
The Lush Fucales Underwater Forests off the Cilento Coast: An Overlooked Mediterranean Biodiversity Hotspot

 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
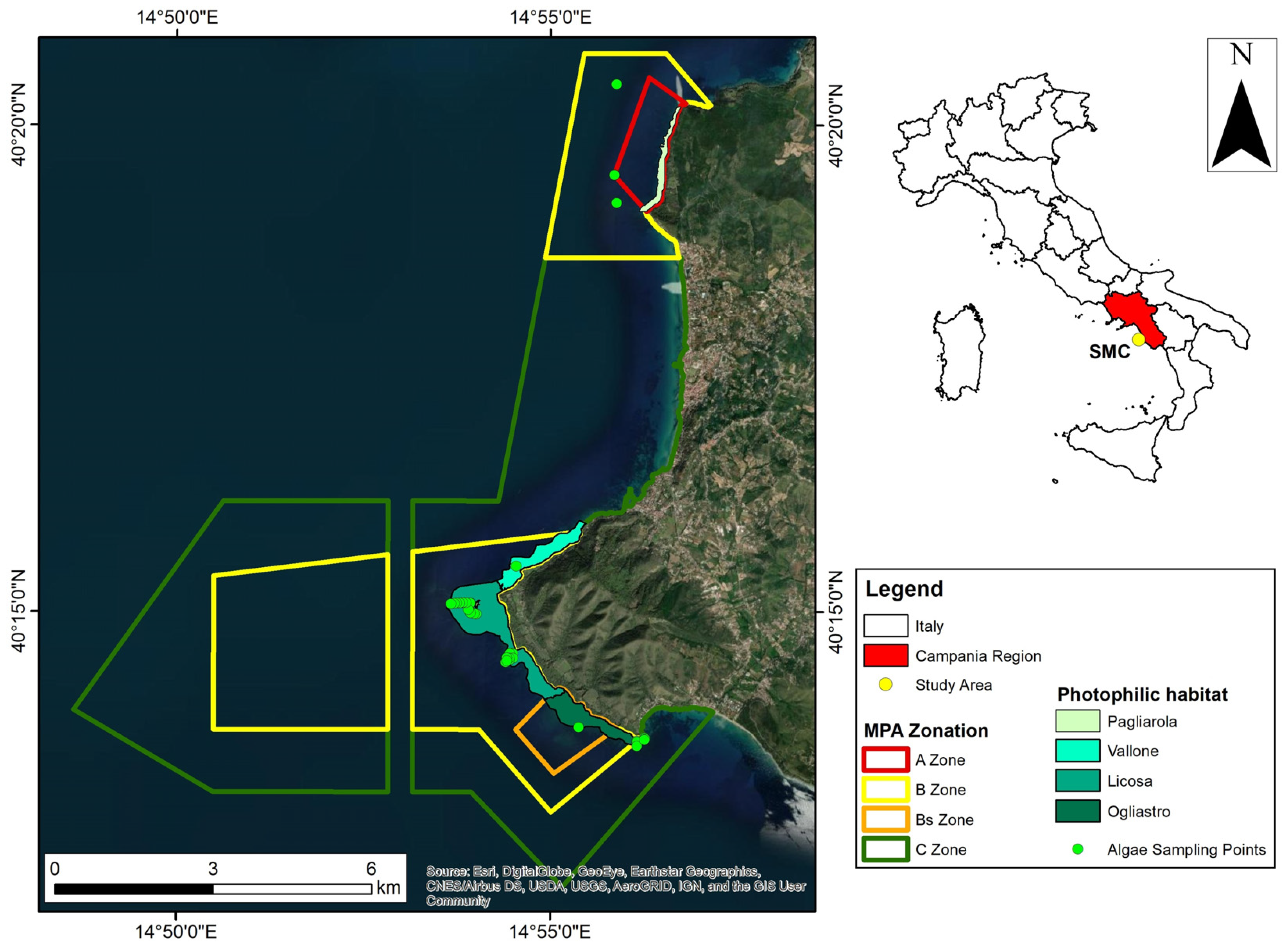
2.1. Study Area
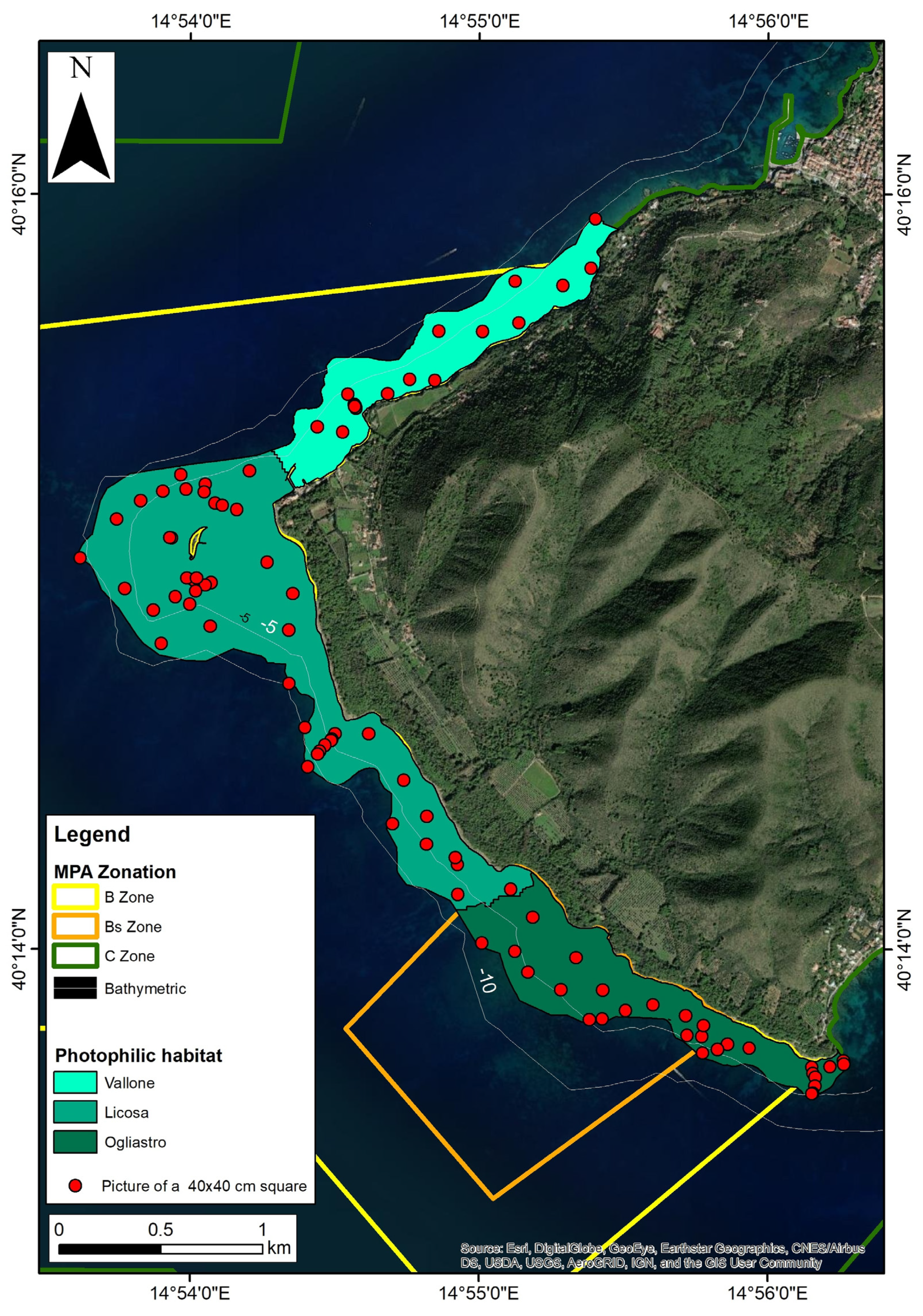
2.2. Field Surveys and Data Collection
2.3. Genetic Identifications
2.4. Geo-Statistical Analyses
3. Results
3.1. Genetic Identifications
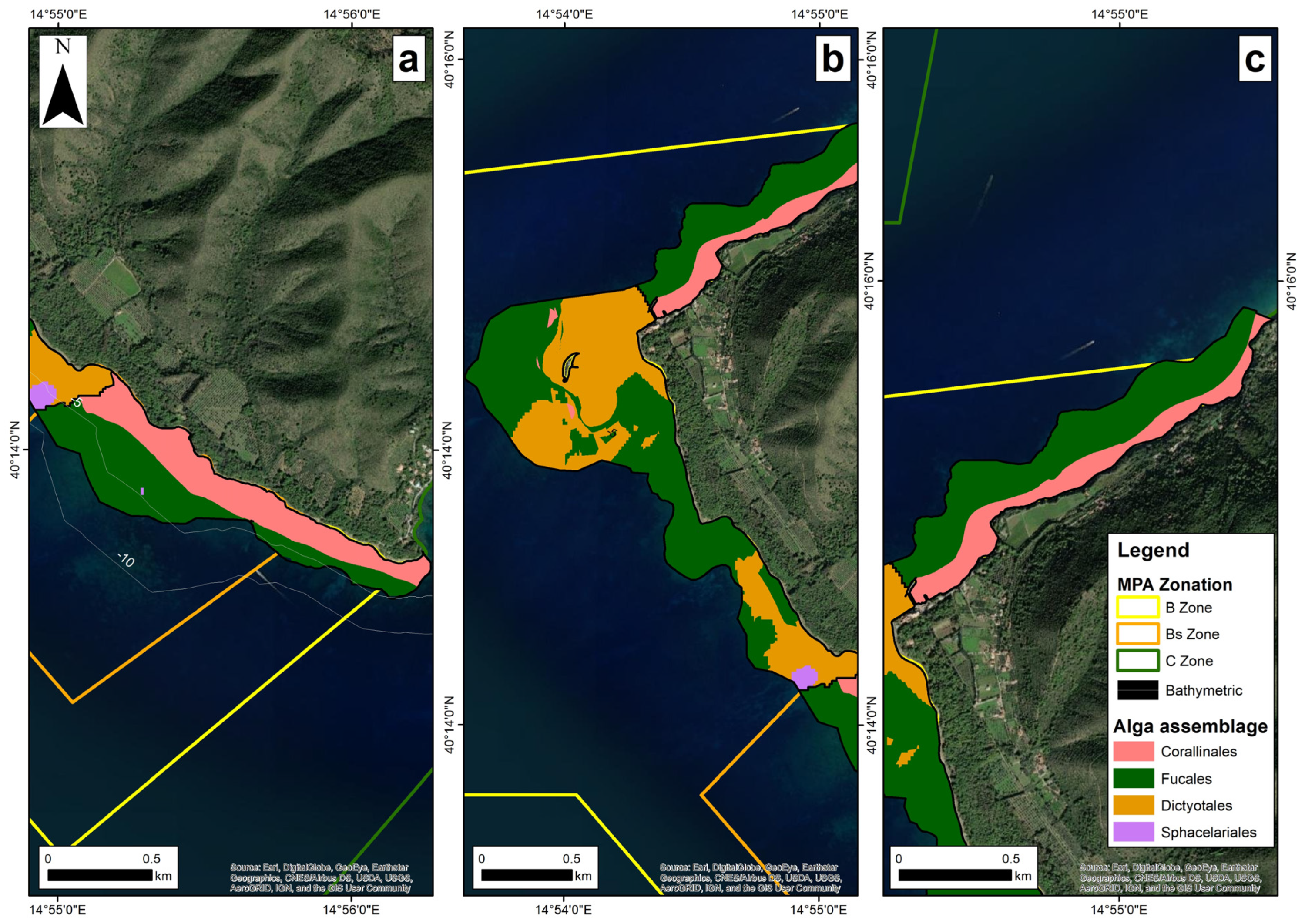
3.2. Spatial Distribution of Photophilic Habitat
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dayton, P.K. Ecology of kelp communities. Annu. Rev. Ecol. Syst. 1985, 16, 215–245. [Google Scholar] [CrossRef]
- Steneck, R.S.; Graham, M.H.; Bourque, B.J.; Corbett, D.; Erlandson, J.M.; Estes, J.A.; Tegner, M.J. Kelp forest ecosystems: Biodiversity, stability, resilience and future. Environ. Conserv. 2002, 29, 436–459. [Google Scholar] [CrossRef] [Green Version]
- Schiel, D.R.; Foster, M.S. The population biology of large brown seaweeds: Ecological consequences of multiphase life histories in dynamic coastal environments. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 343–372. [Google Scholar] [CrossRef] [Green Version]
- Falace, A.; Zanelli, E.; Bressan, G. Morphological and reproductive phenology of Cystoseira compressa (Esper) Gerloff & Nizamuddin (Fucales, Fucophyceae) in the Gulf of Trieste (North Adriatic Sea). In Annales: Series Historia Naturalis; Scientific and Research Center of the Republic of Slovenia: Ljubljana, Slovenia, 2005; Volume 15, p. 71. [Google Scholar]
- Ballesteros, E.; Torras, X.; Pinedo, S.; García, M.; Mangialajo, L.; De Torres, M. A new methodology based on littoral community cartography dominated by macroalgae for the implementation of the European Water Framework Directive. Mar. Pollut. Bull. 2007, 55, 172–180. [Google Scholar] [CrossRef]
- Sales, M.; Ballesteros, E. Shallow Cystoseira (Fucales: Ochrophyta) assemblages thriving in sheltered areas from Menorca (NW Mediterranean): Relationships with environmental factors and anthropogenic pressures. Estuar. Coast. Shelf Sci. 2009, 84, 476–482. [Google Scholar] [CrossRef]
- Lasinio, G.J.; Tullio, M.A.; Ventura, D.; Ardizzone, G.; Abdelahad, N. Statistical analysis of the distribution of infralittoral Cystoseira populations on pristine coasts of four Tyrrhenian islands: Proposed adjustment to the CARLIT index. Ecol. Indic. 2017, 73, 293–301. [Google Scholar] [CrossRef] [Green Version]
- Mancuso, F.P.; Strain, E.M.A.; Piccioni, E.; De Clerck, O.; Sarà, G.; Airoldi, L. Status of vulnerable Cystoseira populations along the Italian infralittoral fringe, and relationships with environmental and anthropogenic variables. Mar. Pollut. Bull. 2018, 129, 762–771. [Google Scholar] [CrossRef]
- Falace, A.; Bressan, G. A qualitative and quantitative analysis of the evolution of macroalgal vegetation on an artificial reef with anti-grazing nets (Loano-Ligurian Sea). ICES J. Mar. Sci. 2002, 59, S150–S156. [Google Scholar] [CrossRef]
- Piazzi, L.; Ceccherelli, G. Concomitance of oligotrophy and low grazing pressure is essential for the resilience of Mediterranean subtidal forests. Mar. Pollut. Bull. 2017, 123, 197–204. [Google Scholar] [CrossRef]
- Savonitto, G.; De La Fuente, G.; Tordoni, E.; Ciriaco, S.; Srijemsi, M.; Bacaro, G.; Chiantore, M.; Falace, A. Addressing reproductive stochasticity and grazing impacts in the restoration of a canopy-forming brown alga by implementing mitigation solutions. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 1611–1623. [Google Scholar] [CrossRef]
- Neiva, J.; Bermejo, R.; Medrano, A.; Capdevila, P.; Milla-Figueras, D.; Afonso, P.; Ballesteros, E.; Sabour, B.; Serio, D.; Nóbrega, E.; et al. DNA barcoding reveals cryptic diversity, taxonomic conflicts and novel biogeographical insights in Cystoseira sl (Phaeophyceae). Eur. J. Phycol. 2022, 1–25. [Google Scholar] [CrossRef]
- Rozic, S.; Puizina, J.; Samanic, I.; Zuljevic, A.; Antolic, B. Molecular identification of the brown algae, Cystoseira spp. (Phaeophycae, Fucales) from theAdriatic Sea–preliminary results. Acta Adriat. 2012, 53, 447–456. [Google Scholar]
- Orellana, S.; Hernández, M.; Sansón, M. Diversity of Cystoseira sensu lato (Fucales, Phaeophyceae) in the eastern Atlantic and Mediterranean based on morphological and DNA evidence, including Carpodesmia gen. emend. and Treptacantha gen. emend. Eur. J. Phycol. 2019, 54, 447–465. [Google Scholar] [CrossRef]
- Buonocore, E.; Grande, U.; Franzese, P.P.; Russo, G.F. Trends and evolution in the concept of marine ecosystem services: An overview. Water 2021, 13, 2060. [Google Scholar] [CrossRef]
- Mancuso, F.P.; Milazzo, M.; Chemello, R. Decreasing in patch-size of Cystoseira forests reduces the diversity of their associated molluscan assemblage in Mediterranean rocky reefs. Estuar. Coast. Shelf Sci. 2021, 250, 107163. [Google Scholar] [CrossRef]
- Benedetti-Cecchi, L.; Cinelli, F. Canopy removal experiments in Cystoseira-dominated rockpools from the Western coast of the Mediterranean (Ligurian Sea). J. Exp. Mar. Biol. Ecol. 1992, 155, 69–83. [Google Scholar] [CrossRef]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rais Lasram, F.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The biodiversity of the Mediterranean Sea: Estimates, patterns, and threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [Green Version]
- Sales, M.; Cebrian, E.; Tomas, F.; Ballesteros, E. Pollution impacts and recovery potential in three species of the genus Cystoseira (Fucales, Heterokontophyta). Estuar. Coast. Shelf Sci. 2011, 92, 347–357. [Google Scholar] [CrossRef]
- Rendina, F.; Ferrigno, F.; Appolloni, L.; Donnarumma, L.; Sandulli, R.; Russo, G.F. Anthropic pressure due to lost fishing gears and marine litter on different rhodolith beds off the Campania Coast (Tyrrhenian Sea, Italy). Ecol. Quest. 2020, 31, 41–51. [Google Scholar] [CrossRef]
- Ferrigno, F.; Appolloni, L.; Donnarumma, L.; Di Stefano, F.; Rendina, F.; Sandulli, R.; Russo, G.F. Diversity loss in coralligenous structuring species impacted by fishing gear and marine litter. Diversity 2021, 13, 331. [Google Scholar] [CrossRef]
- Fabbrizzi, E.; Scardi, M.; Ballesteros, E.; Benedetti-Cecchi, L.; Cebrian, E.; Ceccherelli, G.; De Leo, D.; Deidun, A.; Guarnieri, G.; Falace, A.; et al. Modeling macroalgal forest distribution at Mediterranean scale: Present status, drivers of changes and insights for conservation and management. Front. Mar. Sci. 2020, 7, 20. [Google Scholar] [CrossRef]
- Wernberg, T.; Russell, B.D.; Moore, P.J.; Ling, S.D.; Smale, D.A.; Campbell, A.; Coleman, M.A.; Steinberg, P.D.; Kendrick, G.A.; Connell, S.D. Impacts of climate change in a global hotspot for temperate marine biodiversity and ocean warming. J. Exp. Mar. Biol. Ecol. 2011, 400, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Sunday, J.M.; Fabricius, K.E.; Kroeker, K.J.; Anderson, K.M.; Brown, N.E.; Barry, J.P.; Connell, S.D.; Dupont, S.; Gaylord, B.; Hall-Spencer, J.M.; et al. Ocean acidification can mediate biodiversity shifts by changing biogenic habitat. Nat. Clim. Chang. 2017, 7, 81–85. [Google Scholar] [CrossRef]
- Buonomo, R.; Chefaoui, R.M.; Lacida, R.B.; Engelen, A.H.; Serrão, E.A.; Airoldi, L. Predicted extinction of unique genetic diversity in marine forests of Cystoseira spp. Mar. Environ. Res. 2018, 138, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Rendina, F.; Bouchet, P.J.; Appolloni, L.; Russo, G.F.; Sandulli, R.; Kolzenburg, R.; Putra, A.; Ragazzola, F. Physiological response of the coralline alga Corallina officinalis L. to both predicted long-term increases in temperature and short-term heatwave events. Mar. Environ. Res. 2019, 150, 104764. [Google Scholar] [CrossRef] [PubMed]
- Bevilacqua, S.; Savonitto, G.; Lipizer, M.; Mancuso, P.; Ciriaco, S.; Srijemsi, M.; Falace, A. Climatic anomalies may create a long-lasting ecological phase shift by altering the reproduction of a foundation species. Ecology 2019, 100, e02838. [Google Scholar] [CrossRef]
- Falace, A.; Marletta, G.; Savonitto, G.; Carniel, F.C.; Srijemsi, M.; Bevilacqua, S.; Tretiach, M.; Alongi, G. Is the South-Mediterranean Canopy-Forming Ericaria giacconei (=Cystoseira hyblaea) a Loser From Ocean Warming? Front. Mar. Sci. 2021, 8, 1758. [Google Scholar] [CrossRef]
- Benedetti-Cecchi, L.; Pannacciulli, F.; Bulleri, F.; Moschella, P.S.; Airoldi, L.; Relini, G.; Cinelli, F. Predicting the consequences of anthropogenic disturbance: Large-scale effects of loss of canopy algae on rocky shores. Mar. Ecol. Prog. Ser. 2001, 214, 137–150. [Google Scholar] [CrossRef] [Green Version]
- Soltan, D.; Verlaque, M.; Boudouresque, C.F.; Francour, P. Changes in macroalgal communities in the vicinity of a Mediterranean sewage outfall after the setting up of a treatment plant. Mar. Pollut. Bull. 2001, 42, 59–70. [Google Scholar] [CrossRef]
- Thibaut, T.; Pinedo, S.; Torras, X.; Ballesteros, E. Long-term decline of the populations of Fucales (Cystoseira spp. and Sargassum spp.) in the Alberes coast (France, North-western Mediterranean). Mar. Pollut. Bull. 2005, 50, 1472–1489. [Google Scholar] [CrossRef]
- Mangialajo, L.; Chiantore, M.; Cattaneo-Vietti, R. Changes in marine macroalgal assemblages due to human impacts: General trends and applications in coastal management. Biol. Mar. Mediterr. 2008, 15, 115–117. [Google Scholar]
- Falace, A.; Alongi, G.; Cormaci, M.; Furnari, G.; Curiel, D.; Cecere, E.; Petrocelli, A. Changes in the benthic algae along the Adriatic Sea in the last three decades. Chem. Ecol. 2010, 26, 77–90. [Google Scholar] [CrossRef]
- Perkol-Finkel, S.; Airoldi, L. Loss and recovery potential of marine habitats: An experimental study of factors maintaining resilience in subtidal algal forests at the Adriatic Sea. PLoS ONE 2010, 5, e10791. [Google Scholar] [CrossRef] [PubMed]
- Connell, S.D.; Foster, M.S.; Airoldi, L. What are algal turfs? Towards a better description of turfs. Mar. Ecol. Prog. Ser. 2014, 495, 299–307. [Google Scholar] [CrossRef] [Green Version]
- Strain, E.M.A.; Thomson, R.J.; Micheli, F.; Mancuso, F.P.; Airoldi, L. Identifying the interacting roles of stressors in driving the global loss of canopy-forming to mat-forming algae in marine ecosystems. Glob. Change Biol. 2014, 20, 3300–3312. [Google Scholar] [CrossRef]
- Agnetta, D.; Badalamenti, F.; Ceccherelli, G.; Di Trapani, F.; Bonaviri, C.; Gianguzza, P. Role of two co-occurring Mediterranean Sea urchins in the formation of barren from Cystoseira canopy. Estuar. Coast. Shelf Sci. 2015, 152, 73–77. [Google Scholar] [CrossRef] [Green Version]
- Catra, M.; Alongi, G.; Leonardi, R.; Negri, M.P.; Sanfilippo, R.; Sciuto, F.; Serio, D.; Viola, A.; ROSSO, A. Degradation of a photophilic algal community and its associated fauna from eastern Sicily (Mediterranean Sea). Mediterr. Mar. Sci. 2019, 20, 74–89. [Google Scholar] [CrossRef] [Green Version]
- Airoldi, L.; Ballesteros, E.; Buonuomo, R.; Van Belzen, J.; Bouma, T.J.; Cebrian, E.; de Clerk, O.; Engelen, A.H.; Ferrario, F.; Fraschetti, S.; et al. Marine forests at risk: Solutions to halt the loss and promote the recovery of Mediterranean canopy-forming seaweeds. In Proceedings of the 5th Mediterranean Symposium on Marine Vegetation, Portoroz, Slovenia, 27–28 October 2014; pp. 28–33. [Google Scholar]
- Blanfuné, A.; Boudouresque, C.F.; Verlaque, M.; Thibaut, T. The fate of Cystoseira crinita, a forest-forming Fucale (Phaeophyceae, Stramenopiles), in France (North Western Mediterranean Sea). Estuar. Coast. Shelf Sci. 2016, 181, 196–208. [Google Scholar] [CrossRef]
- Porzio, L.; Grech, D.; Buia, M.C. Long-term changes (1800–2019) in marine vegetational habitats: Insights from a historic industrialised coastal area. Mar. Environ. Res. 2020, 161, 105003. [Google Scholar]
- Thibaut, T.; Blanfuné, A.; Markovic, L.; Verlaque, M.; Boudouresque, C.F.; Perret-Boudouresque, M.; Maćic, V.; Bottin, L. Unexpected abundance and long-term relative stability of the brown alga Cystoseira amentacea, hitherto regarded as a threatened species, in the north-western Mediterranean Sea. Mar. Pollut. Bull. 2014, 89, 305–323. [Google Scholar] [CrossRef]
- Thibaut, T.; Blanfuné, A.; Boudouresque, C.F.; Cottalorda, J.M.; Hereu, B.; Susini, M.L.; Verlaque, M. Unexpected temporal stability of Cystoseira and Sargassum forests in Port-Cros, one of the oldest Mediterranean marine National Parks. Cryptogam. Algol. 2016, 37, 61–90. [Google Scholar] [CrossRef]
- Falace, A.; Kaleb, S.; De La Fuente, G.; Asnaghi, V.; Chiantore, M. Ex situ cultivation protocol for Cystoseira amentacea var. stricta (Fucales, Phaeophyceae) from a restoration perspective. PLoS ONE 2018, 13, e0193011. [Google Scholar]
- Verdura, J.; Sales, M.; Ballesteros, E.; Cefalì, M.E.; Cebrian, E. Restoration of a canopy-forming alga based on recruitment enhancement: Methods and long-term success assessment. Front. Plant Sci. 2018, 9, 1832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De La Fuente, G.; Chiantore, M.; Asnaghi, V.; Kaleb, S.; Falace, A. First ex situ outplanting of the habitat-forming seaweed Cystoseira amentacea var. stricta from a restoration perspective. PeerJ 2019, 7, e7290. [Google Scholar] [CrossRef] [Green Version]
- Savonitto, G.; Alongi, G.; Falace, A. Reproductive phenology, zygote embryology and germling development of the threatened Carpodesmia barbatula (=Cystoseira barbatula) (Fucales, Phaeophyta) towards its possible restoration. Webbia 2019, 74, 317–323. [Google Scholar] [CrossRef]
- Orlando-Bonaca, M.; Pitacco, V.; Slavinec, P.; Šiško, M.; Makovec, T.; Falace, A. First Restoration Experiment for Gongolaria barbata in Slovenian Coastal Waters. What Can Go Wrong? Plants 2021, 10, 239. [Google Scholar] [CrossRef] [PubMed]
- Orlando-Bonaca, M.; Savonitto, G.; Asnaghi, V.; Trkov, D.; Pitacco, V.; Šiško, M.; Makovec, T.; Slavinec, P.; Lokovšek, A.; Ciriaco, S.; et al. Where and how-new insight for brown algal forest restoration in the Adriatic. Front. Mar. Sci. 2022, 9, 988584. [Google Scholar] [CrossRef]
- Orfanidis, S.; Panayotidis, P.; Stamatis, N. An insight to the ecological evaluation index (EEI). Ecol. Indic. 2003, 3, 27–33. [Google Scholar] [CrossRef]
- Blowes, S.A.; Chase, J.M.; Di Franco, A.; Frid, O.; Gotelli, N.J.; Guidetti, P.; Knight, T.M.; May, F.; McGlinn, D.; Micheli, F.; et al. Mediterranean marine protected areas have higher biodiversity via increased evenness, not abundance. J. Appl. Ecol. 2020, 57, 578–589. [Google Scholar] [CrossRef] [Green Version]
- Crosti, R.; Arcangeli, A.; Campagnuolo, S.; Castriota, L.; Falautano, M.; Maggio, T.; Andaloro, F. Assessing worth of marine protected areas for the protection of threatened biodiversity using IUCN Red List and Red List Index. A pilot study in six mediterranean areas. Ecol. Indic. 2020, 119, 106765. [Google Scholar] [CrossRef]
- Ferrigno, F.; Appolloni, L.; Rendina, F.; Donnarumma, L.; Russo, G.F.; Sandulli, R. Red coral (Corallium rubrum) populations and coralligenous characterization within “Regno di Nettuno MPA” (Tyrrhenian Sea, Italy). Eur. Zool. J. 2020, 87, 203–213. [Google Scholar] [CrossRef] [Green Version]
- Meehan, M.C.; Ban, N.C.; Devillers, R.; Singh, G.G.; Claudet, J. How far have we come? A review of MPA network performance indicators in reaching qualitative elements of Aichi Target 11. Conserv. Lett. 2020, 13, e12746. [Google Scholar] [CrossRef]
- Edwards, P.; Bird, E.; Cotgreave, B.; Cossins, A.; Crompton, K.; Fowler, W.; De Leo, F.; Deidun, A.; Guarnieri, G.; Falace, A.; et al. Marine phytobenthos of the Castellabate (Cilento) Natural Park, Salerno, Italy. Phytocoenologia 1975, 403–426. [Google Scholar] [CrossRef]
- Appolloni, L.; Russo, G.F. In the Kingdom of Leucosia. A Guide to the Virtual Aquarium of the Santa Maria di Castellabate Marine Protected Area; Edizioni dell’Ippogrifo: Sarno, Italy, 2021; pp. 14–29. [Google Scholar]
- Russo, G.F.; Di Donato, R.; Di Stefano, F. Gli habitat sottomarini delle coste della Campania. Biol. Ital. 2008, 6, 36–40. [Google Scholar]
- Pérès, J.M.; Picard, J. Nouveau Manuel de Bionomie Benthique de la mer Méditerranée; Station Marine d’Endoume: Marseille, France, 1964. [Google Scholar]
- Lane, C.E.; Lindstrom, S.C.; Saunders, G.W. A molecular assessment of northeast Pacific Alaria species (Laminariales, Phaeophyceae) with reference to the utility of DNA barcoding. Mol. Phylogenetics Evol. 2007, 44, 634–648. [Google Scholar] [CrossRef]
- Silberfeld, T.; Leigh, J.W.; Verbruggen, H.; Cruaud, C.; De Reviers, B.; Rousseau, F. A multi-locus time-calibrated phylogeny of the brown algae (Heterokonta, Ochrophyta, Phaeophyceae): Investigating the evolutionary nature of the “brown algal crown radiation”. Mol. Phylogenetics Evol. 2010, 56, 659–674. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication; National University of Ireland: Galway, Ireland, 2023; Available online: http://www.algaebase.org (accessed on 14 February 2023).
- Krivoruchko, K. Empirical Bayesian Kriging Implemented in ArcGIS Geostatistical Analyst. ArcUser Fall 2012, 6, 1150. [Google Scholar]
- Boudouresque, C.F. Marine biodiversity in the Mediterranean: Status of species, populations and communities. Sci. Rep. Port-Cros Natl. Park 2004, 20, 97–146. [Google Scholar]
- Thibaut, T.; Blanfuné, A.; Boudouresque, C.F.; Verlaque, M. Decline and local extinction of Fucales in French Riviera: The harbinger of future extinctions? Mediterr. Mar. Sci. 2015, 16, 206–224. [Google Scholar] [CrossRef] [Green Version]
- Micheli, F.; Levin, N.; Giakoumi, S.; Katsanevakis, S.; Abdulla, A.; Coll, M.; Fraschetti, S.; Kark, S.; Koutsoubas, D.; Mackelworth, P.; et al. Setting priorities for regional conservation planning in the Mediterranean Sea. PLoS ONE 2013, 8, e59038. [Google Scholar] [CrossRef] [Green Version]
- Verlaque, M.; Ballesteros, E.; Sala, E.; Garrabou, J. Cystoseira jabukae (Cystoseiraceae, Fucophyceae) from Corsica (Mediterranean) with notes on the previously misunderstood species C. funkii. Phycologia 1999, 38, 77–86. [Google Scholar] [CrossRef]
- Serio, D.; Furnari, G. Ericaria giacconei sp. nov.(Sargassaceae, Fucophyceae), the species to which the invalidly published Cystoseira hyblaea Giaccone should be referred. Cryptogam. Algol. 2021, 42, 141–151. [Google Scholar] [CrossRef]
- Sadogurska, S.S.; Neiva, J.; Serrão, E.A.; Falace, A.; Israel, A. The genus Cystoseira s.l. (Ochrophyta, Fucales, Sargassaceae) in the Black Sea: Morphological variability and molecular taxonomy of Gongolaria barbata and endemic Ericaria crinita f. bosphorica comb. nov. Phytotaxa 2021, 480, 1–21. [Google Scholar] [CrossRef]
- Hereu, B.; Mangialajo, L.; Ballesteros, E.; Thibaut, T. On the occurrence, structure and distribution of deep-water Cystoseira populations in the Port-Cros National Park (North-western Mediterranean). Eur. J. Phycol. 2008, 43, 263–273. [Google Scholar] [CrossRef]
- Ballesteros, E.; Garrabou, J.; Hereu, B.; Zabala, M.; Cebrian, E.; Sala, E. Deep-water stands of Cystoseira zosteroides C. Agardh (Fucales, Ochrophyta) in the Northwestern Mediterranean: Insights into assemblage structure and population dynamics. Estuar. Coast. Shelf Sci. 2009, 82, 477–484. [Google Scholar] [CrossRef]
- Ballesteros, E.; Sala, E.; Garrabou, J.; Zabala, M. Community structure and frond size distribution of a deep water stand of Cystoseira spinosa (Phaeophyta) in the northwestern Mediterranean. Eur. J. Phycol. 1998, 33, 121–128. [Google Scholar] [CrossRef]
- Giaccone, G. Écologie et chorologie des Cystoseira de Méditerranée. Rapp. Commun. Int. Mer. Méditerranéenne 1973, 22, 49–50. [Google Scholar]
- Capdevila, P.; Linares, C.; Aspillaga, E.; Riera, J.L.; Hereu, B. Effective dispersal and density-dependence in mesophotic macroalgal forests: Insights from the Mediterranean species Cystoseira zosteroides. PLoS ONE 2018, 13, e0191346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capdevila, P.; Linares, C.; Aspillaga, E.; Navarro, L.; Kersting, D.K.; Hereu, B. Recruitment patterns in the Mediterranean deep-water alga Cystoseira Zosteroides. Mar. Biol. 2015, 162, 1165–1174. [Google Scholar] [CrossRef]
- Serio, D.; Alongi, G.; Catra, M.; Cormaci, M.; Furnari, G. Changes in the benthic algal flora of Linosa Island (Straits of Sicily, Mediterranean Sea). Bot. Mar. 2006, 49, 135–144. [Google Scholar] [CrossRef]
- Boudouresque, C.F.; Meinesz, A.; Ballesteros, E.; Ben Maiz, N.; Boisset, F.; Cinelli, F.; Cirik, S.; Cormaci, M.; Jeudy de Grissac, A.; Laborel, J.; et al. Livre Rouge ‘‘Gèrard Vuignier’’ des Végétaux, Peuplements et Paysages Marins Menacés de Méditerranée; MAP Technical Report Series 43; UNEP/IUCN/GIS: Posidonie, Athens, 1990; p. 245. [Google Scholar]
- Cormaci, M.; Furnari, G.; Alongi, G.; Catra, M.; Pizzuto, F.; Serio, D. Spring marine vegetation on rocky substrata of the Tremiti islands (Adriatic Sea, Italy). In Structures and Processes in the Mediterranean Ecosystems; Faranda, F., Spezie, G., Guglielmo, L., Eds.; Springer: Milano, Italy, 2001; pp. 245–254. [Google Scholar]
- Alongi, G.; Catra, M.; Cormaci, M.; Furnari, G.; Serio, D. Spring marine vegetation on rocky substrata of Pantelleria Island (the Straits of Sicily, Italy). Nova Hedwig. 2004, 79, 447–478. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Genetic ID (MOTU) | Comment |
|---|---|---|
| CIL7A | Cystoseira pustulata | Confirmed species throughout the whole Mediterranean, Macaronesia |
| CIL8A | ||
| CIL8B | ||
| CIL1A | Ericaria brachycarpa | Confirmed at least from the central Mediterranean, apparently replaced in the western Mediterranean by Ericaria balearica |
| CIL3A | Ericaria crinita/ E. barbatula | Species complex confirmed throughout the whole Mediterranean |
| CIL12A | Ericaria funkii | So far sequenced only from Cilento. Reported also from the western Med. but additional sequences required to confirm it is the same taxon |
| CIL12B | ||
| CIL4A | Gongolaria sp. 2 | Gongolaria sp. 2 has been commonly identified as G. elegans, also as G. montagnei or G. squarrosa, but represents a distinct entity confirmed from throughout the central and eastern Mediterranean. The true G. montagnei and G. elegans are apparently restricted to the western Mediterranean, where they appear to replace Gongolaria sp. 2 (the names apply to these western species on the basis of type material) |
| CIL4B | ||
| CIL5A | ||
| CIL9A | ||
| CIL11A |
| Species | Site | Depth (m) |
|---|---|---|
| Cystoseira pustulata (Ercegovic) Neiva and Serrão | O, L | 3–12 |
| Cystoseira compressa (Esper) Gerloff and Nizamuddin | L | 0.2–0.5 |
| Ericaria corniculata (Turner) Neiva and Serrão | O, L, V | 2–8 |
| E. crinita/barbatula (as Cystoseira crinitophylla) | L | 3–12 |
| Gongolaria sp. 2 (as Gongolaria elegans) | O, L | 4–12 |
| Ericaria brachycarpa (J. Agardh) Molinari and Guiry | O, L, V | 2–10 |
| Cystoseira foeniculacea (Linnaeus) Greville | L | 5–12 |
| Gongolaria montagnei (J. Agardh) Kuntze | P | 35 |
| Ericaria funkii (Gerloff and Nizamuddin) Molinari and Guiry | P | 30 |
| Ericaria zosteroides (C. Agardh) Molinari and Guiry | P | 35 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rendina, F.; Falace, A.; Alongi, G.; Buia, M.C.; Neiva, J.; Appolloni, L.; Marletta, G.; Russo, G.F. The Lush Fucales Underwater Forests off the Cilento Coast: An Overlooked Mediterranean Biodiversity Hotspot. Plants 2023, 12, 1497. https://doi.org/10.3390/plants12071497
Rendina F, Falace A, Alongi G, Buia MC, Neiva J, Appolloni L, Marletta G, Russo GF. The Lush Fucales Underwater Forests off the Cilento Coast: An Overlooked Mediterranean Biodiversity Hotspot. Plants. 2023; 12(7):1497. https://doi.org/10.3390/plants12071497
Chicago/Turabian StyleRendina, Francesco, Annalisa Falace, Giuseppina Alongi, Maria Cristina Buia, João Neiva, Luca Appolloni, Giuliana Marletta, and Giovanni Fulvio Russo. 2023. "The Lush Fucales Underwater Forests off the Cilento Coast: An Overlooked Mediterranean Biodiversity Hotspot" Plants 12, no. 7: 1497. https://doi.org/10.3390/plants12071497
APA StyleRendina, F., Falace, A., Alongi, G., Buia, M. C., Neiva, J., Appolloni, L., Marletta, G., & Russo, G. F. (2023). The Lush Fucales Underwater Forests off the Cilento Coast: An Overlooked Mediterranean Biodiversity Hotspot. Plants, 12(7), 1497. https://doi.org/10.3390/plants12071497