
Gamma-Aminobutyric Acid Enhances Cadmium Phytoextraction by Coreopsis grandiflora by Remodeling the Rhizospheric Environment
 and
and
Abstract
:
1. Introduction
2. Results
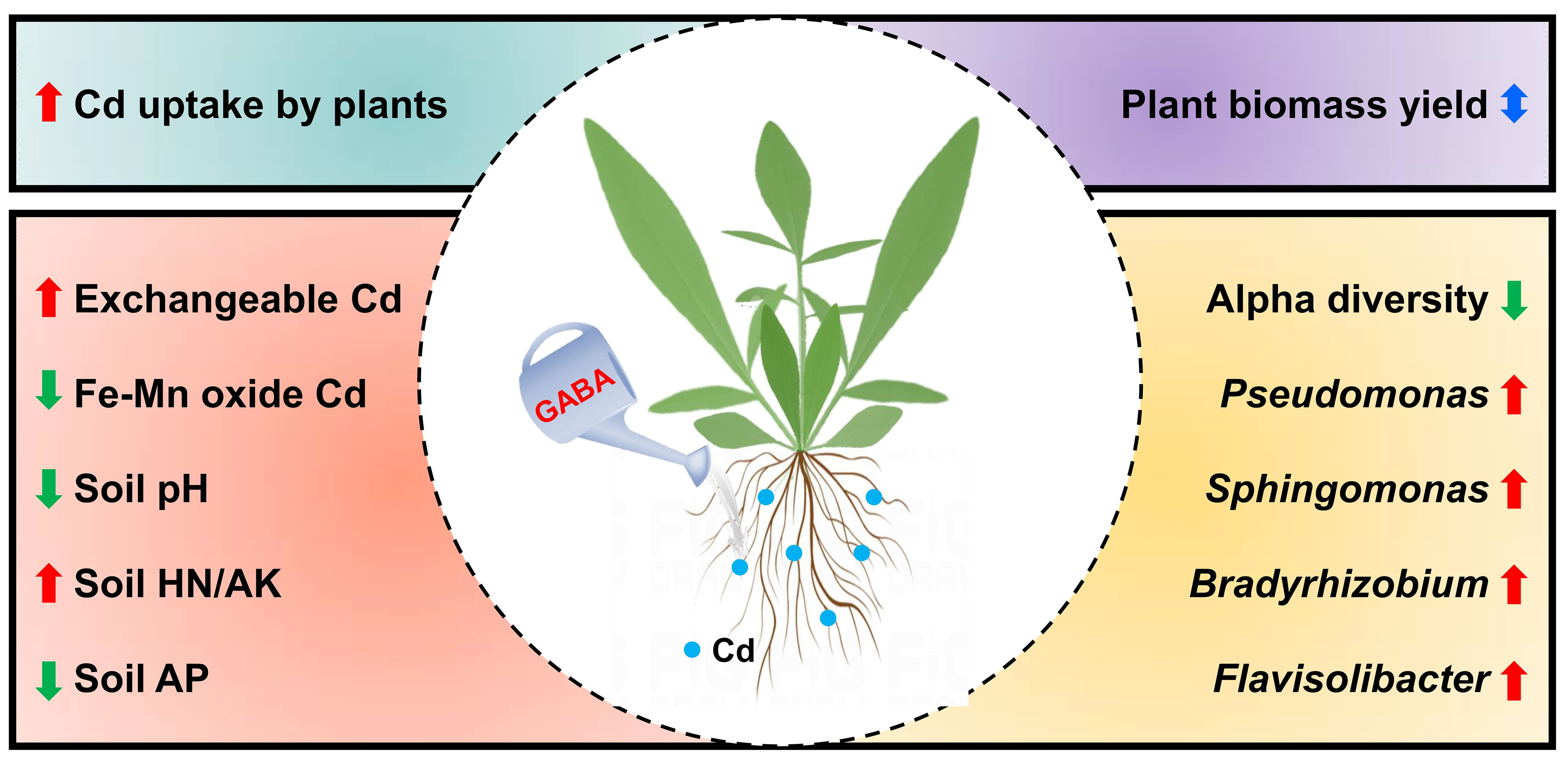
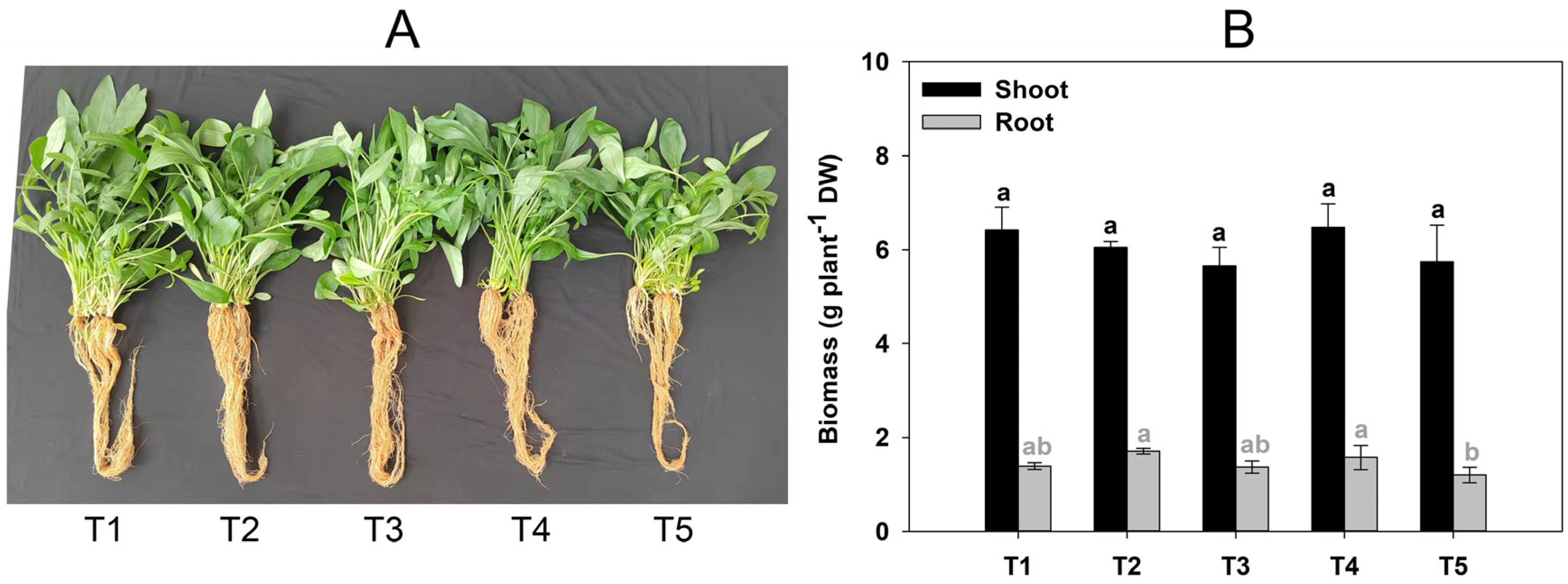
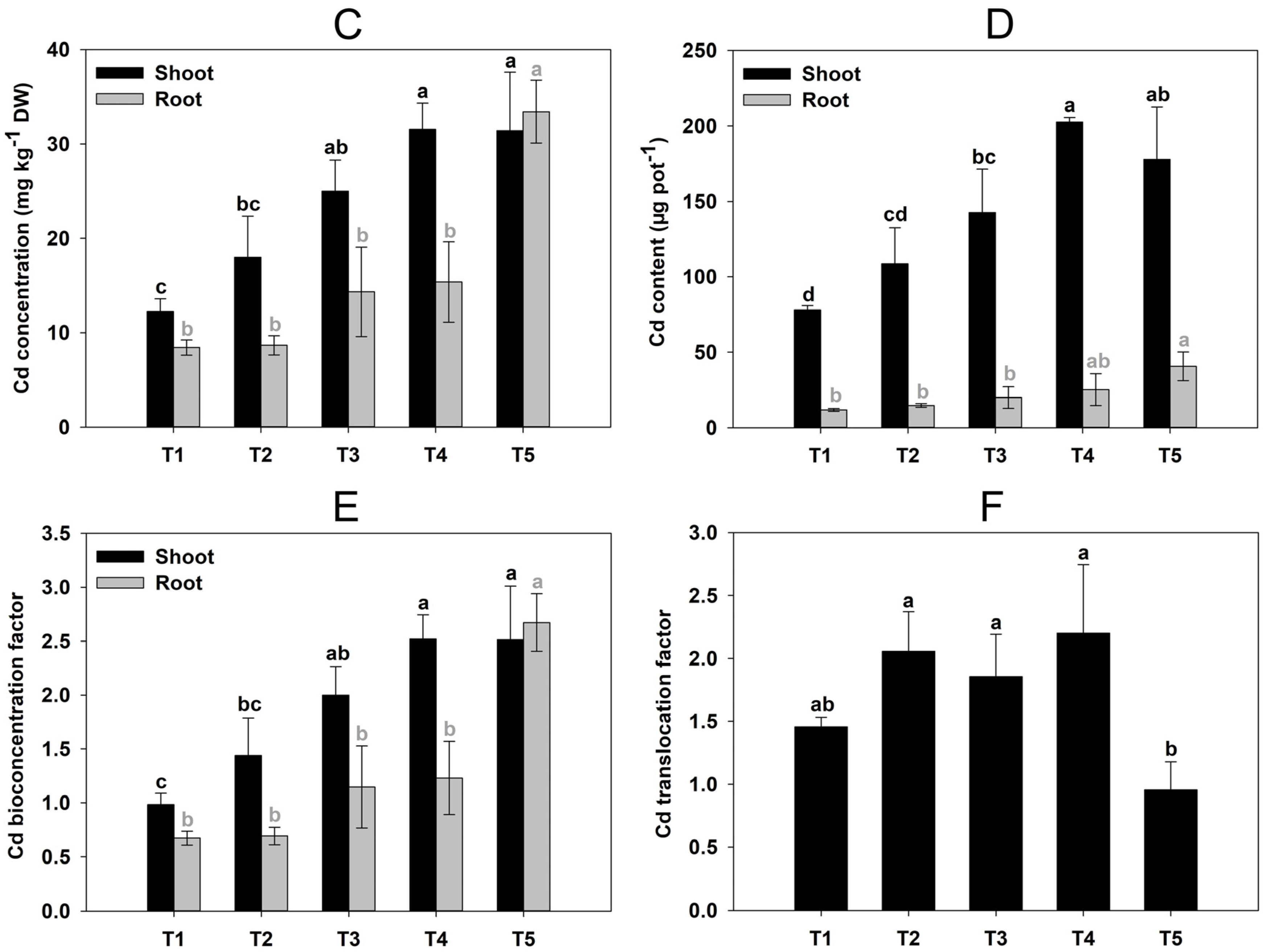
2.1. Effects of GABA Addition on the Growth and Cd Uptake of C. grandiflora
2.2. Effects of GABA Addition on Cd Bioavailability in the C. grandiflora Rhizosphere
2.3. Leaching of Cd in Soils by GABA
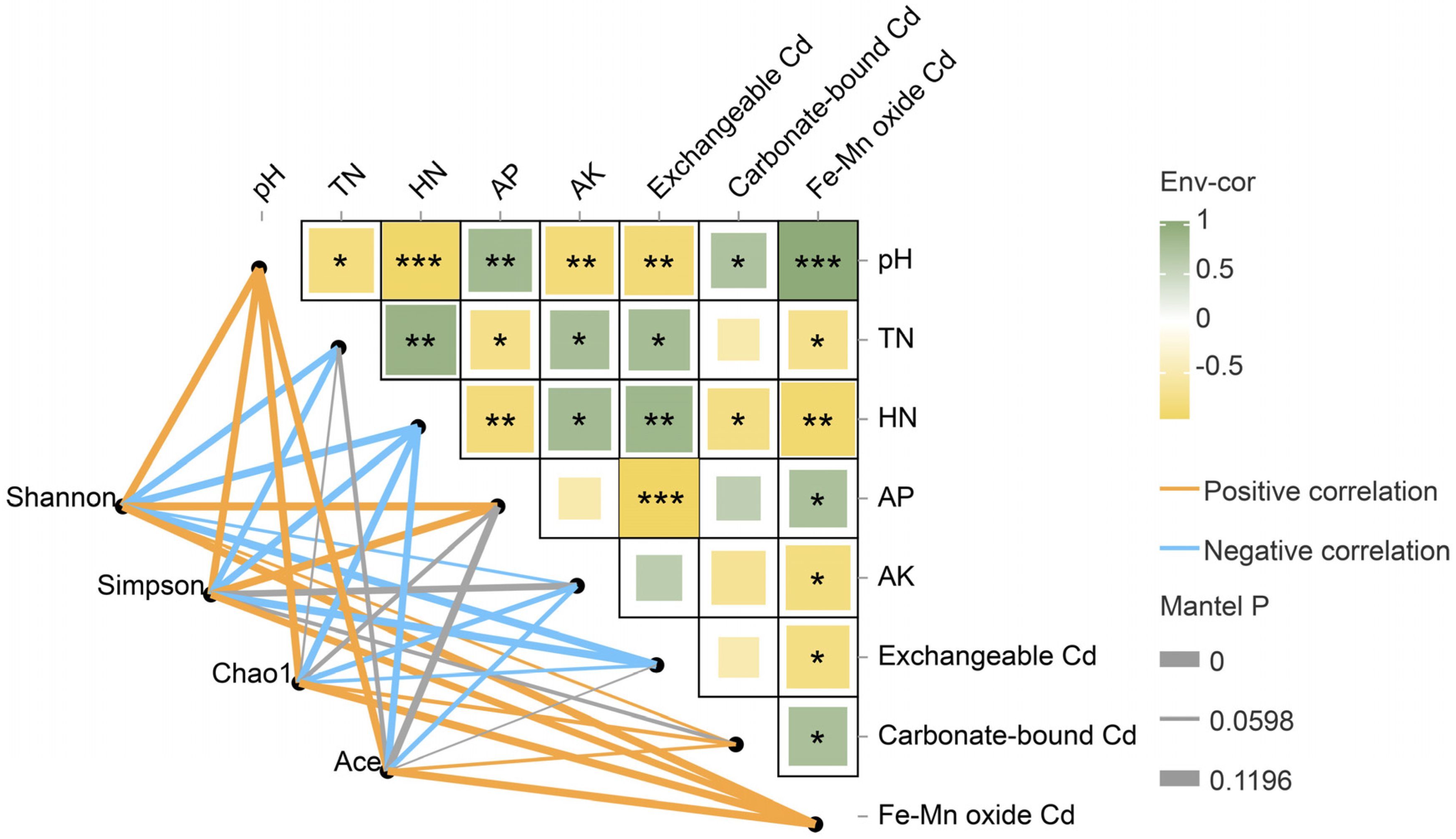
2.4. Effects of GABA Addition on Physicochemical Indices in the C. grandiflora Rhizosphere
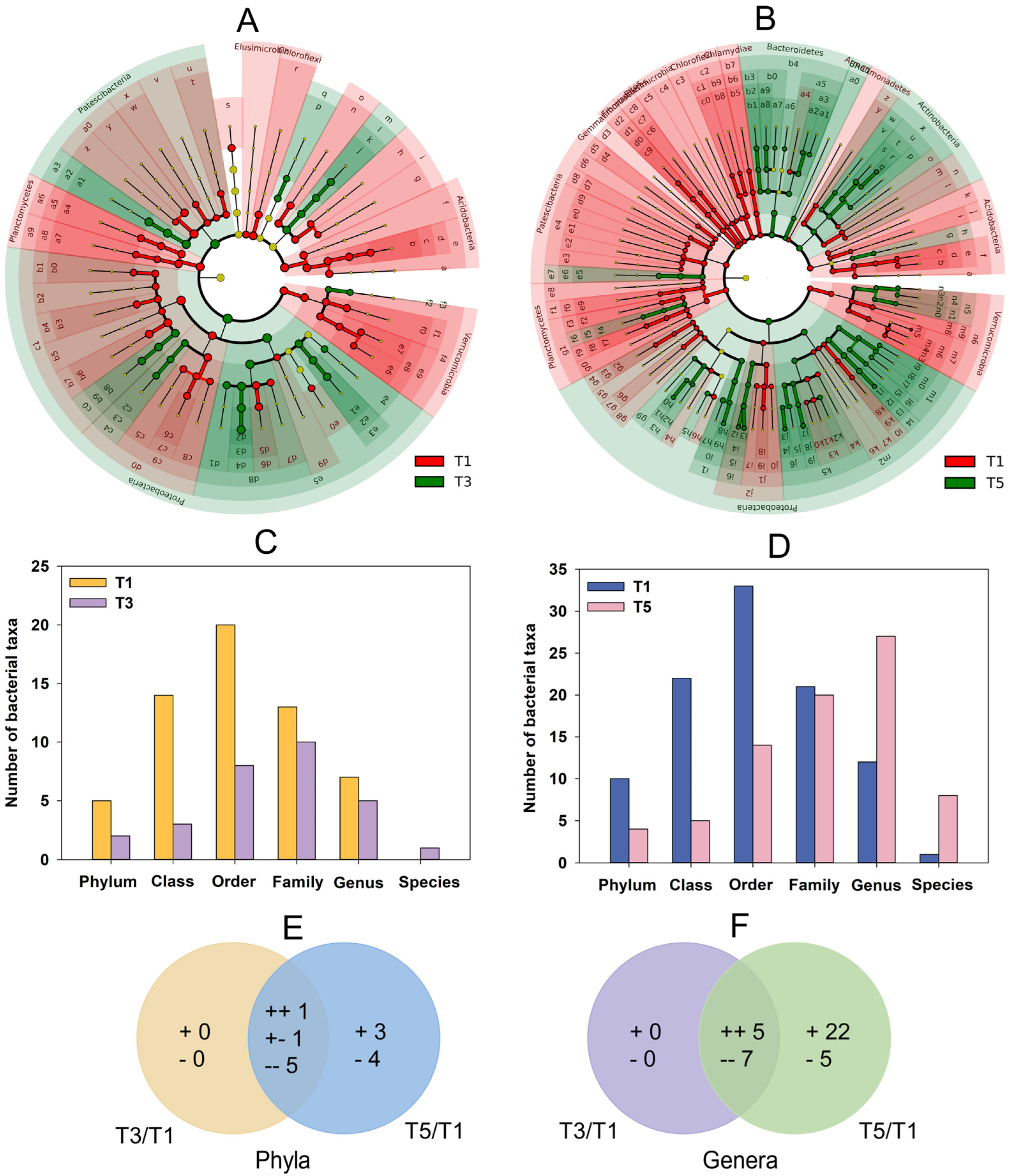
2.5. Bacterial Community Composition in the C. grandiflora Rhizosphere
2.5.1. Composition of Bacterial Communities
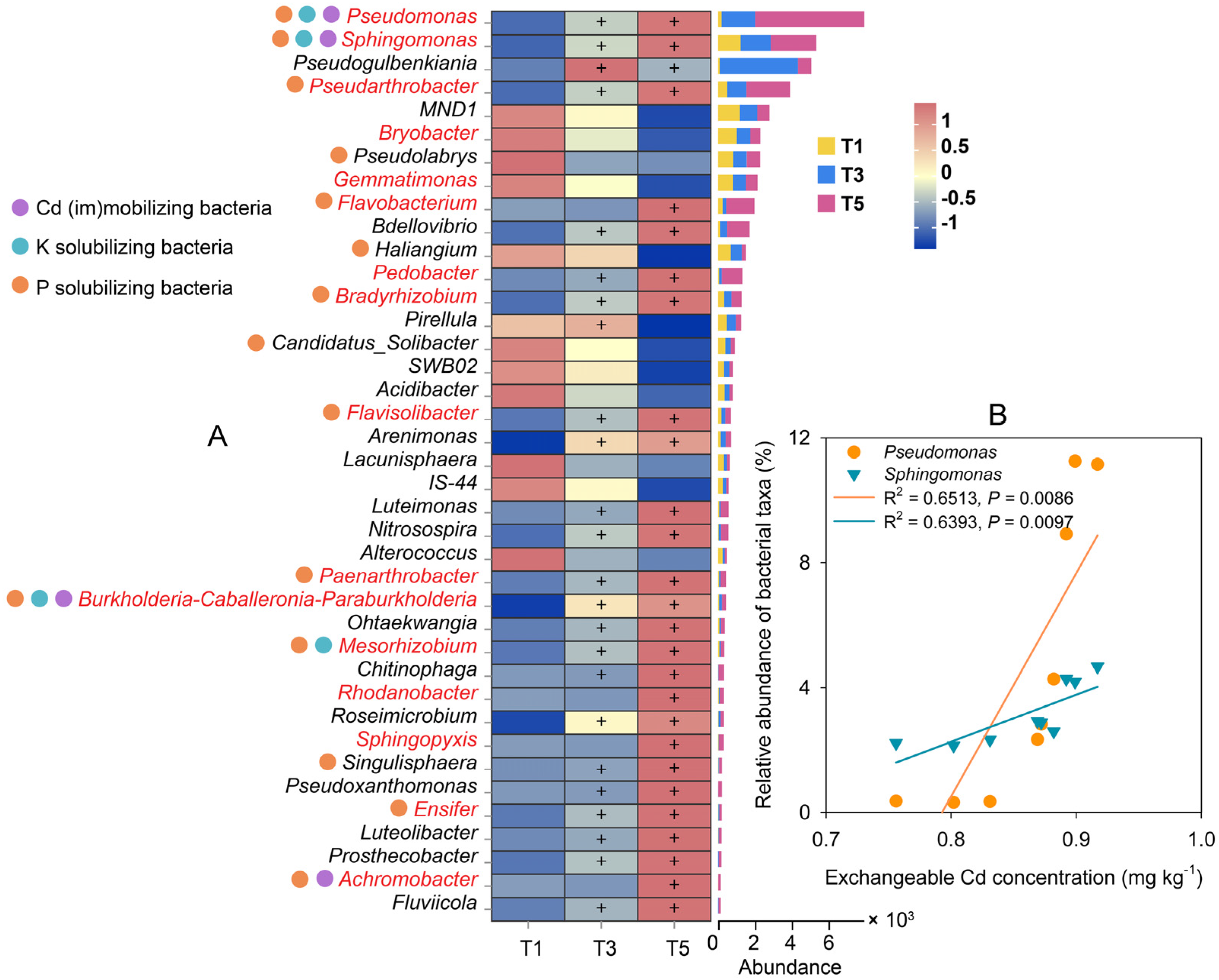
2.5.2. Variations in Rhizobacteria under GABA Addition
3. Discussion
4. Materials and Methods
4.1. Experimental Treatment
4.2. Sample Collection and Biomass Measurement
4.3. Cd Accumulation Characteristics Analysis in Plants
4.4. Cd Speciation Determination in Soil
4.5. Leaching Experiment
4.6. Determination of Soil Physicochemical Indices
4.7. Soil Bacterial Community Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Briffa, J.; Sinagra, E.; Blundell, R. Heavy metal pollution in the environment and their toxicological effects on humans. Heliyon 2020, 6, e04691. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Prasad, S.; Yadav, K.K.; Shrivastava, M.; Gupta, N.; Nagar, S.; Bach, Q.V.; Kamyab, H.; Khan, S.A.; Yadav, S.; et al. Hazardous heavy metals contamination of vegetables and food chain: Role of sustainable remediation approaches-A review. Environ. Res. 2019, 179, 108792. [Google Scholar] [CrossRef] [PubMed]
- Yan, A.; Wang, Y.M.; Tan, S.N.; Yusof, M.L.M.; Ghosh, S.; Chen, Z. Phytoremediation: A promising approach for revegetation of heavy metal-polluted land. Front. Plant. Sci. 2020, 11, 359. [Google Scholar] [CrossRef]
- Yuan, C.; Gao, B.; Peng, Y.; Gao, X.; Fan, B.; Chen, Q. A meta-analysis of heavy metal bioavailability response to biochar aging: Importance of soil and biochar properties. Sci. Total Environ. 2021, 756, 144058. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.Y.; Chen, T.B.; Yan, X.L.; NIe, C.J. Enhancement of heavy metal removal in phytoremediation of soils contaminated with heavy metals. Acta Sci. Circum. 2007, 27, 882–893. [Google Scholar]
- Palansooriya, K.N.; Shaheen, S.M.; Chen, S.S.; Tsang, D.C.W.; Hashimoto, Y.; Hou, D.Y.; Bolan, N.S.; Rinklebe, J.; Ok, Y.S. Soil amendments for immobilization of potentially toxic elements in contaminated soils: A critical review. Environ. Int. 2020, 134, 105046. [Google Scholar] [CrossRef] [PubMed]
- Gul, I.; Manzoor, M.; Hashim, N.; Shah, G.M.; Waani, S.P.T.; Shahid, M.; Antoniadis, V.; Rinklebe, J.; Arshad, M. Challenges in microbially and chelate-assisted phytoextraction of cadmium and lead—A review. Environ. Pollut. 2021, 287, 117667. [Google Scholar] [CrossRef]
- He, X.M.; Zhang, J.; Ren, Y.N.; Sun, C.Y.; Deng, X.P.; Qian, M.; Hu, Z.B.; Li, R.; Chen, Y.H.; Shen, Z.G.; et al. Polyaspartate and liquid amino acid fertilizer are appropriate alternatives for promoting the phytoextraction of cadmium and lead in Solanum nigrum L. Chemosphere 2019, 237, 124483. [Google Scholar] [CrossRef]
- Dolev, N.; Katz, Z.; Ludmer, Z.; Ullmann, A.; Brauner, N.; Goikhman, R. Natural amino acids as potential chelators for soil remediation. Environ. Res. 2020, 183, 109140. [Google Scholar] [CrossRef]
- Li, X.; Tian, L.Y.; Li, B.Q.; Chen, H.F.; Zhao, G.J.; Qin, X.S.; Liu, Y.Y.; Yang, Y.P.; Xu, J.C. Polyaspartic acid enhances the Cd phytoextraction efficiency of Bidens pilosa by remolding the rhizospheric environment and reprogramming plant metabolism. Chemosphere 2022, 307, 136068. [Google Scholar] [CrossRef]
- Luo, H.Z.; Liu, Z.; Xie, F.; Bilal, M.; Liu, L.N.; Yang, R.L.; Wang, Z.Y. Microbial production of gamma-aminobutyric acid: Applications, state-of-the-art achievements, and future perspectives. Crit. Rev. Biotechnol. 2021, 41, 491–512. [Google Scholar] [CrossRef] [PubMed]
- Bor, M.; Seckin, B.; Ozgur, R.; Yilmaz, O.; Ozdemir, F.; Turkan, I. Comparative effects of drought, salt, heavy metal and heat stresses on gamma-aminobutryric acid levels of sesame (Sesamum indicum L.). Acta Physiol. Plant 2009, 31, 655–659. [Google Scholar] [CrossRef]
- Kumar, N.; Gautam, A.; Dubey, A.K.; Ranjan, R.; Pandey, A.; Kumari, B.; Singh, G.; Mandotra, S.; Chauhan, P.S.; Srikrishna, S.; et al. GABA mediated reduction of arsenite toxicity in rice seedling through modulation of fatty acids, stress responsive amino acids and polyamines biosynthesis. Ecotox. Environ. Safe 2019, 173, 15–27. [Google Scholar] [CrossRef]
- Li, Y.; Li, Y.; Cui, Y.; Xie, Y.; Shi, Y.; Shang, Y.; Ma, F.; Zhang, J.; Li, C. GABA-mediated inhibition of cadmium uptake and accumulation in apples. Environ. Pollut. 2022, 300, 118867. [Google Scholar] [CrossRef]
- Mahmud, J.A.L.; Hasanuzzaman, M.; Nahar, K.; Rahman, A.; Hossain, M.S.; Fujita, M. gamma-aminobutyric acid (GABA) confers chromium stress tolerance in Brassica juncea L. by modulating the antioxidant defense and glyoxalase systems. Ecotoxicology 2017, 26, 675–690. [Google Scholar] [CrossRef]
- Seifikalhor, M.; Aliniaeifard, S.; Bernard, F.; Seif, M.; Latifi, M.; Hassani, B.; Didaran, F.; Bosacchi, M.; Rezadoost, H.; Li, T. gamma-Aminobutyric acid confers cadmium tolerance in maize plants by concerted regulation of polyamine metabolism and antioxidant defense systems. Sci. Rep. 2020, 10, 3356. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Li, B.; Zheng, Y.; Luo, L.; Qin, X.; Yang, Y.; Xu, J. Physiological and rhizospheric response characteristics to cadmium of a newly identified cadmium accumulator Coreopsis grandiflora Hogg. (Asteraceae). Ecotox. Environ. Safe 2022, 241, 113739. [Google Scholar] [CrossRef]
- He, G.Q.; Zhang, H.B.; Liu, S.Q.; Li, H.Q.; Huo, Y.Z.; Guo, K.W.; Xu, Z.S.; Zhang, H.H. Exogenous γ-glutamic acid (GABA) induces proline and glutathione synthesis in alleviating Cd-induced photosynthetic inhibition and oxidative damage in tobacco leaves. J. Plant Interact. 2021, 16, 296–306. [Google Scholar] [CrossRef]
- Yang, C.J.; Wei, S.H.; Zhou, Q.X.; Zhang, L.; Bao, Y.Y.; Gu, P.; Ren, L.P.; Liu, R. Promotion effects of exogenous amino acids phytoremediation of Cd-PAHs contaminated soils by using hyperaccumulators plant Solanum nigrum. Chin. J. Ecol. 2009, 28, 1829–1834. [Google Scholar]
- Kim, Y.O.; Gwon, Y.; Kim, J. Exogenous cysteine improves mercury uptake and tolerance in Arabidopsis by regulating the expression of heavy metal chelators and antioxidative enzymes. Front. Plant Sci. 2022, 13, 898247. [Google Scholar] [CrossRef]
- Yildiz, M.; Terzi, H. Exogenous cysteine alleviates chromium stress via reducing its uptake and regulating proteome in roots of Brassica napus L. seedlings. S. Afr. J. Bot. 2021, 139, 114–121. [Google Scholar] [CrossRef]
- Terzi, H.; Yildiz, M. Proteomic analysis reveals the role of exogenous cysteine in alleviating chromium stress in maize seedlings. Ecotox. Environ. Safe 2021, 209, 111784. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Dai, M.; Yang, J.; Sun, L.; Tan, X.; Peng, C.; Ali, I.; Naz, I. A critical review on the phytoremediation of heavy metals from environment: Performance and challenges. Chemosphere 2022, 291, 132979. [Google Scholar] [CrossRef] [PubMed]
- Tessier, A.; Campbell, P.G.C.; Bisson, M. Sequential extraction procedure for the speciation of particulate trace-metals. Anal. Chem. 1979, 51, 844–851. [Google Scholar] [CrossRef]
- Kaurin, A.; Gluhar, S.; Tilikj, N.; Lestan, D. Soil washing with biodegradable chelating agents and EDTA: Effect on soil properties and plant growth. Chemosphere 2020, 260, 127673. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Liu, Y. Effects of single and combined organic acids on form of Cd in soil. J. Soil Sci. 2012, 43, 186–189. [Google Scholar]
- Song, X.F. The Effects of Different Fertilization on Cadmium Accumulation and Transference in Soil. Master’ Dissertation, Shenyang Agricultural University, Shenyang, China, 2018. [Google Scholar]
- Cerozi, B.D.S.; Fitzsimmons, K. The effect of pH on phosphorus availability and speciation in an aquaponics nutrient solution. Bioresource Technol. 2016, 219, 778–781. [Google Scholar] [CrossRef] [Green Version]
- Xiang, J.; Haden, V.R.; Peng, S.B.; Bouman, B.A.M.; Visperas, R.M.; Nie, L.X.; Huang, J.L.; Cui, K.H. Improvement in nitrogen availability, nitrogen uptake and growth of aerobic rice following soil acidification. Soil Sci. Plant Nutr. 2009, 55, 705–714. [Google Scholar] [CrossRef]
- Chen, C.C.; Xiao, B.; Yu, Y.B.; Gong, X.F. Spatial variability of soil organic matter and pH and the correlation to available nutrients in the tea garden of southern Shaanxi. J. Northwest A F Univ. 2009, 37, 182–188. [Google Scholar]
- Hadi, F.; Ahmad, A.; Ullah, R. Cadmium phytoextraction potential of Ricinus communis significantly increased with exogenous application of growth regulators and macronutrients. Soil Sediment Contam. 2021, 30, 663–685. [Google Scholar] [CrossRef]
- Chen, B.; Tan, S.D.; Zeng, Q.R.; Wang, A.D.; Zheng, H.B. Soil nutrient heterogeneity affects the accumulation and transfer of cadmium in Bermuda grass (Cynodon dactylon (L.) pers). Chemosphere 2019, 221, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, A.; Dennis, P.G.; Forstner, C.; Raghavendra, A.K.H.; Mathesius, U.; Richardson, A.E.; Delhaize, E.; Gilliham, M.; Watt, M.; Ryan, P.R. Manipulating exudate composition from root apices shapes the microbiome throughout the root system. Plant Physiol. 2021, 187, 2279–2295. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Lopes, L.D.; Lopez-Guerrero, M.G.; van Dijk, K.; Alvarez, S.; Riethoven, J.J.; Schachtman, D.P. Natural variation in root exudation of GABA and DIMBOA impacts the maize root endosphere and rhizosphere microbiomes. J. Exp. Bot. 2022, 73, 5052–5066. [Google Scholar] [CrossRef]
- Shen, Y.F.; E, Y.Y.; Yang, F.; Zhang, N.; Huang, Q.W.; Shen, Q.R. Effects of amino acids in root exudates of watermelon on the chemotactic reaction and root colonization of Paenibacillus polymyxa SQR-21. J. Nanjing Agric. Univ. 2017, 40, 101–108. [Google Scholar]
- Wan, W.J.; Tan, J.D.; Wang, Y.; Qin, Y.; He, H.M.; Wu, H.Q.; Zuo, W.L.; He, D.L. Responses of the rhizosphere bacterial community in acidic crop soil to pH: Changes in diversity, composition, interaction, and function. Sci. Total Environ. 2020, 700, 134418. [Google Scholar] [CrossRef]
- Liu, W.S.; Li, H.Q.; He, Y.; Huang, Y.Y.; Qiu, K.Y.; Xie, Y.Z. Research progress on the interaction regulation of plant and soil by rhizosphere microbiome. Soil Fertil. Sci. China 2021, 5, 318–327. [Google Scholar]
- Wang, W.Y.; Jia, T.H.; Qi, T.Y.; Li, S.S.; Degen, A.A.; Han, J.; Bai, Y.F.; Zhang, T.; Qi, S.; Huang, M.; et al. Root exudates enhanced rhizobacteria complexity and microbial carbon metabolism of toxic plants. Iscience 2022, 25, 105243. [Google Scholar] [CrossRef]
- Bhanse, P.; Kumar, M.; Singh, L.; Awasthi, M.K.; Qureshi, A. Role of plant growth-promoting rhizobacteria in boosting the phytoremediation of stressed soils: Opportunities, challenges, and prospects. Chemosphere 2022, 303, 134954. [Google Scholar] [CrossRef]
- Khatoon, Z.; Huang, S.; Rafique, M.; Fakhar, A.; Kamran, M.A.; Santoyo, G. Unlocking the potential of plant growth-promoting rhizobacteria on soil health and the sustainability of agricultural systems. J. Environ. Manag. 2020, 273, 111118. [Google Scholar] [CrossRef]
- Asaf, S.; Numan, M.; Khan, A.L.; Al-Harrasi, A. Sphingomonas: From diversity and genomics to functional role in environmental remediation and plant growth. Crit. Rev. Biotechnol. 2020, 40, 138–152. [Google Scholar] [CrossRef]
- Manoj, S.R.; Karthik, C.; Kadirvelu, K.; Arulselvi, P.I.; Shanmugasundaram, T.; Bruno, B.; Rajkumar, M. Understanding the molecular mechanisms for the enhanced phytoremediation of heavy metals through plant growth promoting rhizobacteria: A review. J. Environ. Manag. 2020, 254, 109779. [Google Scholar] [CrossRef]
- Sattar, A.; Naveed, M.; Ali, M.; Zahir, Z.A.; Nadeem, S.M.; Yaseen, M.; Meena, V.S.; Farooq, M.; Singh, R.; Rahman, M.; et al. Perspectives of potassium solubilizing microbes in sustainable food production system: A review. Appl. Soil. Ecol. 2019, 133, 146–159. [Google Scholar] [CrossRef]
- Rajkumar, M.; Bruno, L.B.; Banu, J.R. Alleviation of environmental stress in plants: The role of beneficial Pseudomonas spp. Crit. Rev. Env. Sci. Tec. 2017, 47, 372–407. [Google Scholar] [CrossRef]
- Shi, P.L.; Zhu, K.X.; Zhang, Y.X.; Chai, T.Y. Growth and cadmium accumulation of Solanum nigrum L. seedling were enhanced by heavy metal-tolerant strains of Pseudomonas aeruginosa. Water Air Soil Poll. 2016, 227, 459. [Google Scholar] [CrossRef]
- Chen, B.; Zhang, Y.; Rafiq, M.T.; Khan, K.Y.; Pan, F.; Yang, X.; Feng, Y. Improvement of cadmium uptake and accumulation in Sedum alfredii by endophytic bacteria Sphingomonas SaMR12: Effects on plant growth and root exudates. Chemosphere 2014, 117, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, Q.; Gao, H.; Zhang, R.; Yang, L.; Guo, Y.; Li, H.; Awasthi, M.K.; Li, G. Long-term cover crops improved soil phosphorus availability in a rain-fed apple orchard. Chemosphere 2021, 275, 130093. [Google Scholar] [CrossRef]
- Elhaissoufi, W.; Ghoulam, C.; Barakat, A.; Zeroual, Y.; Bargaz, A. Phosphate bacterial solubilization: A key rhizosphere driving force enabling higher P use efficiency and crop productivity. J. Adv. Res. 2022, 38, 13–28. [Google Scholar] [CrossRef]
- Li, X.; Chen, D.; Li, B.Q.; Yang, Y.P. Cd accumulation characteristics of Salvia tiliifolia and changes of rhizospheric soil enzyme activities and bacterial communities under a Cd concentration gradient. Plant Soil 2021, 463, 225–247. [Google Scholar] [CrossRef]
- Wu, M.X.; Luo, Q.; Liu, S.L.; Zhao, Y.; Long, Y.; Pan, Y.Z. Screening ornamental plants to identify potential Cd hyperaccumulators for bioremediation. Ecotox. Environ. Safe 2018, 162, 35–41. [Google Scholar] [CrossRef]
- Acosta, J.A.; Abbaspour, A.; Martinez, G.R.; Martinez-Martinez, S.; Zornoza, R.; Gabarron, M.; Faz, A. Phytoremediation of mine tailings with Atriplex halimus and organic/inorganic amendments: A five-year field case study. Chemosphere 2018, 204, 71–78. [Google Scholar] [CrossRef]
- Li, X.; Yang, Y.P. Preliminary study on Cd accumulation characteristics in Sansevieria trifasciata Prain. Plant Divers. 2020, 42, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Li, L.X. The Cd, Pb and Zn Removal from Contaminated Soils by Polyepoxysuccinic Acid Washing: Optimization and Improvement. Master’ Dissertation, Sichuan Agricultural University, Chengdu, China, 2018. [Google Scholar]
- Yang, Z.Y. Experimental Study on Remediation of Cd-Contaminated Soil by Surface Leaching & Deep Solidification. Master’ Dissertation, Southwest University, Chongqing, China, 2020. [Google Scholar]
- Gao, S.Q. Study on Leaching Remediation of Heavy Metal Contaminated Farmland Soil and Field Application. Master’ Dissertation, South China University of Technology, Guangzhou, China, 2020. [Google Scholar]
- Luo, L.Y.; Jiang, H.H.; Wang, D.D.; Zheng, L.M.; Zhang, D.Y.; Zeng, J.; Peng, J.C.; Zhang, Z. Effects of cadmium pollution on fungal community structure and diversity in paddy field. J. South. Agric. 2022, 53, 1908–1916. [Google Scholar]
- Zhang, Y.; Wu, C.F.; Zhang, J.L.; Fu, Z.C.; Wang, C. Effect of soil washing with different washing agents on the structure of microbial community in agricultural soil contaminated with cadmium. Acta Sci. Circum. 2022, 42, 398–404. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cd Chemsical Fractions | T1 | T3 | T5 | ANOVA | ||
|---|---|---|---|---|---|---|
| F Value | p Value | DF | ||||
| Total Cd | 12.26 ± 0.53 a | 12.40 ± 0.67 a | 11.97 ± 0.76 a | 0.224 | 0.805 | 2 |
| Exchangeable Cd | 0.80 ± 0.03 b | 0.87 ± 0.01 a | 0.90 ± 0.01 a | 16.610 | 0.004 | 2 |
| Carbonate-bound Cd | 0.07 ± 0.01 a | 0.07 ± 0.02 a | 0.04 ± 0.01 b | 3.452 | 0.101 | 2 |
| Fe-Mn oxide Cd | 0.25 ± 0.01 a | 0.24 ± 0.01 b | 0.18 ± 0.00 c | 90.799 | 0.000 | 2 |
| Organic-bound Cd | 0.04 ± 0.00 a | 0.04 ± 0.00 a | 0.03 ± 0.00 a | 0.853 | 0.472 | 2 |
| Residual Cd | 11.11 ± 0.54 a | 11.18 ± 0.67 a | 10.81 ± 0.76 a | 0.172 | 0.846 | 2 |
| Soil Indices | Unit | T1 | T3 | T5 | ANOVA | ||
|---|---|---|---|---|---|---|---|
| F Value | p Value | DF | |||||
| pH | / | 6.01 ± 0.06 a | 5.75 ± 0.02 b | 5.11 ± 0.04 c | 238.888 | 0.000 | 2 |
| OM | g kg−1 | 220.03 ± 8.49 a | 205.97 ± 11.61 a | 217.57 ± 14.13 a | 0.832 | 0.480 | 2 |
| TN | g kg−1 | 6.09 ± 0.21 b | 6.43 ± 0.28 ab | 6.77 ± 0.04 a | 5.617 | 0.042 | 2 |
| TP | g kg−1 | 1.41 ± 0.02 a | 1.44 ± 0.02 a | 1.41 ± 0.04 a | 0.672 | 0.545 | 2 |
| TK | g kg−1 | 9.38 ± 0.12 a | 8.82 ± 0.48 a | 9.02 ± 0.36 a | 1.276 | 0.345 | 2 |
| HN | mg kg−1 | 342.94 ± 20.13 b | 366.22 ± 7.33 b | 417.39 ± 6.34 a | 17.435 | 0.003 | 2 |
| AP | mg kg−1 | 29.56 ± 0.77 a | 25.85 ± 0.62 b | 25.06 ± 0.46 b | 29.329 | 0.001 | 2 |
| AK | mg kg−1 | 55.80 ± 2.67 b | 57.50 ± 3.83 b | 77.63 ± 10.47 a | 6.733 | 0.029 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Li, B.; Chen, H.; Li, J.; Xu, J.; Li, X. Gamma-Aminobutyric Acid Enhances Cadmium Phytoextraction by Coreopsis grandiflora by Remodeling the Rhizospheric Environment. Plants 2023, 12, 1484. https://doi.org/10.3390/plants12071484
Huang Y, Li B, Chen H, Li J, Xu J, Li X. Gamma-Aminobutyric Acid Enhances Cadmium Phytoextraction by Coreopsis grandiflora by Remodeling the Rhizospheric Environment. Plants. 2023; 12(7):1484. https://doi.org/10.3390/plants12071484
Chicago/Turabian StyleHuang, Yingqi, Boqun Li, Huafang Chen, Jingxian Li, Jianchu Xu, and Xiong Li. 2023. "Gamma-Aminobutyric Acid Enhances Cadmium Phytoextraction by Coreopsis grandiflora by Remodeling the Rhizospheric Environment" Plants 12, no. 7: 1484. https://doi.org/10.3390/plants12071484
APA StyleHuang, Y., Li, B., Chen, H., Li, J., Xu, J., & Li, X. (2023). Gamma-Aminobutyric Acid Enhances Cadmium Phytoextraction by Coreopsis grandiflora by Remodeling the Rhizospheric Environment. Plants, 12(7), 1484. https://doi.org/10.3390/plants12071484