
Metabolomic Analysis of Key Metabolites and Their Pathways Revealed the Response of Alfalfa (Medicago sativa L.) Root Exudates to rac-GR24 under Drought Stress


Abstract
:1. Introduction
2. Methods and Materials
2.1. Material Cultivation
2.2. Treatments
2.3. Determination of Triphenyltetrazolium Chloride (TTC), Soluble Sugar (SS), Soluble Protein (SP), and Malondialdehyde (MDA)
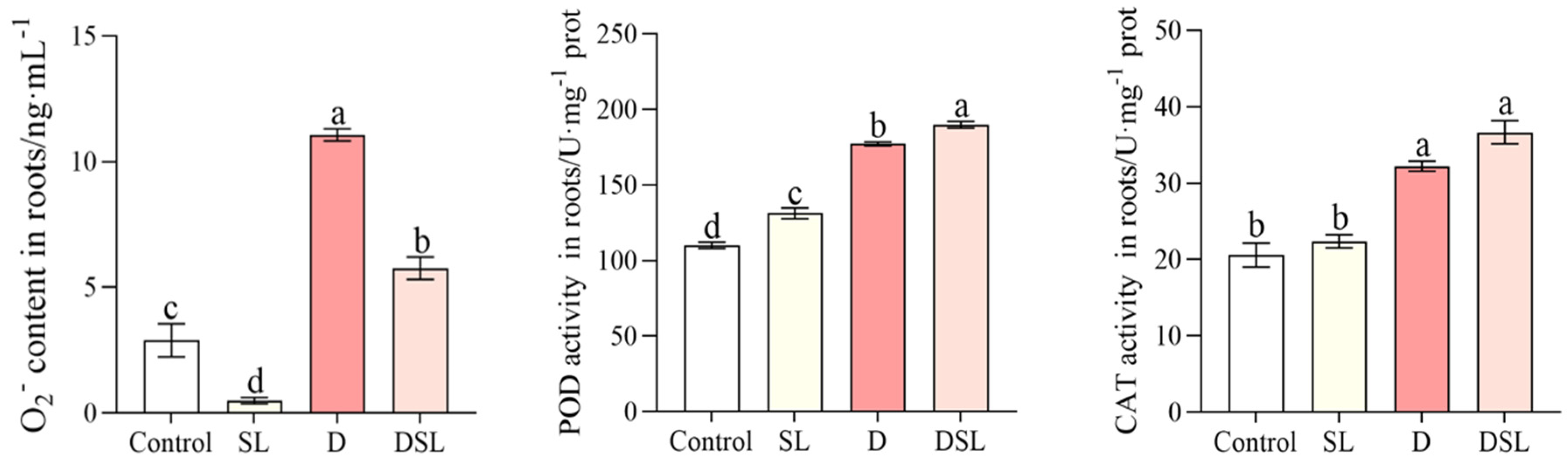
2.4. Determination of Strigolactone, O2−, and Enzyme Activity
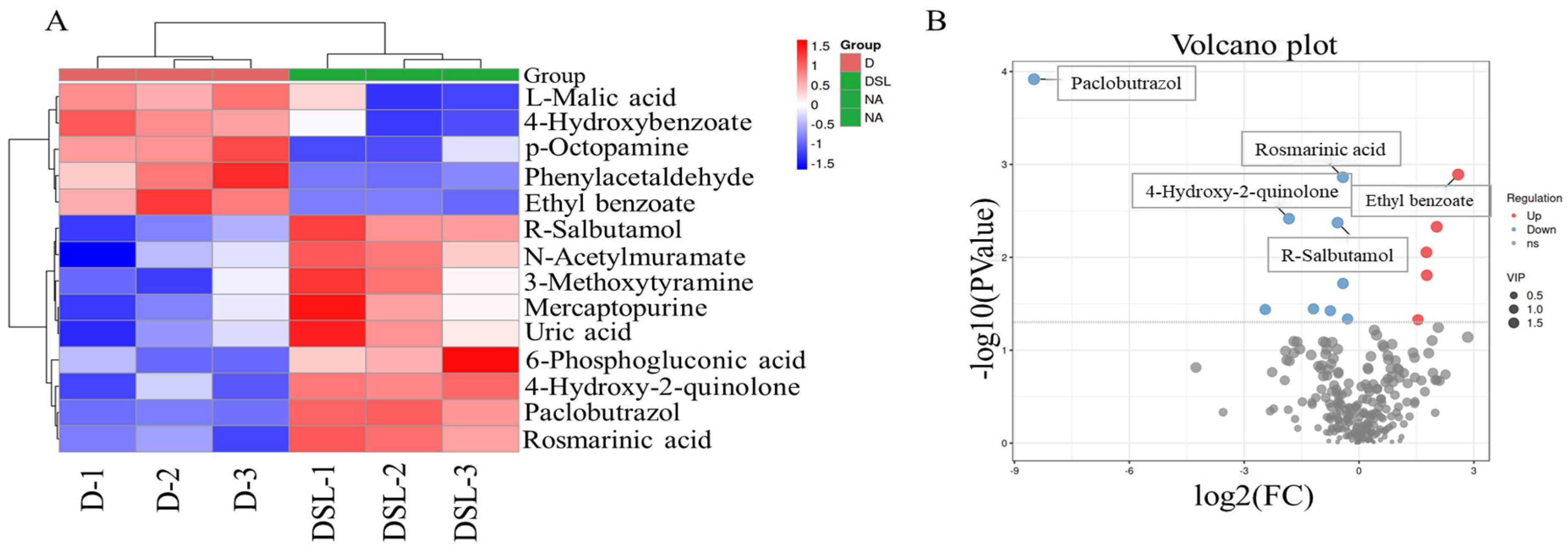
2.5. Untargeted Metabolomics Analysis
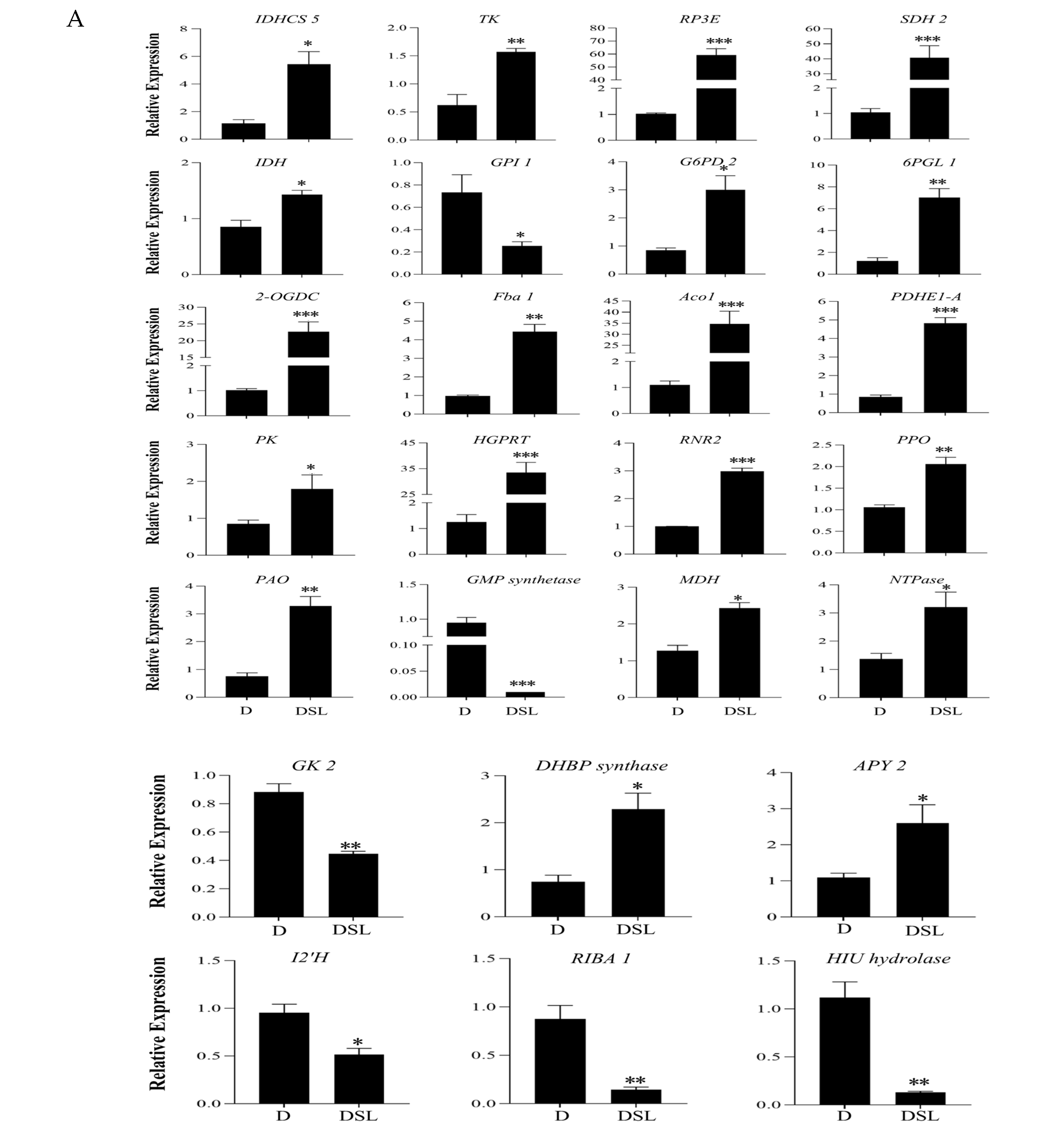
2.6. RNA Extraction and Gene Expression Analysis
2.7. Statistical Analysis
3. Results
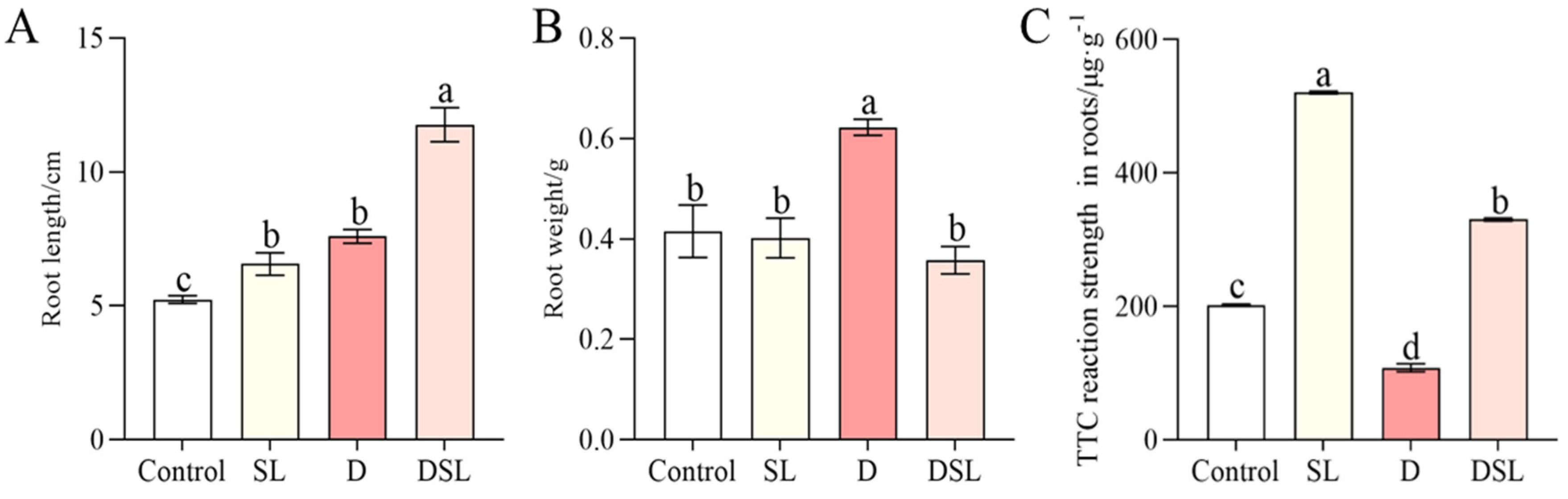
3.1. Morphological Response to Drought Stress in Alfalfa Root
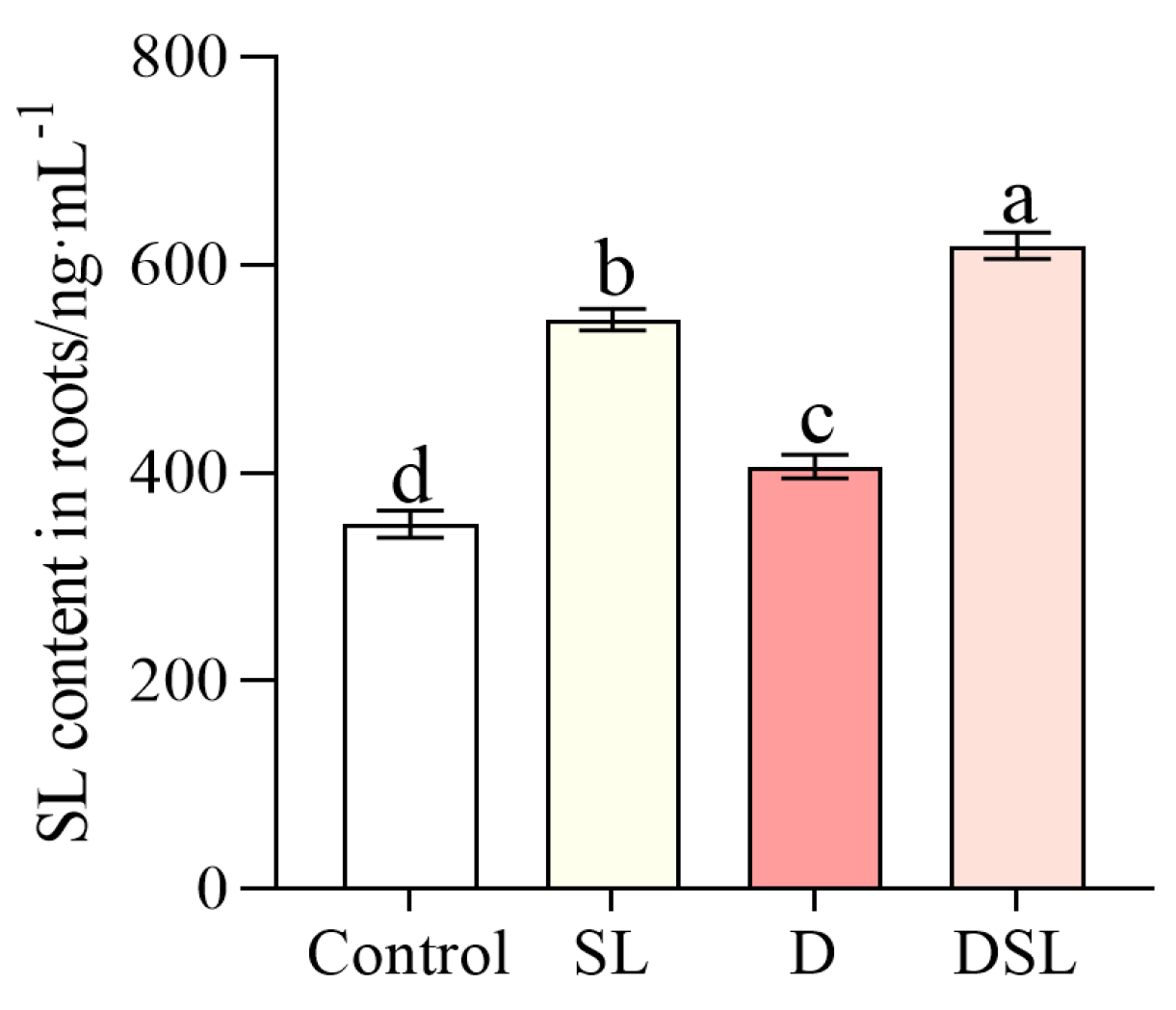
3.2. SL Response to Drought Stress in Alfalfa Root
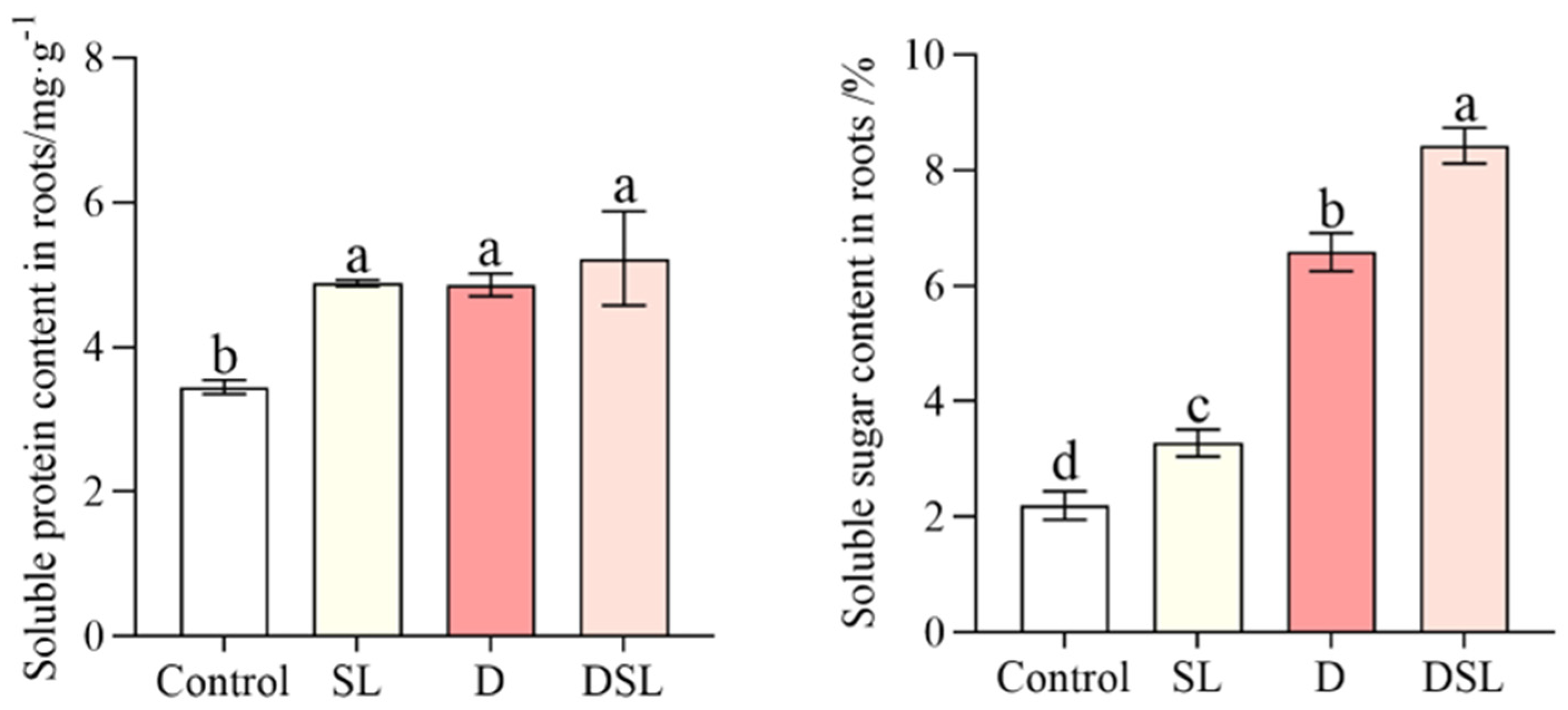
3.3. Osmotic Substance Response to Drought Stress in Alfalfa Root
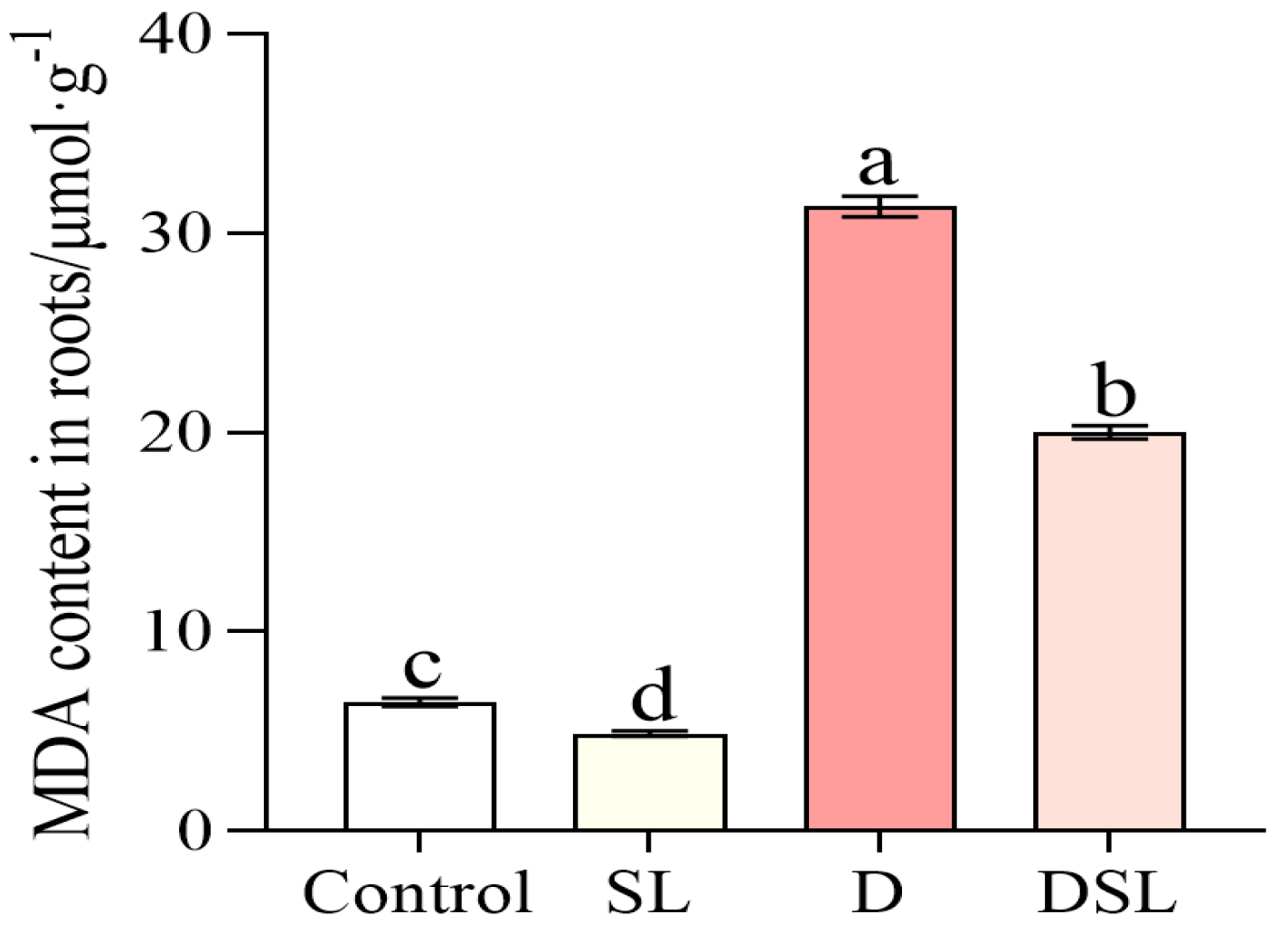
3.4. Membrane Lipid Peroxidation Response to Drought Stress in Alfalfa Root
3.5. Antioxidant Response to Drought Stress in Alfalfa Root
3.6. Metabolic Profiling
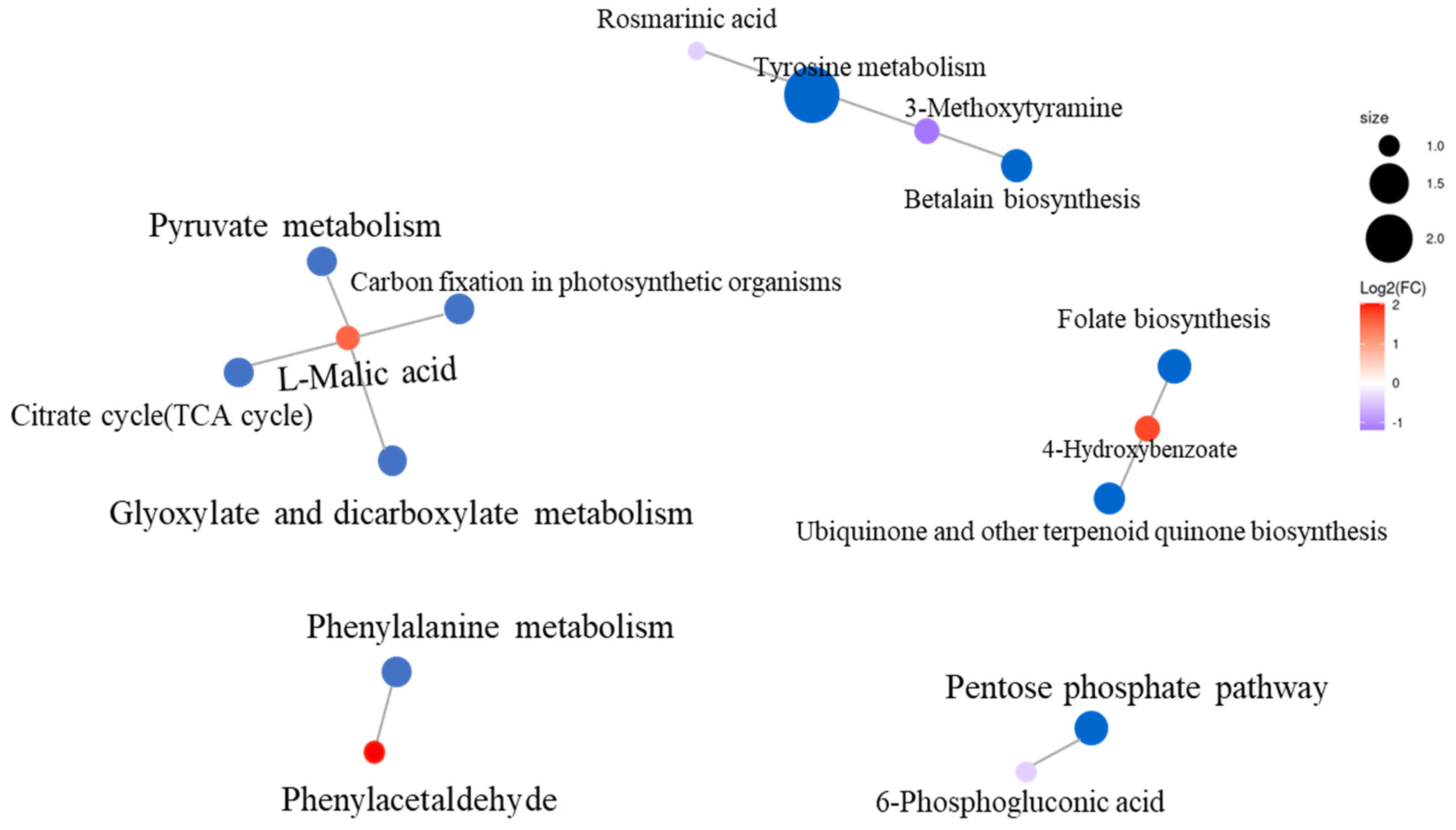
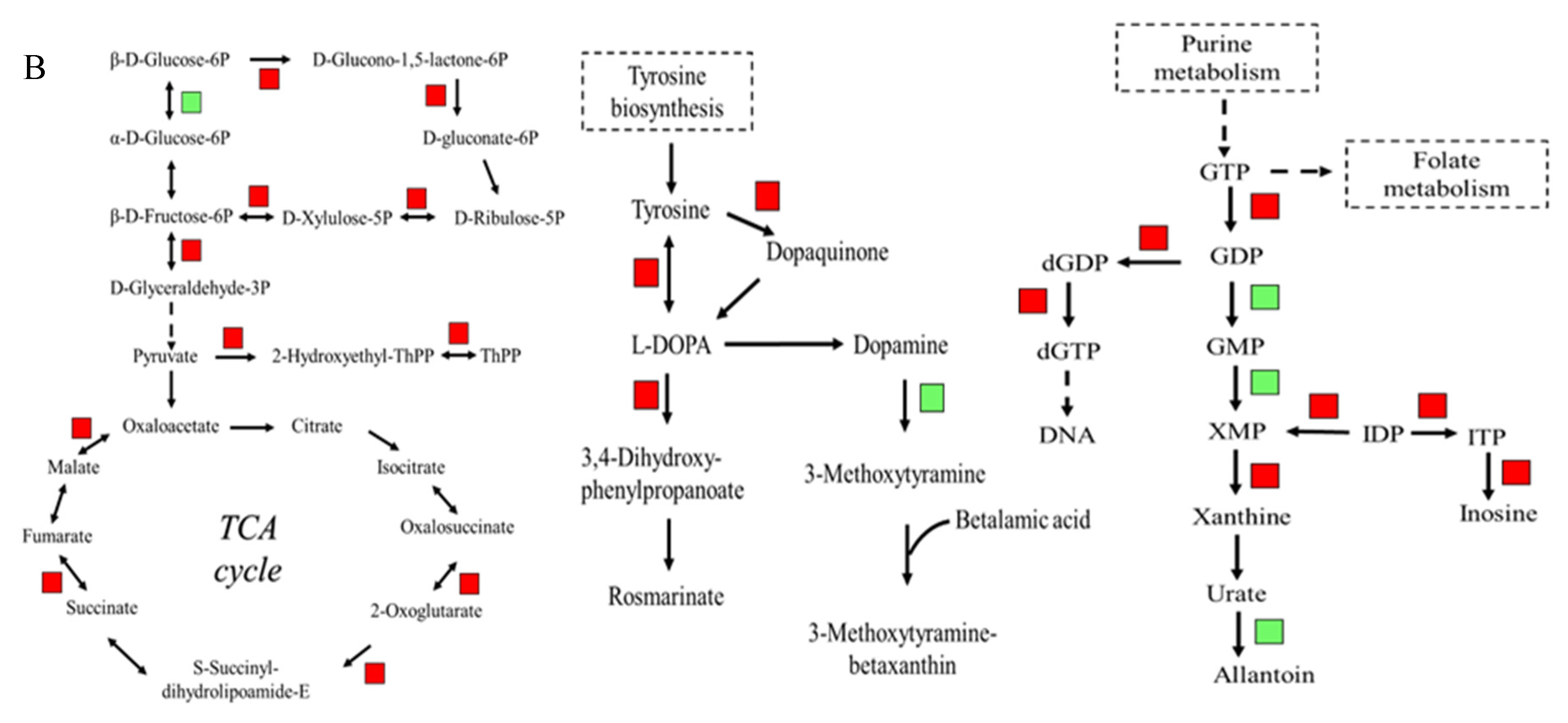
3.7. KEGG Enrichment Analysis
3.8. Genes Related to rac-GR24-Induced in Alfalfa Roots under Drought Stress
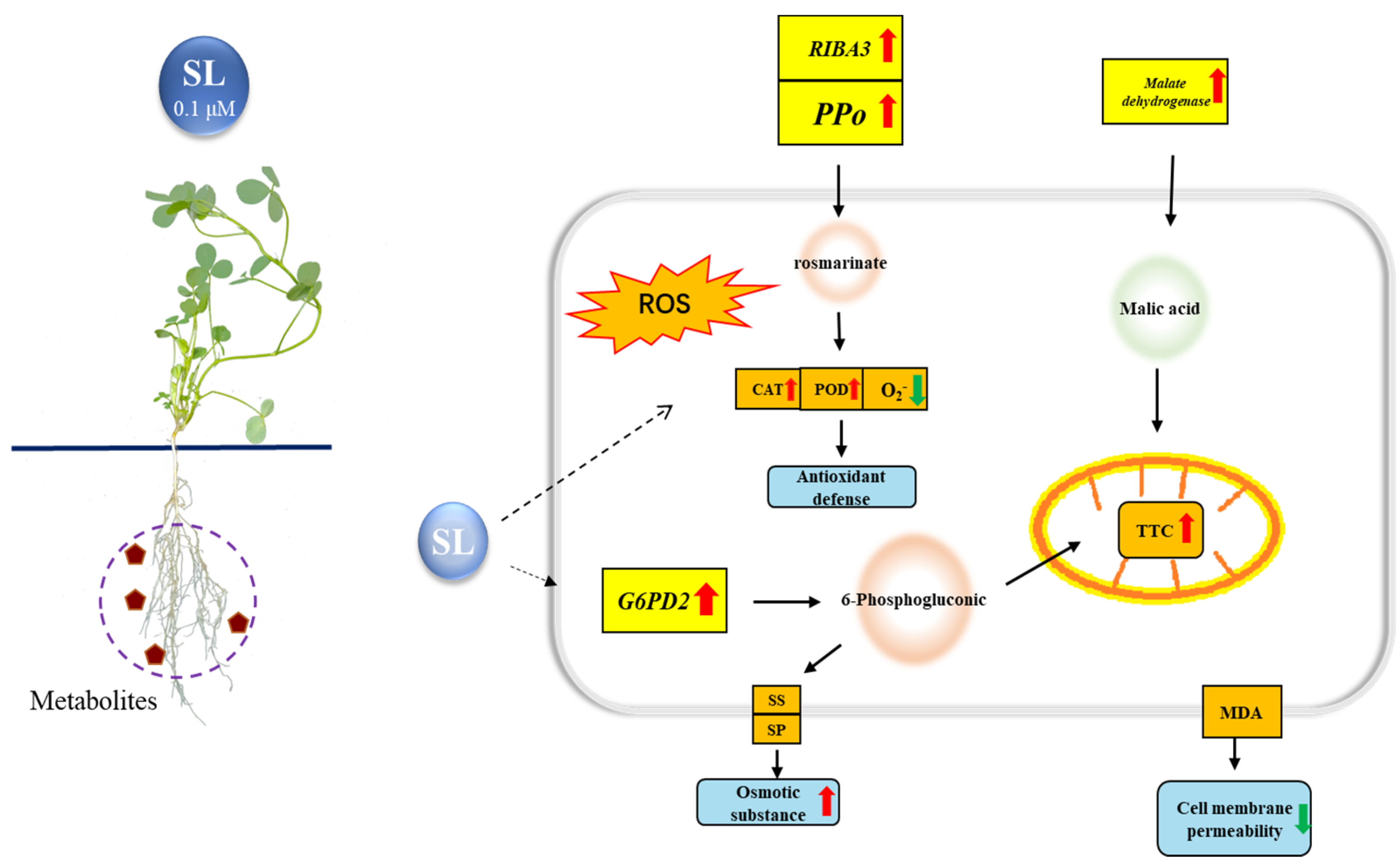
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Name | Short Name | Primer Sequence |
|---|---|---|---|
| MsG0780036878.01 | Glucose-6-phosphate 1-dehydrogenase 2 | G6PD 2 | F: AACCAACATGACACCACCGATACTG |
| R: AGACACGACACAACACACAGACAC | |||
| MsG0580028227.01 | Glucose-6-phosphate isomerase 1 | G6PI 1 | F: ATTGCAGCCATTTCACTAGCCATTG |
| R: GGTTTAGGTCTTGGTGGTGGGTTAG | |||
| MsG0580027701.01 | Probable 6-phosphogluconolactonase 1 | 6PGL 1 | F: CGTGCTCTATGGCGGCGTTAC |
| R: TCGGCTCCATTTCCTTCACAAACTC | |||
| MsG0480019171.01 | Transketolase | TK | F: TGGAATGGGAGCAATCAGCAATGG |
| R: CCTAATGGCACCACGCATGTAGTC | |||
| MsG0880046469.01 | Ribulose-phosphate 3-epimerase | RP3E | F: TTGAGAAGATTGTGCGTGGAGAAGG |
| R: ACAGCAGAACCAGCAACCAGTG | |||
| MsG0480021331.01 | Fructose-bisphosphate aldolase 1 | Fba 1 | F: TGCCATGAACCAATCTCCAAACCC |
| R: AGTGCTTCTTGTGCTGCCTGAAC | |||
| MsG0580025903.01 | Pyruvate dehydrogenase E1 component subunit alpha | PDHE1A | F: GATGGGTGCTCTAAAGGGAAAGGTG |
| R: CCGCAACCAAGTGGAACCTGAG | |||
| MsG0580025144.01 | Aconitate hydratase 1 | Aco 1 | F: GCTTCTCCTCCTCTCGTTGTTGC |
| R: TCTGCTATTTCTTCGCTGGATGGC | |||
| MsG0380017124.01 | Isocitrate dehydrogenase [NAD] catalytic subunit 5 | IDHCS 5 | F: TGTGTGCAGGCTGGAAAGTCAAG |
| R: CTCTCCAATCCGCAGGTTACCAAG | |||
| MsG0280009437.01 | Isocitrate dehydrogenase [NADP] | IDH | F: CCAGATGAAGGTCGTGTCAAGGAG |
| R: TTCGTCCAGCCTGAAACAAGTCG | |||
| MsG0780036931.01 | 2-oxoglutarate dehydrogenase | 2-OGDC | F: AAATATGTCGGTCGTGCTCCATCTG |
| R: TTGAAGGGCTTTCTCGGCAATCTC | |||
| MsG0180005032.01 | Succinate dehydrogenase [ubiquinone] iron–sulfur subunit 2 | SDH 2 | F: GGACGGATTCGACTTCGACGATAAC |
| R: CCTCTTCAACCACGGCTCAATACTC | |||
| MsG0780041035.01 | Malate dehydrogenase | MDH | F: GAGTTACAGCCCTCGATGTTGTGAG |
| R: TCCTGCATGACCTCCTACCACTG | |||
| MsG0280006694.01 | Polyphenol oxidase A1 | PPO | F: TGCTTTGCCATTTTGGAACTACGAC |
| R: GTTGGAGGTTGATGACTAGCGGTTC | |||
| MsG0280010998.01 | Monofunctional riboflavin biosynthesis protein RIBA 3 | DHBP synthase | F: TGGGTTAGGTCACAAACTTCAAGCC |
| R: TGCCATACTCACGAGCATCAACAG | |||
| MsG0480022833.01 | Isoflavone 2′-hydroxylase | I2′H | F: CACTGGCGTAACCTTCGTCGTATC |
| R: TCAACCTCTGAGTCTCGTCCCTTC | |||
| MsG0480020253.01 | Primary amine oxidase | PAO | F: AGATGTTGCATGGCCTGGTTGTC |
| R: TTGCCGCTGGAAGTGAGAATAGATC | |||
| MsG0780040024.01 | Nucleoside-triphosphatase | NTPase | F: GGTGGAGGAGGAAGTGGACAGAG |
| R: CAGTCTCAAGATCCACAGGACGAAG | |||
| MsG0780040027.01 | Apyrase 2 | APY2 | F: CGCCGTTCAATCCACTTCTCCTC |
| R: AACAAGAAGAGTGCCACGGAATCTG | |||
| MsG0480023648.01 | GMP synthase [glutamine-hydrolyzing] | GMP synthase | F: AGCAGGGTGGAAGATGGAGGATG |
| R: TGGTAGCAGCAACAGTGGAATCAAC | |||
| MsG0180004865.01 | Hydroxyisourate hydrolase | HIU hydrolase | F: GGAAGGAGCTGCTGATGAAGATGG |
| R: TTCGGGTTGACGGGTCCTCTAC | |||
| MsG0480022643.01 | Bifunctional riboflavin biosynthesis protein RIBA 1 | RIBA 1 | F: CGTGGCTCAGATTTGGCATTTGTTC |
| R: TCCAGAGGTCAACGCAGCTTTAAC | |||
| MsG0780040519.01 | Guanylate kinase 2 | GK2 | F: GGAGTAAGGGTGTGATAGGCAATGC |
| R: CTCGGAGCACGGGTTGTATGAC | |||
| MsG0180005342.01 | Hypoxanthine-guanine phosphoribosyltransferase | HGPRT | F: TCATCTTCCTTGCTGACCTTGTTCG |
| R: TGGAGACAGTAGCAGAACCGTAGG | |||
| MsG0880046930.01 | Ribonucleoside-diphosphate reductase small chain A | RNR2 | F: AAGGAAGGCTGAATGGGCATTGAG |
| R: GCAAGCAAAATCGCAGTGAAGACC | |||
| MsG0180005028.01 | Pyruvate kinase 1 | PK | F: GAGCCGATGAGCCACATGGAATC |
| R: CAGGCATGGTTGGTCTGTACTTAGC |
References
- Wu, X.; Cai, K.; Zhang, G.P.; Zeng, F.R. Metabolite profiling of barley grains subjected to water stress: To explain the genotypic difference in drought-induced impacts on malting quality. Front. Plant Sci. 2017, 8, 1547. [Google Scholar] [CrossRef] [PubMed]
- Mi, Z.R.; Ma, Y.Y.; Liu, P.L.; Zhang, H.Y.; Zhang, L.; Jia, W.Q.; Zhu, X.P.; Wang, Y.L.; Zhang, C.; Du, L.; et al. Combining metabolic analysis with biological endpoints provides a view into the drought resistance mechanism of Carex breviculmis. Front. Plant Sci. 2022, 13, 945441. [Google Scholar] [CrossRef]
- Zaher-Ara, T.; Boroomand, N.; Sadat-Hosseini, M. Physiological and morphological response to drought stress in seedlings of ten citrus. Trees 2016, 30, 985–993. [Google Scholar] [CrossRef]
- Ma, X.S.; Xia, H.; Liu, Y.H.; Wei, H.B.; Zheng, X.G.; Song, C.Z.; Chen, L.; Liu, H.Y.; Luo, L.J. Transcriptomic and metabolomic studies disclose key metabolism pathways contributing to well-maintained photosynthesis under the drought and the consequent drought-tolerance in rice. Front. Plant Sci. 2016, 7, 1886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, F.D.; Wang, X.J.; Hou, L.; Zhao, S.Z.; Li, G.H.; Xia, H. Research progress in plant metabolomics. Shandong Agric. Sci. 2017, 49, 154–162. [Google Scholar] [CrossRef]
- Dixon, R.A.; Strack, D. Phytochemistry meets genome analysis, and beyond. Phytochemistry 2003, 62, 815–816. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.W.; Wang, G.D. Application of LC-MS in plant metabolomic analysis. Chin. Bull. Life Sci. 2015, 27, 978–985. [Google Scholar] [CrossRef]
- Sanchez, D.H.; Schwabe, F.; Erban, A.; Udvardi, M.K.; Kopka, J. Comparative metabolomics of drought acclimation in model and forage legumes. Plant Cell Environ. 2011, 35, 136–149. [Google Scholar] [CrossRef]
- Ullah, N.; Yüce, M.; Gökçe, Z.N.Ö.; Budak, H. Comparative metabolite profiling of drought stress in roots and leaves of seven Triticeae species. BMC Genom. 2017, 18, 969. [Google Scholar] [CrossRef]
- Moradi, P.; Ford-Lloyd, B.; Pritchard, J. Metabolomic approach reveals the biochemical mechanisms underlying drought stress tolerance in thyme. Anal. Biochem. 2017, 527, 49–62. [Google Scholar] [CrossRef] [Green Version]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 62, 1593–1608. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, D.; Kannan, M.; Reddy, A.R. A root proteomics-based insight reveals dynamic regulation of root proteins under progressive drought stress and recovery in Vigna radiata L. wilczek. Planta 2011, 233, 1111–1127. [Google Scholar] [CrossRef]
- Benlloch-Gonzalez, M.; Sanchez-Lucas, R.; Benlloch, M. Effects of olive root warming on potassium transport and plant growth. J. Plant Physiol. 2017, 218, 182–188. [Google Scholar] [CrossRef]
- Ibrahim, R.S.; El-Mezayen, N.S.; Khairy, A.; Zaatout, H.H.; Hammoda, H.M.; Metwallly, A.M. Biologically-guided isolation of natural lead antithyroid drug from Medicago sativa L. Sprouts and Its Toxic Profile in Comparison with Propylthiouracil. J. Food Drug Anal. 2020, 28, 407–448. [Google Scholar] [CrossRef]
- Hadi, F.; Ayaz, M.; Ali, S.; Shafiq, M.; Ullah, R.; Jan, A. Comparative effect of polyethylene glycol and mannitol induced drought on growth (in vitro) of canola (Brassica napus), cauliflower (Brassica oleracea) and tomato (Lycopersicon esculentum) seedlings. Int. J. Biosci. 2014, 4, 34–41. [Google Scholar] [CrossRef]
- Luke, O.; Manoj, G.K.; Jeffrey, F.F.; Tomas, P.; Miroslav, S.; Johannes, V.S. Synthetic strigolactone (rac-GR24) alleviates the adverse effects of heat stress on seed germination and photosystem II function in lupine seedlings. Plant Physiol. Biochem. 2020, 155, 965–979. [Google Scholar] [CrossRef]
- Iftikhar, I.; Shahbaz, M.; Wahid, M.A. Potential role of foliage applied strigolactone (GR24) on photosynthetic pigments, gas exchange attributes, mineral nutrients and yield components of Zea mays (L.) under saline regimes. Gesunde Pflanz. 2022. [Google Scholar] [CrossRef]
- Wang, N.Q.; Kong, C.H.; Wang, P.; Meiners, S.J. Root exudate signals in plant-plant interactions. Plant Cell Environ. 2021, 44, 1044–1058. [Google Scholar] [CrossRef]
- Yang, Y.W.; Gu, M.Z.; Chen, J.M.; Zhang, R.L.; Liu, Z.Y.; Shi, Y.H.; Liu, D.L.; Wang, L. Comparative Transcriptomes Reveal the Mitigation Effect of GR24 in Alfalfa Under Drought Stress. J. Plant Growth Regul. 2022. [Google Scholar] [CrossRef]
- Yang, Y.W.; Gu, M.Z.; Lu, J.F.; Li, X.T.; Liu, D.L.; Li, X.E.; Wang, L. GR24 alleviates the adverse effects of drought stress on physiology and photosystem II function in alfalfa (Medicago sativa L.). Grassl. Sci. 2022. [Google Scholar] [CrossRef]
- Clememsson-Lindell, A. Triphenyltetrazolium chloride as an indicator of fine-root vitality and environmental stress in coniferous forest stands: Applications and limitations. Plant Soil 1994, 159, 297–300. [Google Scholar] [CrossRef]
- Buysse, J.; Merckx, R. An improved colorimetric method to quantify sugar content of plant tissue. J. Exp. Bot. 1993, 44, 1627–1629. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–265. [Google Scholar] [CrossRef]
- Liu, X. Experimental Guidance of Plant Physiology, 1st ed.; China Agriculture: Beijing, China, 2015; pp. 15–16. [Google Scholar]
- Blume, D.E.; McClure, J.W. Developmental Effects of Sandoz 6706 on Activities of Enzymes of Phenolic and General Metabolism in Barley Shoots Grown in the Dark or under Low or High Intensity Light. Plant Physiol. 1980, 65, 238–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranieri, A.; Petacco, F.; Castagna, A.; Soldatini, G.F. Redox state and peroxidase system in sunflower plants exposed to ozone. Plant Sci. 2000, 1, 159–167. [Google Scholar] [CrossRef]
- Ling, F.; Su, Q.; Jiang, H.; Cui, J.; He, X.; Wu, Z.H.; Zhang, Z.A.; Liu, J.; Zhao, Y.J. Effects of strigolactone on photosynthetic and physiological characteristics in salt-stressed rice seedlings. Sci. Rep. 2020, 10, 6183. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.H.; Ma, C.; Zhang, L.; Su, M.; Wang, J.; Zheng, S.; Zhang, T.G. GR24-mediated enhancement of salt tolerance and roles of H2O2 and Ca2+ in regulating this enhancement in cucumber. J. Plant Physiol. 2022, 270, 153640. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Breusegem, F.V. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef]
- Wang, Z. Plant Physiology, 4th ed.; China Agriculture: Beijing, China, 2014; pp. 202–206. [Google Scholar]
- Arkorful, E.; Hu, S.K.; Zou, Z.W.; Yu, Y.; Chen, X.; Li, X.H. Metabolomic Analyses Provide New Insights into Signaling Mechanisms for Nutrient Uptake by Lateral Roots of Pruned Tea Plant (Camellia sinensis). J. Agric. Food Chem. 2020, 68, 7890–7903. [Google Scholar] [CrossRef]
- Kong, L.; Liu, J.; Han, Q.; Zhou, Q.X.; He, J.L. Integrating metabolomics and physiological analysis to investigate the toxicological mechanisms of sewage sludge derived biochars to wheat. Ecotoxicol. Environ. Saf. 2019, 185, 109664. [Google Scholar] [CrossRef]
- Watanabe, S.; Kounosu, Y.; Shimada, H.; Sakamoto, A. Arabidopsis xanthine dehydrogenase mutants defective in purine degradation show a compromised protective response to drought and oxidative stress. Plant Biotechnol. 2014, 31, 173–178. [Google Scholar] [CrossRef] [Green Version]
- Boo, Y.C.; Jung, J. Alteration in pyridine nucleotide status in cells as an adaptive response to water stress in Rice (Oryza sativa L.) seedlings. Agric. Chem. Biotechnol. 1998, 41, 228–234. [Google Scholar]
- Zhang, C.L.; Chen, J.H.; Huang, W.X.; Song, X.; Niu, J. Transcriptomics and Metabolomics Reveal Purine and Phenylpropanoid Metabolism Response to Drought Stress in Dendrobium sinense, an Endemic Orchid Species in Hainan Island. Front. Genet. 2021, 12, 692702. [Google Scholar] [CrossRef]
- Zhao, D.K.; Shi, Y.N.; Senthilkumar, H.A.; Qiao, Q.; Wang, Q.X.; Shen, Y.; Hu, G.W. Enriched networks ‘nucleoside/nucleotide and ribonucleoside/ribonucleotide metabolic processes’ and ‘response to stimulus’ potentially conferred to drought adaptation of the epiphytic orchid Dendrobium wangliangii. Physiol. Mol. Biol. Plants 2019, 25, 31–45. [Google Scholar] [CrossRef]
- Zoref-Shani, E.; Bromberg, Y.; Brosh, S.; Sidi, Y.; Sperling, O. Characterization of the alterations in purine nucleotide metabolism in hypoxanthine-guanine phosphoribosyltransferase-deficient rat neuroma cell line. J. Neurochem. 1993, 61, 457–463. [Google Scholar] [CrossRef]
- Xu, J.J.; Fang, X.; Li, C.Y.; Yanng, L.; Chen, X.Y. General and specialized tyrosine metabolism pathways in plants. aBIOTECH 2020, 1, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Falk, J.; Munne-Bosch, S. Tocochromanol functions in plants: Antioxidation and beyond. J. Exp. Bot. 2010, 61, 1549–1566. [Google Scholar] [CrossRef] [Green Version]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Gu, M.; Lu, J.; Li, X.; Liu, D.; Wang, L. Metabolomic Analysis of Key Metabolites and Their Pathways Revealed the Response of Alfalfa (Medicago sativa L.) Root Exudates to rac-GR24 under Drought Stress. Plants 2023, 12, 1163. https://doi.org/10.3390/plants12051163
Yang Y, Gu M, Lu J, Li X, Liu D, Wang L. Metabolomic Analysis of Key Metabolites and Their Pathways Revealed the Response of Alfalfa (Medicago sativa L.) Root Exudates to rac-GR24 under Drought Stress. Plants. 2023; 12(5):1163. https://doi.org/10.3390/plants12051163
Chicago/Turabian StyleYang, Yuwei, Mingzhou Gu, Junfeng Lu, Xin’e Li, Dalin Liu, and Lin Wang. 2023. "Metabolomic Analysis of Key Metabolites and Their Pathways Revealed the Response of Alfalfa (Medicago sativa L.) Root Exudates to rac-GR24 under Drought Stress" Plants 12, no. 5: 1163. https://doi.org/10.3390/plants12051163
APA StyleYang, Y., Gu, M., Lu, J., Li, X., Liu, D., & Wang, L. (2023). Metabolomic Analysis of Key Metabolites and Their Pathways Revealed the Response of Alfalfa (Medicago sativa L.) Root Exudates to rac-GR24 under Drought Stress. Plants, 12(5), 1163. https://doi.org/10.3390/plants12051163