
Does Potassium Modify the Response of Zinnia (Zinnia elegans Jacq.) to Long-Term Salinity?



Abstract
:1. Introduction
2. Results
2.1. Results of the Preliminary Experiment
2.2. Results of Main Experiments
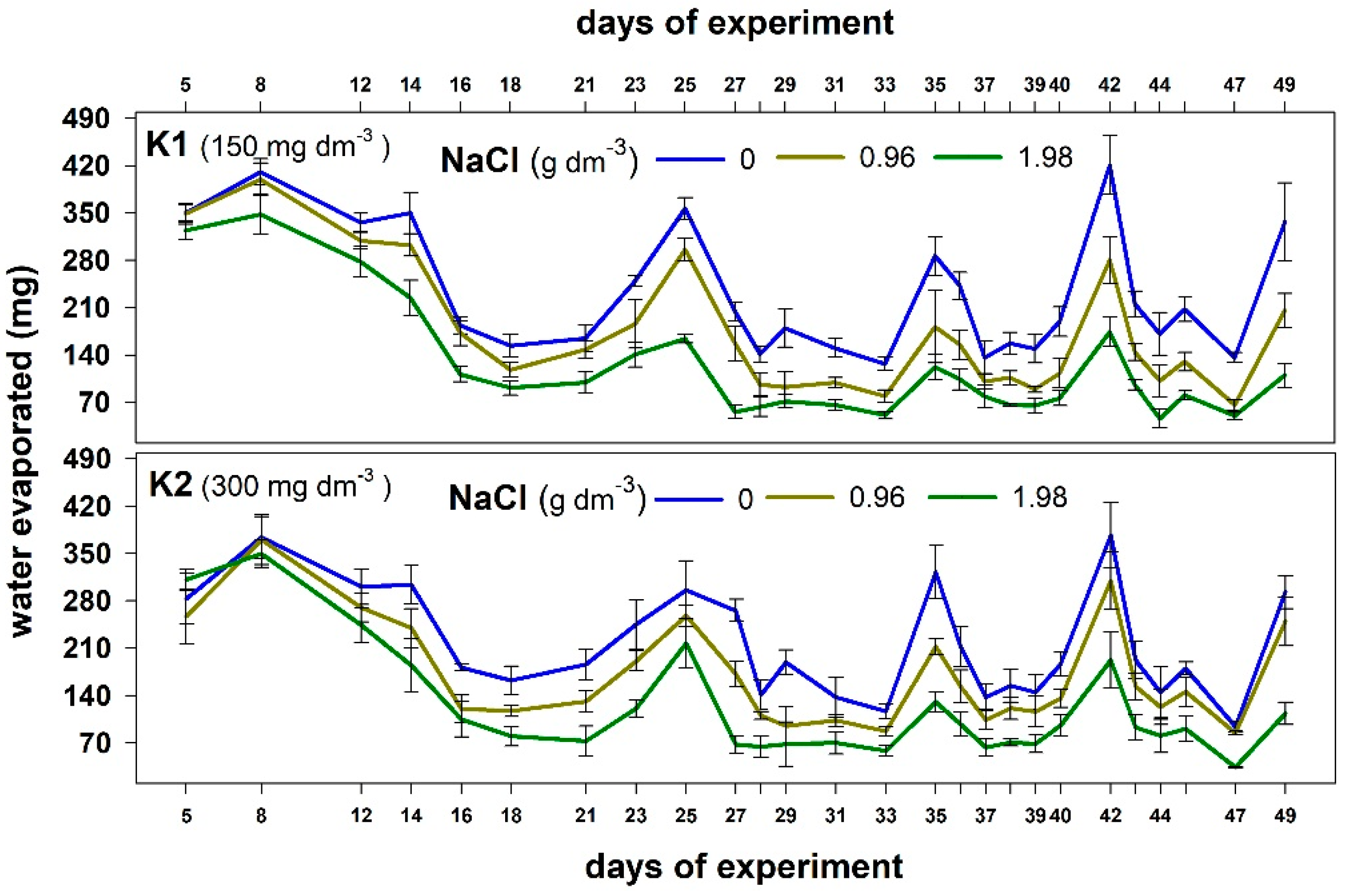
2.2.1. Water Management Parameters
2.2.2. Na, Cl, and K Concentrations in Leaves
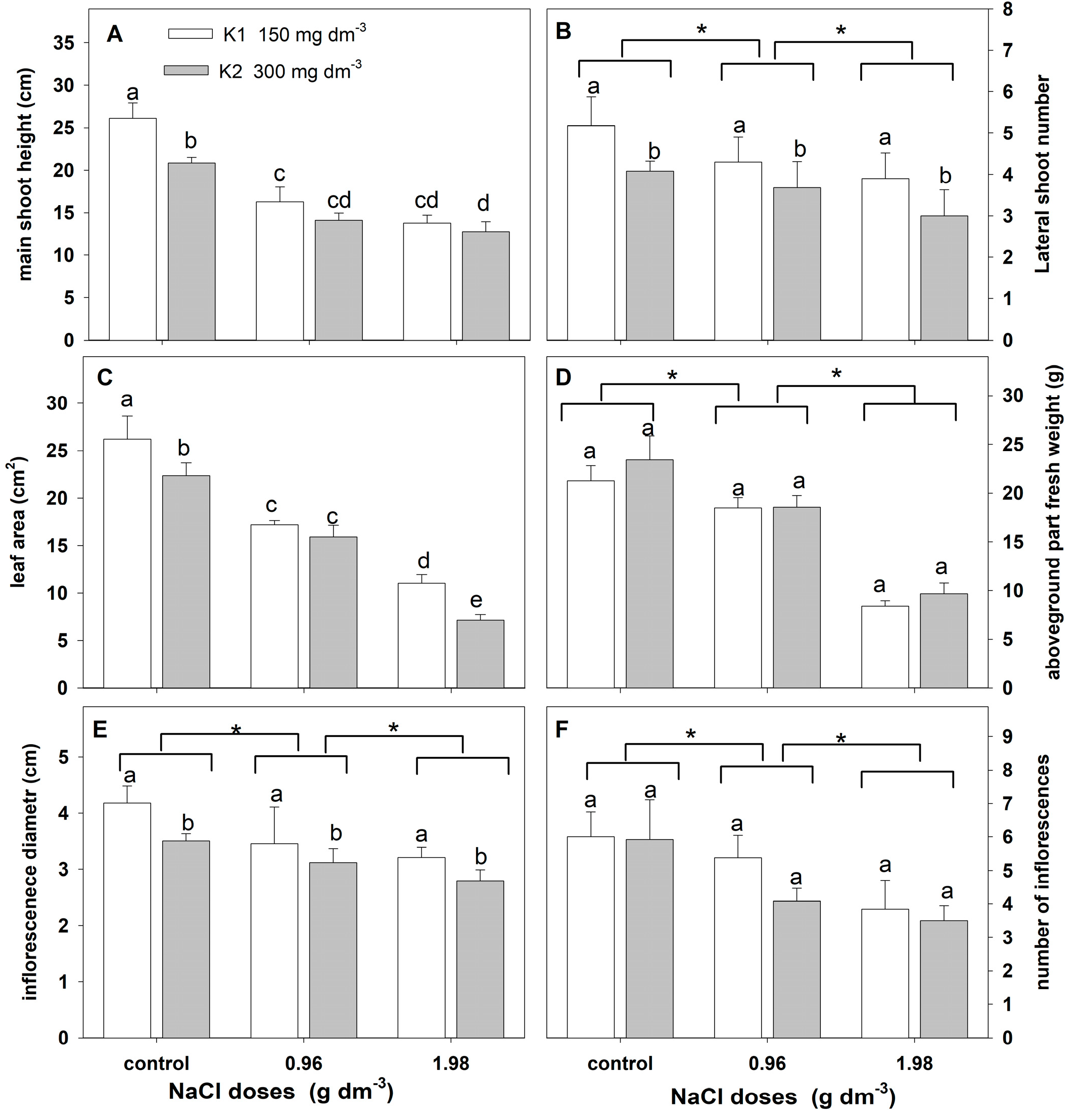
2.2.3. Growth Parameters
2.2.4. Chlorophyll, Lignin, and Membrane Injury Index
2.2.5. Proline Content and Enzyme Activity
3. Discussion
4. Materials and Methods
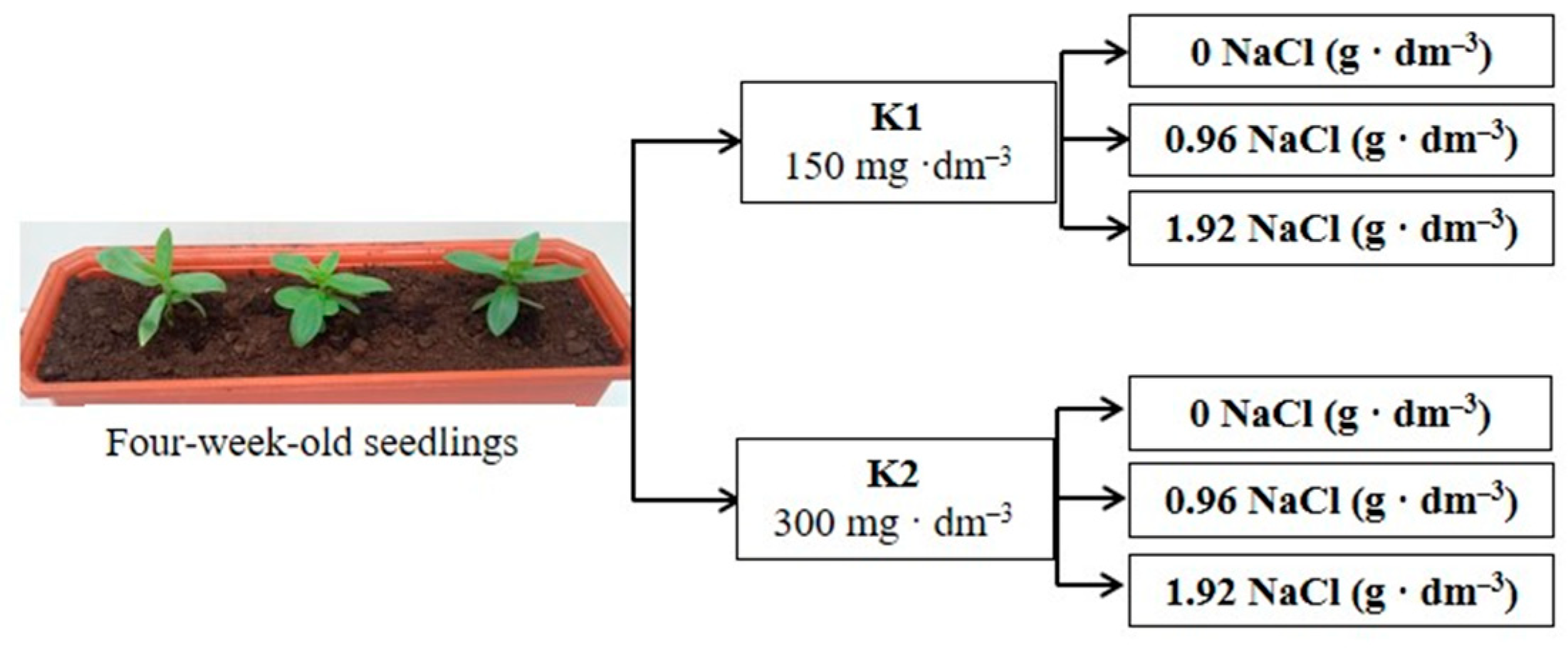
4.1. Plant Cultivation and Treatment
4.2. Water Loss Measurements
4.3. Growth Measurements
4.4. Na, Cl, and K Measurements
4.5. Physiological and Biochemical Parameter Measurements
4.6. Water Content in Leaves
4.7. Proline
4.8. Enzyme Extraction and Assays
4.9. Protein Concentration
4.10. Chlorophyll Content
4.11. Lignin Determination
4.12. Membrane Injury Index
4.13. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mishra, A.; Tanna, B. Halophytes: Potential resources for salt stress tolerance genes and promoters. Front. Plant Sci. 2017, 8, 829. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, E.A. Seed priming to alleviate salinity stress in germinating seeds. J Plant Physiol. 2016, 192, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Rengasamy, P. World salinization with emphasis on Australia. J. Exp. Bot. 2006, 57, 1017–1023. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.; Irfan, M.; Ahmad, A.; Hayat, A. Causes of salinity and plant manifestations to salt stress: A review. J. Environ. Biol. 2011, 32, 667–685. [Google Scholar] [PubMed]
- Singh, M.; Singh, A.; Prasad, S.M.; Singh, R.K. Regulation of plants metabolisms in response to salt stress: An omics approach. Acta Physiol. Plant. 2017, 39, 48. [Google Scholar] [CrossRef]
- Guo, J.; Shan, C.; Zhang, Y.; Wang, X.; Tian, H.; Han, G.; Zhang, Y.; Wang, B. Mechanisms of Salt Tolerance and Molecular Breeding of Salt-Tolerant Ornamental Plants. Front. Plant Sci. 2022, 13, 854116. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Nara, U.; Kumar, A.; Choudhary, A.; Singh, H.; Thapa, S. Salinity tolerance mechanisms and their breeding implications. J. Genet. Eng. Biotechnol. 2021, 19, 173. [Google Scholar] [CrossRef]
- García-Caparrós, P.; Lao, M.T. The effects of salt stress on ornamental plants and integrative cultivation practices. Sci. Hort. 2018, 240, 430–439. [Google Scholar] [CrossRef]
- Devecchi, M.; Remotti, D. Effect of Salts on Ornamental Ground Covers for Green Urban Areas. Acta Hortic. 2004, 643, 153–156. [Google Scholar] [CrossRef]
- Cunningham, M.A.; Snyder, E.; Yonkin, D.; Ross, M.; Elsen, T. Accumulation of deicing salts in soils in urban environment. Urban Ecosyst. 2008, 11, 17–31. [Google Scholar] [CrossRef]
- Ordóñez-Barona, C.; Sabetski, V.; Millward, A.A.; Steenberg, J. De-icing salt contamination reduces urban tree performance in structural soil cells. Environ. Pollut. 2018, 253, 562–571. [Google Scholar] [CrossRef] [PubMed]
- Cassaniti, C.; Romano, D.; Flowers, T.J. The response of ornamental plants to saline irrigation water. In Irrigation–Water Management, Pollution and Alternative Strategies; Garcia-Garizabal, I., Ed.; InTechOpen: London, UK, 2012; pp. 131–139. [Google Scholar] [CrossRef] [Green Version]
- Reid, M.S.; Jiang, C.Z. Postharvest biology and technology of cut flowers and potted plants. Hortic. Rev. 2012, 40, 1–54. [Google Scholar]
- Villarino, G.H.; Mattson, N.S. Assessing tolerance to sodium chloride salinity in fourteen floriculture species. HortTechnology 2011, 21, 539–545. [Google Scholar] [CrossRef] [Green Version]
- Esringü, A.; Ekinci, M.; Turan, M. Effects of Different Growing Media on Growth Parameters of Zinnia (Zinnia elegans). Yuzuncu Yil University. J. Agric. Sci. 2022, 32, 175–185. [Google Scholar] [CrossRef]
- Iannotti, M. How to plant and care for Zinnia flowers. The Spruce 2022. Available online: https://www.thespruce.com/zinnias-a-burst-of-hot-flower-colors-from-zinnias-1402919 (accessed on 15 December 2022).
- Oliveira, E.V.; de Lacerda, C.F.; de Neves, A.L.R.; Gheyi, H.R.; Oliveira, D.R.; Oliveira, F.Í.F.; de Viana, T.V.A. A new method to evaluate salt tolerance of ornamental plants. Theor. Exp. Plant Physiol. 2018, 30, 173–180. [Google Scholar] [CrossRef]
- Niu, G.; Wang, M.; Rodriges, D. Response of Zinna plants to saline water irrigation. HortScience 2012, 478, 793–797. [Google Scholar] [CrossRef] [Green Version]
- Macherla, K.; McAvoy, R.J. The effect of salinity on the growth and nutrient status of zinnia grown under short- and long-cycle subirrigation management. HortScience 2017, 52, 770–773. [Google Scholar] [CrossRef]
- Bizhaini, S.; Jowkar, A.; Abdolmaleki, M. Growth and antioxidant response of Zinnia elegans under salt stress conditions. Techn. J. Eng. Appl. Sci. 2013, 13, 1285–1292. [Google Scholar]
- Escalona, A.; Salas-Sanjuán, M.C.; Dos Santos, C.; Guzmán, M. The effect of water salinity on growth and ionic concentration and relation in plant tissues in Zinnia elegans and Tagetes erecta for use in urban landscaping. ITEA 2014, 110, 325–334. [Google Scholar] [CrossRef]
- Carter, C.T.; Grieve, C.M. Growth and nutrition of two cultivars of Zinnia elegans under saline conditions. HortScience 2010, 45, 1058–1063. [Google Scholar] [CrossRef] [Green Version]
- Van Zelm, E.; Zhang, Y.; Testerink, C. Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isayenkov, S.V.; Maathuis, F.J. Plant salinity stress: Many unanswered questions remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.; Choudhary, K.K.; Chaudhary, N.; Gupta, S.; Sahu, M.; Tejaswini, B.; Sarkar, S. Salt stress resilience in plants mediated through osmolyte accumulation and its crosstalk mechanism with phytohormones. Front. Plant Sci. 2022, 13, 1006617. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Ann. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant Responses to Salt Stress: Adaptive Mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Rademacher, W. Plant growth regulators: Backgrounds and uses in plant production. J. Plant Growth Regul. 2015, 34, 845–872. [Google Scholar] [CrossRef]
- Meena, M.; Divyanshu, K.; Kumar, S.; Swapnil, P.; Zehra, A.; Shukla, V.; Yadav, M.; Upadhyay, R.S. Regulation of l-proline biosynthesis, signal transduction, transport, accumulation and its vital role in plants during variable environmental conditions. Heliyon 2019, 5, e02952. [Google Scholar] [CrossRef] [Green Version]
- Szabados, L.; Savourè, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2009, 15, 89–97. [Google Scholar] [CrossRef]
- Abbasi, H.; Jamil, M.; Haq, A.; Ali, S.; Ahmad, R.; Malik, Z.; Parveen. Salt Stress Manifestation on Plants, Mechanism of Salt Tolerance and Potassium Role in Alleviating It: A Review. Zemdirbyste-Agriculture 2016, 103, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Sayed, S.A.A.E.; Hellal, F.A.; El–Rab, N.G.; Zewainy, R.M. Ameliorative Effects of Potassium on the Salinity Stress in Plants: A Review. Asian J. Soil Sci. Plant Nutr. 2019, 4, 1–15. [Google Scholar] [CrossRef]
- Ben-Hayyim, G.; Spiegel-Roy, P.; Neumann, H. Relation between ion accumulation of salt-sensitive and isolated stable salttolerant cell lines of Citrus aurantium. Plant Physiol. 1985, 178, 144–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzortzakis, N.G. Potassium and calcium enrichment alleviate salinity-induced stress in hydroponically grown endives. Hort. Sci. 2010, 37, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Umar, S.; Diva, I.; Anjum, N.; Iqbal, M.; Ahmad, I.; Pereira, E. Potassium-induced alleviation of salinity stress in Brassica campestris L. Open Life Sci. 2011, 6, 1054–1063. [Google Scholar] [CrossRef]
- Amjad, M.; Akhtar, J.; Haq, M.; Aanwar-ul-Haq, M.; Imran, S.; Jacobsen, S.E. Soil and foliar application of potassium enhances fruit yield and quality of tomato under salinity. Turk. J. Biol. 2014, 38, 7. [Google Scholar] [CrossRef]
- Hashi, U.S.; Karim, A.; Saikat, H.M.; Islam, R.; Islam, M.A. Effect of Salinity and Potassium Levels on Different Morpho-Physiological Characters of two Soybean (Glycine max L.) Genotypes. J. Rice Res. 2015, 3, 143. [Google Scholar] [CrossRef]
- Henry, E.E.Y.; Kinsou, E.; Mensah, A.C.G.; Komlan, F.A.; Gandonou, C.B. Response of tomato (Lycopersicon esculentum mill.) plants cultivated under salt stress to exogenous application of calcium and potassium. J. Appl. Biosci. 2021, 159, 16363–16370. [Google Scholar] [CrossRef]
- Yurtseven, E.; Kesmez, G.D.; Ünlükara, A. The effects of water salinity and potassium levels on yield, fruit quality and water consumption of a native central anatolian tomato species (Lycopersicon esculantum). Agric. Water Manage. 2005, 78, 128–135. [Google Scholar] [CrossRef]
- Kleiber, T. Nutritional resources of soil in the localities of monumental large-leaved linden (Tilia platyphyllos f. aurea) alleys. Ecol. Chem. Eng. 2009, 16, 277–286. [Google Scholar]
- Gałuszka, A.; Migaszewski, Z.M.; Podlaski, R.; Dołęgowska, S.; Michalik, A. The influence of chloride deicers on mineral nutrition and the health status of roadside trees in the city of Kielce, Poland. Environ. Monit. Assess. 2011, 176, 451–464. [Google Scholar] [CrossRef] [Green Version]
- Wilkaniec, B.; Breś, W.; Frużyńska-Jóźwiak, D.; Borowiak-Sobkowiak, B.; Wilkaniec, A. The assessment of chemical properties of soil, the chemical composition of leaves and the occurrence of diseases on Acer platanoides and Tilia cordata in selected sites of urban greenery in Poznań. Phytopathology 2012, 65, 19–28. [Google Scholar]
- Breś, W.; Kozłowska, M.; Kupska, A. The salinity of soils located along the selected streets of Poznan. Current trends in the horticultural plants cultivation. In National Scientific Conference; Department of Soil Cultivation and Horticultural Plant Fertilization: Lublin-Susiec, Poland, 2014; p. 40. (In Polish) [Google Scholar]
- Equiza, M.A.; Calvo-Polanco, M.; Cirelli, D.; Señorans, J.; Wartenbe, M.; Saunders, C.; Zwiazek, J.J. Long-term impact of road salt (NaCl) on soil and urban trees in Edmonton, Canada. Urban For. Urban Green. 2017, 21, 16–28. [Google Scholar] [CrossRef]
- Kotuby-Amacher, J.; Koenig, R.; Kitchen, B. Salinity and Plant Tolerance 2000, All Archived Publications. Paper 43. Available online: https://digitalcommons.usu.edu/extension_histall/43 (accessed on 15 December 2022).
- Maas, E.V.; Hoffman, G.J. Crop salt tolerance-current assessment. ASCE J. Irrig. Drain. Divsion 1977, 10, 115–134. [Google Scholar] [CrossRef]
- Miyamoto, S.; Martinez, I.; Portillo, M.; Ornelas, A. Landscape Plant Lists for Salt Tolerance Assessment. USDI Bureau of Reclamation. Texas Agricultural Extension Station, El Paso. 2004. Available online: http://agrilife.org/elpaso/files/2011/10/Landscape-Plant-Lists-for-Salt-Tolerance-Assessment.pdf (accessed on 15 December 2022).
- Cosgrove, D.J. Expansive growth of plant cell walls. Plant Physiol. Biochem. 2000, 38, 109–124. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of Plant Responses to Salt Stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef]
- Xie, M.; Zhang, J.; Tschaplinski, T.J.; Tuskan, G.A.; Chen, J.-G.; Muchero, W. Regulation of Lignin Biosynthesis and Its Role in Growth-Defense Tradeoffs. Front. Plant Sci. 2018, 9, 1427. [Google Scholar] [CrossRef] [Green Version]
- Neuman, P.M. The role of cel wall adjustment in plant resistance to watre deficyt. Crop. Sci. 1995, 35, 1258–1266. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, W.; Long, S.; Zhao, C. Maintenance of CellWall Integrity under High Salinity. Int. J. Mol. Sci. 2021, 22, 3260. [Google Scholar] [CrossRef]
- Oliveira, D.M.; Mota, T.R.; Salatta, F.V.; Sinzker, R.C.; Končitíková, R.; Kopečný, D.; Simister, R.; Silva, M.; Goeminne, G.; Morreel, K.; et al. Cell wall remodeling under salt stress: Insights into changes in polysaccharides, feruloylation, lignification, and phenolic metabolism in maize. Plant Cell Environ. 2020, 43, 2172–2191. [Google Scholar] [CrossRef]
- Tavakkoli, E.; Fatehi, F.; Coventry, S.; Rengasamy, P.; McDonald, G.K. Additive effects of Na+ and Cl− ions on barley growth under salinity stress. J. Exp. Bot. 2011, 62, 2189–2203. [Google Scholar] [CrossRef] [Green Version]
- Wakeel, A. Potassium-sodium interactions in soil and plant under saline-sodic conditions. J. Plant Nutr. Soil Sci. 2013, 176, 344–354. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The Critical Role of Potassium in Plant Stress Response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [Green Version]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Per, T.S.; Khan, N.A.; Reddy, P.S.; Masood, A.; Hasanuzzamn, M.; Khan, I.R.; Ajum, N.A. Approaches in modeling proline metabolism in plants for salt and drought tolerance: Phytohormones, mineral nutrients and transgenics. Plant Physiol. Biochem. 2017, 115, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Trovato, M.; Forlani, G.; Signorelli, S.; Funck, D. Proline Metabolism and Its Functions in Development and Stress Tolerance. In Osmoprotectant-Mediated Abiotic Stress Tolerance in Plants; Springer Nature: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]
- Moukhtari, A.E.; Cabass-Hourton, C.C.; Farissi, M.; Savouré, A. How does proline treatment promote salt stress tolerance during crop plant development? Front. Plant Sci. 2020, 11, 1127. [Google Scholar] [CrossRef] [PubMed]
- Bhaskara, G.B.; Yang, T.H.; Verslues, P.E. Dynamic proline metabolism: Importance and regulation in water limited environments. Front. Plant Sci. 2015, 6, 484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Zhao, L.; Chen, D.; Liang, M.; Liu, Z.; Shao, H.; Long, X. Salt Stress Encourages Proline Accumulation by Regulating Proline Biosynthesis and Degradation in Jerusalem Artichoke Plantlets. PLoS ONE 2013, 8, e62085. [Google Scholar] [CrossRef]
- Li, W.; Meng, R.; Liu, Y.; Chen, S.; Jiang, J.; Wang, L.; Zhao, S.; Wang, Z.; Fang, W.; Chen, F.; et al. Heterografted chrysanthemums enhance salt stress tolerance by integrating reactive oxygen species, soluble sugar, and proline. Hort. Res. 2022, 9, uhac073. [Google Scholar] [CrossRef]
- Zdunek-Zastocka, E.; Grabowska, A.; Michniewska, B.; Orzechowski, S. Proline Concentration and Its Metabolism Are Regulated in a Leaf Age Dependent Manner But Not by Abscisic Acid in Pea Plants Exposed to Cadmium Stress. Cells 2021, 10, 946. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Agarwal, R.; Tomar, N.S.; Shrivastava, M. Potassium induces positive changes in nitrogen metabolism and antioxidant system of oat (Avena sativa L. cultivar Kent). J. Plant Inter. 2015, 10, 211–223. [Google Scholar] [CrossRef]
- Sulandjari, A.; Sakya, T.; Wijayanti, R.N. Salinity and potassium fertilizer on growth and proline of the medicinal plant Pereskia bleo. In IOP Conference Series: Earth and Environmental Science, Proceedings of the 6th International Conference on Climate Change, 25 May 2021, Surakarta, Indonesia; IOP Publishing Ltd.: Bristol, UK, 2021; Volume 824. [Google Scholar] [CrossRef]
- Heidari, M.; Jamshidi. Effects of Salinity and Potassium Application on Antioxidant Enzyme Activities and Physiological Parameters in Pearl Millet. Agric. Sci. China 2011, 10, 228–237. [Google Scholar] [CrossRef]
- Rao, R.C.N.; Krishnasastry, K.S.; Udayakumar, M. Role of potassium in proline metabolism. I. Conversion of precursors into proline under stress conditions in K-sufficient and K-deficient plants. Plant Sci. Lett. 1981, 23, 327–334. [Google Scholar] [CrossRef]
- Kalra, Y.P. Handbook of Reference Methods for Plant Analysis; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 1998; p. 287. [Google Scholar]
- Weatherly, P.E. Studies in water relation of cotton plants. The measurement of water deficits in leaves. New Phytol. 1950, 49, 81–97. [Google Scholar] [CrossRef]
- Bandurska, H. Does proline accumulated in leaves of water deficit stressed barley plants confine cell membrane injury? I. Free proline accumulation and membrane injury index in drought and osmotic stressed plants. Acta Physiol. Plantarum 2000, 22, 409–415. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, J.D. Rapid determination of proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Zhang, C.-S.; Lu, Q.; Verma, D.P.S. Removal of feedback inhibition of Δ1-pyrroline-carboxylate synthetase, a bifunctional enzyme catalyzing the first two steps of proline biosynthesis in plants. J. Biol. Chem. 1995, 270, 20491–20496. [Google Scholar] [CrossRef] [Green Version]
- Rahnama, H.; Ebrahimzadeh, H. The effect of NaCl on proline accumulation in potato seedlings and calli. Acta Physiol. Plant. 2004, 26, 263–270. [Google Scholar] [CrossRef]
- Bradford, M.M.A. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Hiscox, J.C.; Israelstam, G.F. A method for the extraction of chlorophyll from tissue without maceration. Can. J. Bot. 1979, 57, 1332–1334. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Syros, T.; Yupsanis, T.; Zafiriadis, H.; Economou, A. Activity and isoforms of peroxidases, lignin and anatomy, during adventitious rooting in cuttings of Ebenus cretica L. J. Plant Physiol. 2004, 161, 69–77. [Google Scholar] [CrossRef]
- Premachandra, G.S.; Saneoka, G.S.; Ogata, H. Cell membrane stability, as indicator of drought tolerance, as affected by applied nitrogen and soybean. J. Agric. Sci. 1990, 115, 63–66. [Google Scholar] [CrossRef]
- Sakia, R.M. The Box-Cox Transformation Technique: A Review. J. R. Statist. Soc. Ser. D 1992, 41, 169–178. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Df | Main Shoot Height | Leaf Area | Aboveground Part Fresh Weight | Inflorescence Diameter | ||||
|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | ||
| NaCl | 6 | 24.369 | 0.0000 | 95.89 | 0.0000 | 130.435 | 0.0000 | 41.89 | 0.0000 |
| Treatments | Df | Water Loss | Df | RWC | ||
|---|---|---|---|---|---|---|
| F | p | F | p | |||
| Potassium | 1 | 13.21 | 0.0003 | 1 | 5.468 | 0.0221 |
| NaCl | 2 | 1381.32 | 0.0000 | 2 | 133.083 | 0.0000 |
| Date | 2 | 399.73 | 0.0000 | 2 | 69.872 | 0.0000 |
| Date × potassium | 25 | 5.86 | 0.0000 | 2 | 1.889 | 0.1587 |
| NaCl × potassium | 25 | 8.11 | 0.0003 | 2 | 0.925 | 0.4012 |
| NaCl × date | 50 | 12.97 | 0.0000 | 4 | 2.236 | 0.0735 |
| NaCl × date × potassium | 50 | 2.90 | 0.0000 | 4 | 0.977 | 0.4256 |
| Treatments | Df | K | Na | Cl | Na/K | ||||
|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | ||
| NaCl | 2 | 25.911 | 0.0000 | 102.867 | 0.0000 | 360.356 | 0.0000 | 200.123 | 0.0000 |
| Potassium | 1 | 11.509 | 0.0020 | 9.628 | 0.0002 | 8.597 | 0.0064 | 16.122 | 0.0004 |
| NaCl × potassium | 2 | 0.243 | 0.7856 | 7.276 | 0.0007 | 1.477 | 0.2444 | 0.288 | 0.7521 |
| Treatments | Df | Main Shoot Height | Lateral Shoot Number | Leaf Area | |||
| F | p | F | p | F | p | ||
| NaCl | 2 | 145.342 | 0.0000 | 7.9817 | 0.0022 | 385.693 | 0.0000 |
| Potassium | 1 | 29.244 | 0.0000 | 13.1460 | 0.0013 | 55.3951 | 0.0000 |
| NaCl × potassium | 2 | 5.9135 | 0.0082 | 0.3426 | 0.7133 | 11.8915 | 0.0003 |
| Treatments | Df | Aboveground Part Fresh Weight | Inflorescence Diameter | Number of Inflorescences | |||
| F | p | F | p | F | p | ||
| NaCl | 180.543 | 0.0000 | 13.779 | 0.0001 | 17.996 | 0.0000 | |
| Potassium | 3.121 | 0.0900 | 13.390 | 0.0012 | 3.378 | 0.0785 | |
| NaCl × potassium | 0.4259 | 0.7224 | 0.848 | 0.4408 | 1.4145 | 0.2626 | |
| Treatments | Df | Chlorophyll | Lignin | Membrane Injury | |||
|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | ||
| NaCl | 2 | 4.859 | 0.0169 | 19.683 | 0.0000 | 65.818 | 0.0000 |
| Potassium | 1 | 1.323 | 0.2615 | 40.929 | 0.0000 | 9.187 | 0.0080 |
| NaCl × potassium | 1 | 0.871 | 0.4313 | 1.877 | 0.1749 | 0.766 | 0.3943 |
| Treatments | Df | Proline | P5CS | PDH | |||
|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | ||
| NaCl | 2 | 129.330 | 0.0000 | 40.582 | 0.0000 | 191.364 | 0.0000 |
| Date | 2 | 69.634 | 0.0000 | 15.330 | 0.0000 | 17.461 | 0.0000 |
| Potassium | 1 | 32.680 | 0.0000 | 1.270 | 0.2635 | 6.218 | 0.0149 |
| NaCl × date | 4 | 28.069 | 0.0000 | 4.043 | 0.0052 | 3.866 | 0.0067 |
| NaCl × potassium | 2 | 41.251 | 0.0000 | 0.805 | 0.4512 | 10.916 | 0.0000 |
| Date × potassium | 2 | 1.689 | 0.1920 | 0.233 | 0.7930 | 0.974 | 0.3826 |
| NaCl × date × potassium | 4 | 9.329 | 0.0000 | 0.441 | 0.7787 | 11.756 | 0.0000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bandurska, H.; Breś, W.; Zielezińska, M.; Mieloszyk, E. Does Potassium Modify the Response of Zinnia (Zinnia elegans Jacq.) to Long-Term Salinity? Plants 2023, 12, 1439. https://doi.org/10.3390/plants12071439
Bandurska H, Breś W, Zielezińska M, Mieloszyk E. Does Potassium Modify the Response of Zinnia (Zinnia elegans Jacq.) to Long-Term Salinity? Plants. 2023; 12(7):1439. https://doi.org/10.3390/plants12071439
Chicago/Turabian StyleBandurska, Hanna, Włodzimierz Breś, Małgorzata Zielezińska, and Elżbieta Mieloszyk. 2023. "Does Potassium Modify the Response of Zinnia (Zinnia elegans Jacq.) to Long-Term Salinity?" Plants 12, no. 7: 1439. https://doi.org/10.3390/plants12071439
APA StyleBandurska, H., Breś, W., Zielezińska, M., & Mieloszyk, E. (2023). Does Potassium Modify the Response of Zinnia (Zinnia elegans Jacq.) to Long-Term Salinity? Plants, 12(7), 1439. https://doi.org/10.3390/plants12071439