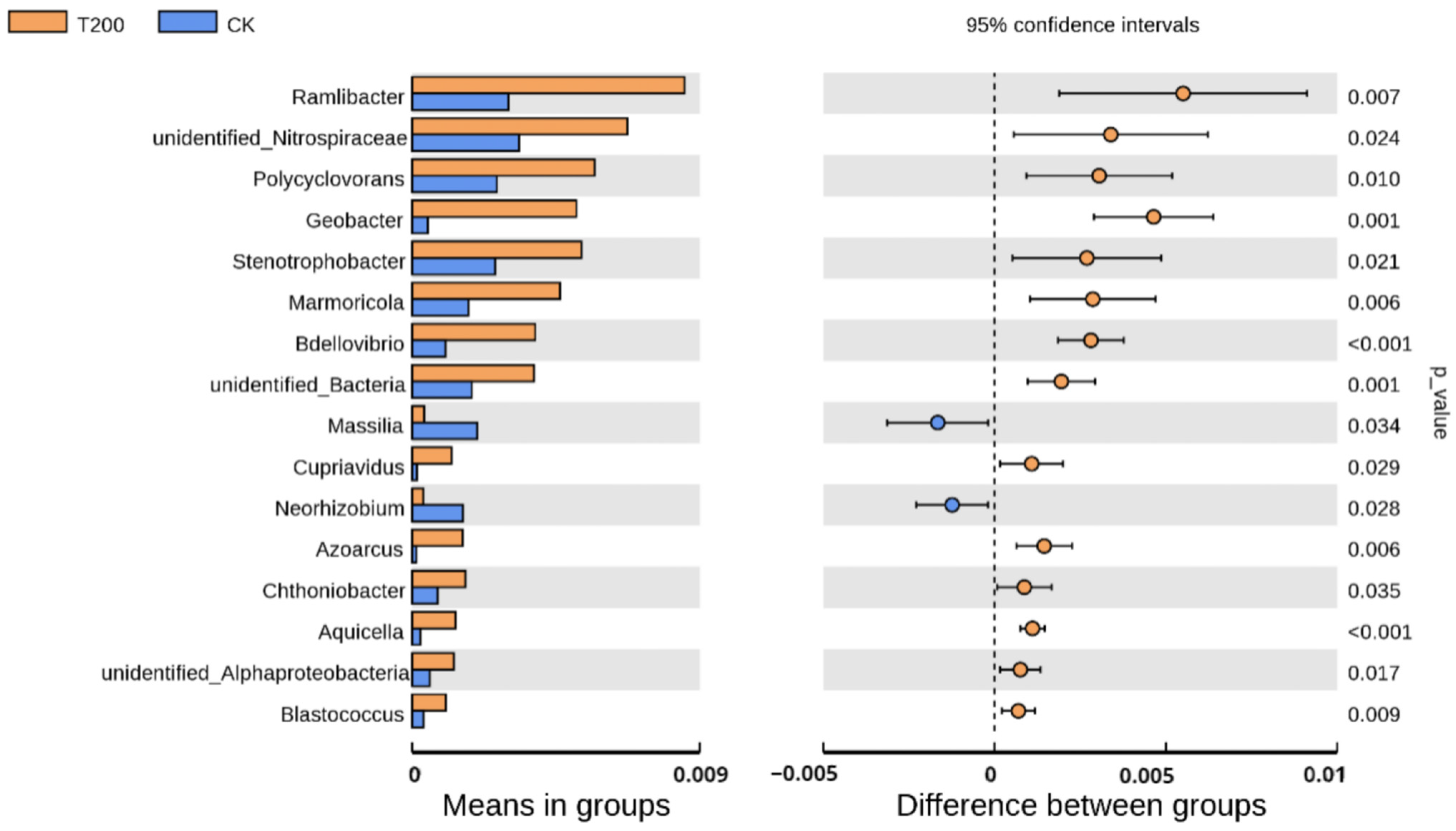
2.2. Comparison of Relative Abundance of Bacterial Communities in Rhizosphere Soil under Phylum and Genus Classification
2.3. Non-Targeted Metabolomics Analysis of Roots Using LC-MS
2.4. Differential Metabolite Analysis in Roots through Non-Targeted Metabolomics
2.5. KEGG Pathway Enrichment Analysis of Differentially Expressed Metabolites in Roots
2.6. Correlation Analysis between Bacterial Genera in the Rhizosphere and Root Metabolites
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Materials and Plant Treatments
5.2. Sampling of Roots and Rhizosphere Soil of Annual Ryegrass
5.3. DNA Extraction and Sequencing
5.4. Sequencing Data Analysis
5.5. Root Metabolites Extraction and LC-MS Analysis
5.6. Metabolomics Data Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Borges, F.; Roleira, F.; Milhazes, N.; Santana, L.; Uriarte, E. Simple coumarins and analogues in medicinal chemistry: Occurrence, synthesis and biological activity. Curr. Med. Chem. 2005, 12, 887–916. [Google Scholar] [CrossRef] [PubMed]
- Hadaček, F.; Müller, C.; Werner, A.; Greger, H.; Proksch, P. Analysis, isolation and insecticidal activity of linear furanocoumarins and other coumarin derivatives from Peucedanum (Apiaceae: Apioideae). J. Chem. Ecol. 1994, 20, 2035–2054. [Google Scholar] [CrossRef] [PubMed]
- Ganzera, M.; Sturm, S.; Stuppner, H. HPLC-MS and MECC analysis of coumarins. Chromatographia 1997, 46, 197–203. [Google Scholar] [CrossRef]
- Hussain, M.I.; Syed, Q.A.; Khattak, M.N.K.; Hafez, B.; Reigosa, M.J.; El-Keblawy, A. Natural product coumarins: Biological and pharmacological perspectives. Biologia 2019, 74, 863–888. [Google Scholar] [CrossRef]
- Stassen, M.J.J.; Hsu, S.-H.; Pieterse, C.M.J.; Stringlis, I.A. Coumarin communication along the microbiome–root–shoot axis. Trends Plant Sci. 2021, 26, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Razavi, S.M.; Zarrini, G. Bioactivity of aviprin and aviprin-3″-O-glucoside, two linear furanocoumarins from Apiaceae. Russ. J. Bioorganic Chem. 2010, 36, 359–362. [Google Scholar] [CrossRef]
- Misra, B.B.; Das, V.; Landi, M.; Abenavoli, M.; Araniti, F. Short-term effects of the allelochemical umbelliferone on Triticum durum L. metabolism through GC–MS based untargeted metabolomics. Plant Sci. 2020, 298, 110548. [Google Scholar] [CrossRef]
- Chuah, T.S.; Tan, P.K.; Ismail, B.S. Effects of adjuvants and soil microbes on the phytotoxic activity of coumarin in combination with p-vanillin on goosegrass (Eleusine indica L.) seedling emergence and growth. S. Afr. J. Bot. 2013, 84, 128–133. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.X.; Zhao, G.Q.; Liu, S.J. Isolation and identification of potential allelochemicals in water extract of yellow sweetclover (Melilotus officinalis). Allelopath. J. 2015, 36, 75–86. [Google Scholar]
- Wu, C.-X.; Zhao, G.-Q.; Liu, D.-L.; Liu, S.-J.; Gun, X.-X.; Tang, Q. Discovery and Weed Inhibition Effects of Coumarin as the Predominant Allelochemical of Yellow Sweetclover (Melilotus officinalis). Int. J. Agric. Biol. 2016, 18, 168–175. [Google Scholar] [CrossRef]
- Abenavoli, M.R.; Sorgonà, A.; Albano, S.; Cacco, G. Coumarin differentially affects the morphology of different root types of maize seedlings. J. Chem. Ecol. 2004, 30, 1871–1883. [Google Scholar] [CrossRef]
- Abenavoli, M.R.; Sorgonà, A.; Sidari, M.; Badiani, M.; Fuggi, A. Coumarin inhibits the growth of carrot (Daucus carota L. cv. Saint Valery) cells in suspension culture. J. Plant Physiol. 2003, 160, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Abenavoli, M.R.; Sorgona, A.; Muscolo, A. Morphological changes in tissue culture of Petunia hybrida in response to coumarin allelochemical. Allelopath. J. 2001, 8, 171–177. [Google Scholar]
- Masuda, T.; Takasugi, M.; Anetai, M. Psoralen and other linear furanocoumarins as phytoalexins in Glehnia littoralis. Phytochemistry 1998, 47, 13–16. [Google Scholar] [CrossRef]
- Dutsadee, C.; Nunta, C. Induction of peroxidase, scopoletin, phenolic compounds and resistance in Hevea brasiliensis by elicitin and a novel protein elicitor purified from Phytophthora palmivora. Physiol. Mol. Plant Pathol. 2008, 72, 179–187. [Google Scholar] [CrossRef]
- Zeid, A.H.S.A. Stress metabolites from Corchorus olitorius L. leaves in response to certain stress agents. Food Chem. 2002, 76, 187–195. [Google Scholar] [CrossRef]
- Valle, T.; López-Pérez, J.L.; Hernández, J.M.; Corchete, P. Antifungal activity of scopoletin and its differential accumulation in Ulmus pumila and Ulmus campestris cell suspension cultures infected with Ophiostoma ulmi spores. Plant Sci. 1997, 125, 97–101. [Google Scholar] [CrossRef]
- El Oirdi, M.; Trapani, A.; Bouarab, K. The nature of tobacco resistance against Botrytis cinerea depends on the infection structures of the pathogen. Environ. Microbiol. 2010, 12, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Wang, L.; Zhang, B.; Ma, J.; Hettenhausen, C.; Cao, G.; Sun, G.; Wu, J.; Wu, J. Scopoletin is a phytoalexin against Alternaria alternata in wild tobacco dependent on jasmonate signalling. J. Exp. Bot. 2014, 65, 4305–4315. [Google Scholar] [CrossRef]
- Lipeeva, A.V.; Zakharov, D.O.; Burova, L.G.; Frolova, T.S.; Baev, D.S.; Shirokikh, I.V.; Evstropov, A.N.; Sinitsyna, O.I.; Tolsikova, T.G.; Shults, E.E. Design, Synthesis and Antibacterial Activity of Coumarin-1,2,3-triazole Hybrids Obtained from Natural Furocoumarin Peucedanin. Molecules 2019, 24, 2126. [Google Scholar] [CrossRef] [Green Version]
- Kwon, Y.S.; Kobayashi, A.; Kajiyama, S.I.; Kawazu, K.; Kanzaki, H.; Kim, C.M. Antimicrobial constituents of Angelica dahurica roots. Phytochemistry 1997, 44, 887–889. [Google Scholar] [CrossRef]
- Al-Barwani, F.M.; Eltayeb, E.A. Antifungal compounds from induced Conium maculatum L. plants. Biochem. Syst. Ecol. 2004, 32, 1097–1108. [Google Scholar] [CrossRef]
- Stringlis, I.A.; Yu, K.; Feussner, K.; de Jonge, R.; Van Bentum, S.; Van Verk, M.C.; Berendsen, R.L.; Bakker, P.A.H.M.; Feussner, I.; Pieterse, C.M.J. MYB72-dependent coumarin exudation shapes root microbiome assembly to promote plant health. Proc. Natl. Acad. Sci. USA 2018, 115, E5213–E5222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zubair, H.M.; Pratley, J.; Sandral, G.A.; Humphries, A. Allelopathic interference of alfalfa (Medicago sativa L.) genotypes to annual ryegrass (Lolium rigidum). J. Plant Res. 2017, 130, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Yao, D.D.; Wang, J.Y.; Zhou, Q.; Tang, Q.; Zhao, G.Q.; Wu, C.X. Effect of coumarin on Italian ryegrass seed germination and seedling growth. Acta Pratacul. Sin. 2017, 26, 136–145. [Google Scholar] [CrossRef]
- Lemanceau, P.; Barret, M.; Mazurier, S.; Mondy, S.; Pivato, B.; Fort, T.; Vacher, C. Plant Communication with Associated Microbiota in the Spermosphere, Rhizosphere and Phyllosphere. In How Plants Communicate with Their Biotic Environment; Becard, G., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; Volume 82, pp. 101–133. [Google Scholar]
- Sasse, J.; Martinoia, E.; Northen, T. Feed Your Friends: Do Plant Exudates Shape the Root Microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinsinger, P.; Bengough, A.G.; Vetterlein, D.; Young, I.M. Rhizosphere: Biophysics, biogeochemistry and ecological relevance. Plant Soil 2009, 321, 117–152. [Google Scholar] [CrossRef]
- Haney, C.; Samuel, B.; Bush, J.; Ausubel, F.M. Associations with rhizosphere bacteria can confer an adaptive advantage to plants. Nat. Plants 2015, 1, 15051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, T.; Ke, M.; Lavoie, M.; Jin, Y.; Fan, X.; Zhang, Z.; Fu, Z.; Sun, L.; Gillings, M.; Peñuelas, J.; et al. Rhizosphere microorganisms can influence the timing of plant flowering. Microbiome 2018, 6, 2738. [Google Scholar] [CrossRef] [Green Version]
- Cook, R.J.; Thomashow, L.S.; Weller, D.M.; Fujimoto, D.; Mazzola, M.; Bangera, G.; Kim, D.S. Molecular mechanisms of defense by rhizobacteria against root disease. Proc. Natl. Acad. Sci. USA 1995, 92, 4197–4201. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Verma, J.P. The Role of Microbes to Improve Crop Productivity and Soil Health. In Ecological Wisdom Inspired Restoration Engineering; Achal, V., Mukherjee, A., Eds.; Springer: Singapore, 2019; pp. 249–265. [Google Scholar]
- Bharti, V.S.; Dotaniya, M.L.; Shukla, S.P.; Yadav, V.K. Managing Soil Fertility Through Microbes: Prospects, Challenges and Future Strategies. In Agro-Environmental Sustainability; Singh, J., Seneviratne, G., Eds.; Springer: Cham, Switzerland; New York, NY, USA, 2017; pp. 81–111. [Google Scholar]
- Qian, H.; Zhu, Y.; Chen, S.; Jin, Y.; Lavoie, M.; Ke, M.; Fu, Z. Interacting effect of diclofop-methyl on the rice rhizosphere microbiome and denitrification. Pestic. Biochem. Physiol. 2018, 146, 90–96. [Google Scholar] [CrossRef]
- Niro, E.; Marzaioli, R.; De Crescenzo, S.; D’Abrosca, B.; Castaldi, S.; Esposito, A.; Fiorentino, A.; Rutigliano, F. Effects of the allelochemical coumarin on plants and soil microbial community. Soil Biol. Biochem. 2016, 95, 30–39. [Google Scholar] [CrossRef]
- Cheng, Z.; Chen, Y.; Zhang, F. Effect of reclamation of abandoned salinized farmland on soil bacterial communities in arid northwest China. Sci. Total Environ. 2018, 630, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.K.; Zuo, X.A.; Zhao, X.Y.; Li, Y.Q.; Zhou, X.; Lv, P.; Luo, Y.Q.; Yun, J.Y. Responses of soil fungal community to the sandy grassland restoration in Horqin Sandy Land, northern China. Environ. Monit. Assess. 2016, 188, 21. [Google Scholar] [CrossRef] [PubMed]
- Cook, R.J. Making Greater Use of Introduced Microorganisms for Biological Control of Plant Pathogens. Annu. Rev. Phytopathol. 1993, 31, 53–80. [Google Scholar] [CrossRef] [PubMed]
- Johnston-Monje, D.; Lundberg, D.S.; Lazarovits, G.; Reis, V.M.; Raizada, M.N. Bacterial populations in juvenile maize rhizospheres originate from both seed and soil. Plant Soil 2016, 405, 337–355. [Google Scholar] [CrossRef] [Green Version]
- Nagel, K.; Schneemann, I.; Kajahn, I.; Labes, A.; Wiese, J.; Imhoff, J. Beneficial effects of 2,4-diacetylphloroglucinol- producing pseudomonads on the marine alga Saccharina latissima. Aquat. Microb. Ecol. 2012, 67, 239–249. [Google Scholar] [CrossRef] [Green Version]
- Schmid, N.B.; Giehl, R.F.; Döll, S.; Mock, H.-P.; Strehmel, N.; Scheel, D.; Kong, X.; Hider, R.C.; von Wirén, N. Feruloyl-CoA 6′-Hydroxylase1-Dependent Coumarins Mediate Iron Acquisition from Alkaline Substrates in Arabidopsis. Plant Physiol. 2014, 164, 160–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, H.; Günther, C.; Weber, M.; Spörlein, C.; Loscher, S.; Böttcher, C.; Schobert, R.; Clemens, S. Metabolome Analysis of Arabidopsis thaliana Roots Identifies a Key Metabolic Pathway for Iron Acquisition. PLoS ONE 2014, 9, e102444. [Google Scholar] [CrossRef] [Green Version]
- Rajniak, J.; Giehl, R.F.H.; Chang, E.; Murgia, I.; von Wirén, N.; Sattely, E.S. Biosynthesis of redox-active metabolites in response to iron deficiency in plants. Nat. Chem. Biol. 2018, 14, 442–450. [Google Scholar] [CrossRef]
- Wang, F.; Xv, Q.; Chen, Z.H.; Chen, Q.M.; Yang, Q.J.; Chen, A.L. Impact of burying different kinds of green manure on soil bacteria in growing tobacco field. Jiangsu Agric. Sci. 2019, 47, 317–321. [Google Scholar] [CrossRef]
- Chang, E.H.; Yang, J.C. Root exudates and their role in plant growth. Tillage Cultiv. 2006, 5, 13–16. [Google Scholar] [CrossRef]
- Xue, J.; Chen, T.T.; Zheng, J.W.; Balamurugan, S.; Cai, J.X.; Liu, Y.H.; Yang, W.D.; Liu, J.S.; Li, H.Y. The role of diatom glucose-6-phosphate dehydrogenase on lipogenic NADPH supply in green microalgae through plastidial oxidative pentose phosphate pathway. Appl. Microbiol. Biotechnol. 2018, 102, 10803–10815. [Google Scholar] [CrossRef]
- de Freitas-Silva, L.; Rodríguez-Ruiz, M.; Houmani, H.; da Silva, L.C.; Palma, J.M.; Corpas, F.J. Glyphosate-induced oxidative stress in Arabidopsis thaliana affecting peroxisomal metabolism and triggers activity in the oxidative phase of the pentose phosphate pathway (OxPPP) involved in NADPH generation. J. Plant Physiol. 2017, 218, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Xu, J.; Wang, J.; Yuan, Z.; Li, Y.; Wu, C. Effects of different concentrations of coumarin aqueous solution on ryegrass seedlings. Guangxi Plant Prot. 2019, 32, 5. [Google Scholar] [CrossRef]
- Andriotis, V.M.E.; Smith, A.M. The plastidial pentose phosphate pathway is essential for postglobular embryo development in Arabidopsis. Proc. Natl. Acad. Sci. USA 2019, 116, 15297–15306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erland, L.A.E.; Saxena, P. Auxin driven indoleamine biosynthesis and the role of tryptophan as an inductive signal in Hypericum perforatum (L.). PLoS ONE 2019, 14, e0223878. [Google Scholar] [CrossRef] [Green Version]
- El Karamany, M.F.; Omer, H.A.A.; Bakry, B.A.; Bakhoum, G.S.; Sadak, M.S. Impact of tryptophan treatment on yield and chemical composition of Berseem green fodder. Biosci. Res. 2018, 15, 3679–3694. [Google Scholar]
- Xu, Z.; Rothstein, S.J. ROS-Induced anthocyanin production provides feedback protection by scavenging ROS and maintaining photosynthetic capacity in Arabidopsis. Plant Signal. Behav. 2018, 13, e1451708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, A.J.; Scheiber, S.A.; Thomas, C. Inhibition of glutathione reductase by flavonoids. A structure-activity study. Biochem. Pharmacol. 1992, 44, 1603–1608. [Google Scholar] [CrossRef] [PubMed]
- Guangcha, O.; Yinglong, X. Physiological role and regulation of phenylpropanoid metabolism in plant. Plant Physiol. Commun. 1988, 3, 9–16. [Google Scholar]
- Johnson, G.; Schaal, L.A. Relation of Chlorogenic Acid to Scab Resistance in Potatoes. Science 1952, 115, 627–629. [Google Scholar] [CrossRef] [PubMed]
- Uritani, I.; Miyano, M. Derivatives of Caffeic Acid in Sweet Potato attacked by Black Rot. Nature 1955, 175, 812. [Google Scholar] [CrossRef]
- Johnson, G.; Schaal, L.A. Accumulation of phenolic substances and ascorbic acid in potato tuber tissue upon injury and their possible role in disease resistance. Am. J. Potato Res. 1957, 34, 200–209. [Google Scholar] [CrossRef]
- Tamagnone, L.; Mérida, A.; Stacey, N.; Plaskitt, K.; Parr, A.; Chang, C.-F.; Lynn, D.; Dow, J.M.; Roberts, K.; Martin, C. Inhibition of phenolic acid metabolism results in precocious cell death and altered cell morphology in leaves of transgenic tobacco plants. Plant Cell 1998, 10, 1801–1816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.J.; Li, Y.C.; Jiang, P.; Zhang, X.; Zhang, X.Y.; Qin, P.Y.; Tian, W. Identification of root exudates of wild chrysanthemum and its allelopathic effects on three plants. Hubei Agric. Sci. 2017, 56, 6. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 2016, e2584. [Google Scholar] [CrossRef] [Green Version]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, B.; Mei, Z.; Zeng, C.; Liu, S. metaX: A flexible and comprehensive software for processing metabolomics data. BMC Bioinform. 2017, 18, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulesteix, A.-L.; Strimmer, K. Partial least squares: A versatile tool for the analysis of high-dimensional genomic data. Brief. Bioinform. 2007, 8, 32–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Jia, S.; Wang, Y.; Hu, J.; Ding, Z.; Liang, Q.; Zhang, Y.; Wang, H. Mineral and metabolic profiles in tea leaves and flowers during flower development. Plant Physiol. Biochem. 2016, 106, 316–326. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Treatments | SEM | p-Value | |
|---|---|---|---|---|
| CK | T200 | |||
| Observed_species | 3349 | 4031 | 170 | 0.015 |
| Shannon | 8.951 | 9.732 | 0.263 | 0.485 |
| Simpson | 0.987 | 0.993 | 0.003 | 0.572 |
| Chao1 | 3753.329 | 4413.943 | 164.114 | 0.026 |
| ACE | 3823.739 | 4449.394 | 158.051 | 0.041 |
| Goods_coverage | 0.989 | 0.989 | 0.001 | 0.737 |
| Metabolites | Log2FC | Metabolites | Log2FC | Metabolites | Log2FC |
|---|---|---|---|---|---|
| 1,2,4-Benzenetriace | 5.27 | Gefitinib | 5.80 | Eriocitrin | 3.00 |
| Benzaldehyde | 5.67 | Omapatrilat | −2.44 | Tryptophol | 2.58 |
| Cinnamic acid | 5.55 | Gloriosine | 2.79 | gibberellin A1 | 4.00 |
| Benzoyl-beta-D-glucoside | 5.09 | Efinaconazole | 1.56 | Maculin | 2.44 |
| Setileuton | 7.31 | Pranlukast | 3.19 | 2-O-ETHYL ASCORBIC ACID | 2.97 |
| Isopropylmalic acid | 7.82 | Maltol | 3.50 | Codeinone | 3.07 |
| Dihydroxyphenylalanine | 3.22 | Cassyfiline | 3.22 | Levallorphan | 2.43 |
| Acetophenone | 4.56 | Myxochelin C | 4.68 | Quercetin | −4.89 |
| Dexloxiglumide | 5.98 | L-Lombricine | −2.86 | Drotaverine | −2.82 |
| Salicin 6-phosphate | 4.78 | MFCD15146035 | −1.87 | L-serine phosphoethanolamine | 2.15 |
| Embramine | 6.44 | Roquefortine E | 2.48 | 20-hydroxy-leukotriene E4 | 1.60 |
| SOA | 5.46 | Andrographolide | 2.68 | Xanthurenic acid | 1.64 |
| Oleuropein | 4.72 | Chlorogenic acid | −3.00 | Stachyflin | 2.47 |
| 7-Methoxy-4-methylcoumarin | 2.21 | Lactide | 2.57 | Naftalofos | 1.78 |
| Mps1-IN-1 | 3.31 | Isofraxidin | −2.02 | Dinocton 6 | 2.89 |
| Dimethyl itaconate | 3.09 | Oxandrolone | 2.21 | Kuwanon G | 3.04 |
| N-Hydroxydecanamide | −2.38 | Paramethadione | 2.24 | Guanidinobutyrate | 1.89 |
| TEMPO | 1.70 | Allantoic acid | 5.58 | Sennoside C | −4.39 |
| Repirinast | 3.31 | Istamycin C1 | 1.92 | Azlocillin | 2.80 |
| Coumarin | 3.47 | Nodakenin | 2.04 | Premithramycin A2′ | 2.74 |
| 8-Bromoguanosine | 4.27 | Homotrypanothione | 2.05 | Pevonedistat | −3.46 |
| Golotimod | 5.67 | Tyramine | 2.87 | Eriocitrin | 3.00 |
| Droxidopa | 2.42 | S-Bicalutamide | 2.91 | Taspine | 1.59 |
| Oxybutynin | 2.05 | 5-Aminopentanamide | 2.83 | Bis(glutathionyl)spermine | 5.78 |
| Valeramide | 2.81 | Payzone | −4.34 | Bis(glutathionyl)spermine disulfide | 2.04 |
| Salidroside | 1.82 | Glycerophosphoglycerol | −2.93 | Levallorphan | 2.43 |
| Dasolampanel | 3.16 | Dibenzylmethane | 2.16 | 7-ACA | −1.87 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Xu, J.; Li, Y.; He, Y.; Yang, Y.; Liu, D.; Wu, C. Effects of Coumarin on Rhizosphere Microbiome and Metabolome of Lolium multiflorum. Plants 2023, 12, 1096. https://doi.org/10.3390/plants12051096
Yang Y, Xu J, Li Y, He Y, Yang Y, Liu D, Wu C. Effects of Coumarin on Rhizosphere Microbiome and Metabolome of Lolium multiflorum. Plants. 2023; 12(5):1096. https://doi.org/10.3390/plants12051096
Chicago/Turabian StyleYang, Yihu, Jun Xu, Yan Li, Yuchen He, Yuqing Yang, Dalin Liu, and Caixia Wu. 2023. "Effects of Coumarin on Rhizosphere Microbiome and Metabolome of Lolium multiflorum" Plants 12, no. 5: 1096. https://doi.org/10.3390/plants12051096
APA StyleYang, Y., Xu, J., Li, Y., He, Y., Yang, Y., Liu, D., & Wu, C. (2023). Effects of Coumarin on Rhizosphere Microbiome and Metabolome of Lolium multiflorum. Plants, 12(5), 1096. https://doi.org/10.3390/plants12051096