

Influence of a phyA Mutation on Polyamine Metabolism in Arabidopsis Depends on Light Spectral Conditions
 , ,
, ,  , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Measurements of the Shoot Weight and Chlorophyll-A Fluorescence Induction Parameters
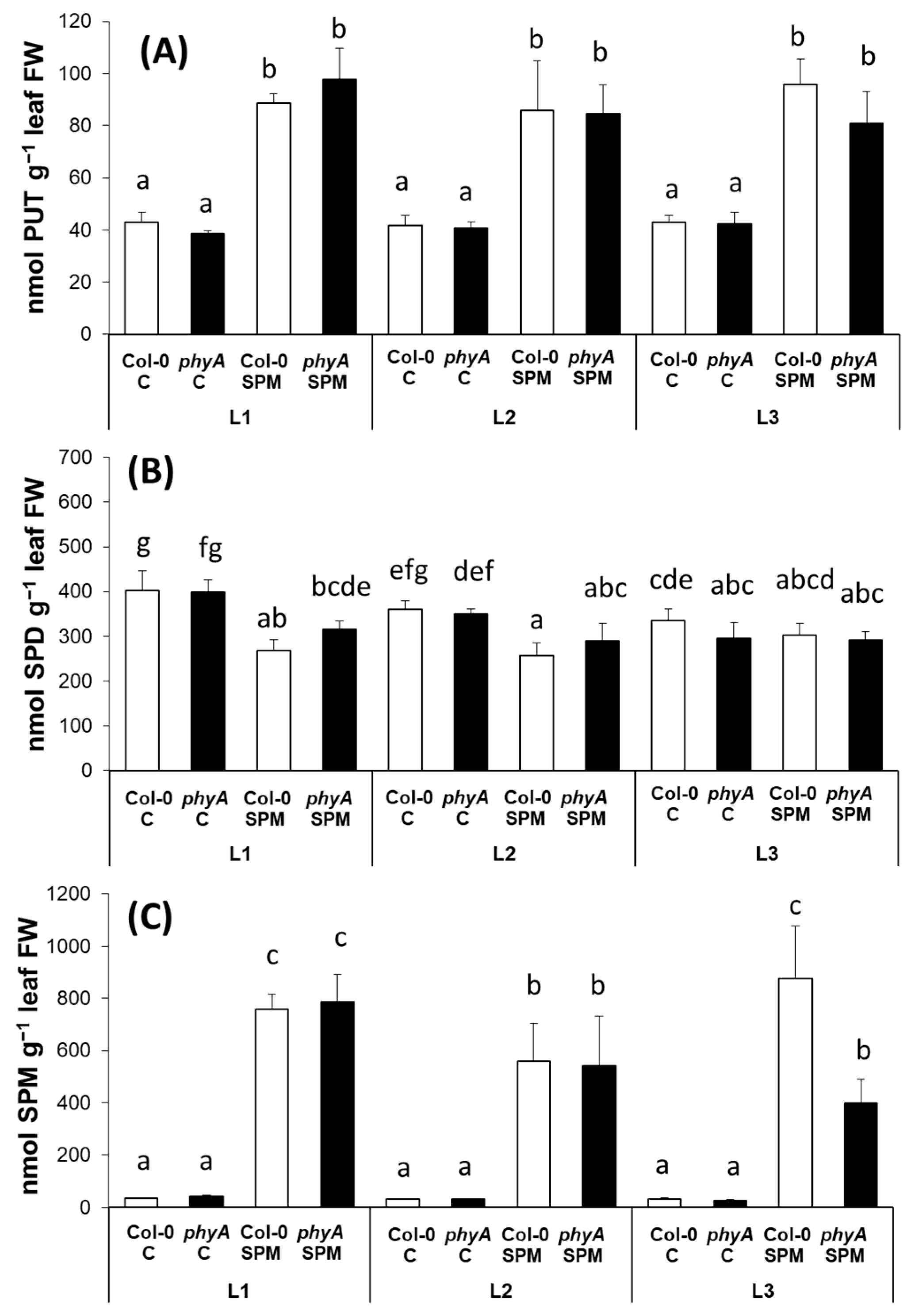
2.2. Modulation of Polyamine Content and Polyamine Metabolism-Related Gene Expression by Light Spectra
2.3. Correlation Analyses and Principal Components Analysis (PCA) of the Measured Parameters
3. Discussion
4. Materials and Methods
4.1. Plant Material, Plant Growth Conditions and Treatments
4.2. Chlorophyll-A Fluorescence Induction (FI) Analysis
4.3. Polyamine Analysis
4.4. Gene Expression Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Poorter, H.; Niinemets, Ü.; Ntagkas, N.; Siebenkäs, A.; Mäenpää, M.; Matsubara, S.; Pons, T. A meta-analysis of plant responses to light intensity for 70 traits ranging from molecules to whole plant performance. New Phytol. 2019, 223, 1073–1105. [Google Scholar] [CrossRef] [PubMed]
- Sharrock, R.A. The phytochrome red/far-red photoreceptor superfamily. Genome Biol. 2008, 9, 230. [Google Scholar] [CrossRef] [PubMed]
- Legris, M.; Ince, Y.Ç.; Fankhauser, C. Molecular mechanisms underlying phytochrome-controlled morphogenesis in plants. Nat. Commun. 2019, 10, 5219. [Google Scholar] [CrossRef]
- Sharrock, R.A.; Clack, T. Patterns of expression and normalized levels of the five Arabidopsis phytochromes. Plant Physiol. 2002, 130, 442–456. [Google Scholar] [CrossRef] [PubMed]
- Fankhauser, C. The phytochromes, a family of red/far-red absorbing photoreceptors. J. Biol. Chem. 2001, 276, 11453–11456. [Google Scholar] [CrossRef]
- Oh, S.; Warnasooriya, S.N.; Montgomery, B.L. Downstream effectors of light- and phytochrome-dependent regulation of hypocotyl elongation in Arabidopsis thaliana. Plant Mol. Biol. 2013, 81, 627–640. [Google Scholar] [CrossRef]
- Wimalasekera, R.; Villar, C.; Begum, T.; Scherer, G.F. COPPER AMINE OXIDASE1 (CuAO1) of Arabidopsis thaliana contributes to abscisic acid- and polyamine-induced nitric oxide biosynthesis and abscisic acid signal transduction. Mol. Plant 2011, 4, 663–678. [Google Scholar] [CrossRef]
- Arana, M.V.; Tognacca, R.S.; Estravis-Barcalá, M.; Sánchez, R.A.; Botto, J.F. Physiological and molecular mechanisms underlying the integration of light and temperature cues in Arabidopsis thaliana seeds. Plant Cell Environ. 2017, 40, 3113–3121. [Google Scholar] [CrossRef]
- Xiang, S.; Wu, S.; Jing, Y.; Chen, L.; Yu, D. Phytochrome B regulates jasmonic acid-mediated defense response against Botrytis cinerea in Arabidopsis. Plant Divers. 2022, 44, 109–115. [Google Scholar] [CrossRef]
- Desai, M.; Hu, J. Light induces peroxisome proliferation in Arabidopsis seedlings through the photoreceptor phytochrome A, the transcription factor HY5 HOMOLOG, and the peroxisomal protein PEROXIN11b. Plant Physiol. 2008, 146, 1117–1127. [Google Scholar] [CrossRef]
- Prerostova, S.; Dobrev, P.I.; Knirsch, V.; Jarosova, J.; Gaudinova, A.; Zupkova, B.; Prášil, I.T.; Janda, T.; Brzobohatý, B.; Skalák, J.; et al. Light quality and intensity modulate cold acclimation in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 2736. [Google Scholar] [CrossRef] [PubMed]
- Jumtee, K.; Bamba, T.; Okazawa, A.; Fukusaki, E.; Kobayashi, A. Integrated metabolite and gene expression profiling revealing phytochrome A regulation of polyamine biosynthesis of Arabidopsis thaliana. J. Exp. Bot. 2008, 59, 1187–1200. [Google Scholar] [CrossRef] [PubMed]
- Jumtee, K.; Okazawa, A.; Harada, K.; Fukusaki, E.; Takano, M.; Kobayashi, A. Comprehensive metabolite profiling of phyA phyB phyC triple mutants to reveal their associated metabolic phenotype in rice leaves. J. Biosci. Bioeng. 2009, 108, 151–159. [Google Scholar] [CrossRef]
- Han, X.; Tohge, T.; Lalor, P.; Dockery, P.; Devaney, N.; Esteves-Ferreira, A.A.; Fernie, A.R.; Sulpice, R. Phytochrome A and B regulate primary metabolism in Arabidopsis leaves in response to light. Front. Plant Sci. 2017, 8, 1394. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Źeljković, S.C.; Piskurewicz, U.; Megies, C.; Tarkowski, P.; Lopez-Molina, L. Polyamine uptake transporter 2 (put2) and decaying seeds enhance phyA-mediated germination by overcoming PIF1 repression of germination. PLoS Genet. 2019, 15, e1008292. [Google Scholar] [CrossRef]
- Chun, L.; Kawakami, A.; Christopher, D.A. Phytochrome A mediates blue light and UV-A-dependent chloroplast gene transcription in green leaves. Plant Physiol. 2001, 125, 1957–1966. [Google Scholar] [CrossRef]
- Sullivan, S.; Hart, J.E.; Rasch, P.; Walker, C.H.; Christie, J.M. Phytochrome A mediates blue-light enhancement of second-positive phototropism in Arabidopsis. Front. Plant Sci. 2016, 7, 290. [Google Scholar] [CrossRef]
- Warnasooriya, S.N.; Porter, K.J.; Montgomery, B.L. Tissue- and isoform-specific phytochrome regulation of light-dependent anthocyanin accumulation in Arabidopsis thaliana. Plant Signal. Behav. 2011, 6, 624–631. [Google Scholar] [CrossRef]
- Kong, Y.; Zheng, Y. Phototropin is partly involved in blue-light-mediated stem elongation, flower initiation, and leaf expansion: A comparison of phenotypic responses between wild Arabidopsis and its phototropin mutants. Environ. Exp. Bot. 2020, 171, 103967. [Google Scholar] [CrossRef]
- Usami, T.; Mochizuki, N.; Kondo, M.; Nishimura, M.; Nagatani, A. Cryptochromes and phytochromes synergistically regulate Arabidopsis root greening under blue light. Plant Cell Physiol. 2004, 45, 1798–1808. [Google Scholar] [CrossRef]
- Sheng, S.; Wu, C.; Xiang, Y.C.; Pu, W.X.; Duan, S.H.; Huang, P.J.; Cheng, X.Y.; Gong, Y.Y.; Liang, Y.L.; Liu, L.H. Polyamine: A potent ameliorator for plant growth response and adaption to abiotic stresses particularly the ammonium stress antagonized by urea. Front. Plant Sci. 2022, 13, 783597. [Google Scholar] [CrossRef] [PubMed]
- Pál, M.; Szalai, G.; Janda, T. Speculation: Polyamines are important in abiotic stress signaling. Plant Sci. 2015, 237, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Li, B.; Li, G.; Charron, J.-B.; Dai, M.; Shi, X.; Deng, X.W. Arabidopsis Phytochrome A directly targets numerous promoters for individualized modulation of genes in a wide range of pathways. Plant Cell 2014, 26, 1949–1966. [Google Scholar] [CrossRef]
- Pál, M.; Szalai, G.; Gondor, O.K.; Janda, T. Unfinished story of polyamines: Role of conjugation, transport and light-related regulation in the polyamine metabolism in plants. Plant Sci. 2021, 308, 110923. [Google Scholar] [CrossRef] [PubMed]
- Hummel, I.; Bourdais, G.; Gouesbet, G.; Couée, I.; Malmber, R.L.; El Amrani, A. Differential gene expression of arginine decarboxylase ADC1 and ADC2 in Arabidopsis thaliana: Characterization of transcriptional regulation during seed germination and seedling development. New Phytol. 2004, 163, 519–531. [Google Scholar] [CrossRef]
- Marco, F.; Busó, E.; Carrasco, P. Overexpression of SAMDC1 gene in Arabidopsis thaliana increases expression of defense-related genes as well as resistance to Pseudomonas syringae and Hyaloperonospora arabidopsidis. Front. Plant Sci. 2014, 5, 115. [Google Scholar] [CrossRef]
- Urano, K.; Yoshiba, Y.; Nanjo, T.; Igarashi, Y.; Seki, M.; Sekiguchi, F.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Characterization of Arabidopsis genes involved in biosynthesis of polyamines in abiotic stress responses and developmental stages. Plant Cell Environ. 2003, 26, 1917–1926. [Google Scholar] [CrossRef]
- Hanzawa, Y.; Imai, A.; Michael, A.J.; Komeda, Y.; Takahashi, T. Characterization of the spermidine synthase-related gene family in Arabidopsis thaliana. FEBS Lett. 2002, 527, 176–180. [Google Scholar] [CrossRef]
- Planas-Portell, J.; Gallart, M.; Tiburcio, A.F.; Altabella, T. Copper-containing amine oxidases contribute to terminal polyamine oxidation in peroxisomes and apoplast of Arabidopsis thaliana. BMC Plant Biol. 2013, 13, 109. [Google Scholar] [CrossRef]
- Takahashi, Y.; Cong, R.; Sagor, G.H.; Niitsu, M.; Berberich, T.; Kusano, T. Characterization of five polyamine oxidase isoforms in Arabidopsis thaliana. Plant Cell Rep. 2010, 29, 955–965. [Google Scholar] [CrossRef]
- Fincato, P.; Moschou, P.N.; Ahou, A.; Angelini, R.; Roubelakis-Angelakis, K.A.; Federico, R.; Tavladoraki, P. The members of Arabidopsis thaliana PAO gene family exhibit distinct tissue- and organ-specific expression pattern during seedling growth and flower development. Amino Acids 2012, 42, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.W.; Watanabe, K.; Murayama, C.; Izawa, S.; Niitsu, M.; Michael, A.J.; Berberich, T.; Kusano, T. Polyamine oxidase5 regulates Arabidopsis thaliana growth through a thermospermine oxidase activity. Plant Physiol. 2014, 165, 1575–1590. [Google Scholar] [CrossRef] [PubMed]
- Zarza, X.; Atanasov, K.E.; Marco, F.; Arbona, V.; Carrasco, P.; Kopka, J.; Fotopoulos, V.; Munnik, T.; Gómez-Cadenas, A.; Tiburcio, A.F.; et al. Polyamine oxidase 5 loss-of-function mutations in Arabidopsis thaliana trigger metabolic and transcriptional reprogramming and promote salt stress tolerance. Plant Cell Environ. 2017, 40, 527–542. [Google Scholar] [CrossRef] [PubMed]
- Kaszler, N.; Benkő, P.; Bernula, D.; Szepesi, Á.; Fehér, A.; Gémes, K. Polyamine metabolism is involved in the direct regeneration of shoots from Arabidopsis lateral root primordia. Plants 2021, 10, 305. [Google Scholar] [CrossRef]
- Wimalasekera, R.; Schaarschmidt, F.; Angelini, R.; Cona, A.; Tavladoraki, P.; Scherer, G.F. POLYAMINE OXIDASE2 of Arabidopsis contributes to ABA mediated plant developmental processes. Plant Physiol. Biochem. 2015, 96, 231–240. [Google Scholar] [CrossRef]
- Tajti, J.; Hamow, K.Á.; Majláth, I.; Gierczik, K.; Németh, E.; Janda, T.; Pál, M. Polyamine-induced hormonal changes in eds5 and sid2 mutant Arabidopsis plants. Int. J. Mol. Sci. 2019, 20, 5746. [Google Scholar] [CrossRef]
- Hunter, D.C.; Burritt, D.J. Light quality influences the polyamine content of lettuce (Lactuca sativa L.) cotyledon explants during shoot production in vitro. Plant Growth Regul. 2005, 45, 53–61. [Google Scholar] [CrossRef]
- Gondor, O.K.; Tajti, J.; Hamow, K.Á.; Majláth, I.; Szalai, G.; Janda, T.; Pál, M. Polyamine metabolism under different light regimes in wheat. Int. J. Mol. Sci. 2021, 22, 11717. [Google Scholar] [CrossRef]
- Pál, M.; Hamow, K.Á.; Rahman, A.; Majláth, I.; Tajti, J.; Gondor, O.K.; Ahres, M.; Gholizadeh, F.; Szalai, G.; Janda, T. Light spectral composition modifies polyamine metabolism in young wheat plants. Int. J. Mol. Sci. 2022, 23, 8394. [Google Scholar] [CrossRef]
- Takács, Z.; Poór, P.; Tari, I. Comparison of polyamine metabolism in tomato plants exposed to different concentrations of salicylic acid under light or dark conditions. Plant Physiol. Biochem. 2016, 108, 266–278. [Google Scholar] [CrossRef]
- Majumdar, R.; Shao, L.; Turlapati, S.A.; Minocha, S.C. Polyamines in the life of Arabidopsis: Profiling the expression of S-adenosylmethionine decarboxylase (SAMDC) gene family during its life cycle. BMC Plant Biol. 2017, 17, 264. [Google Scholar] [CrossRef] [PubMed]
- Devlin, P.F.; Halliday, K.J.; Harberd, N.P.; Whitelam, G.C. The rosette habit of Arabidopsis thaliana is dependent upon phytochrome action: Novel phytochromes control internode elongation and flowering time. Plant J. 1996, 10, 1127–1134. [Google Scholar] [CrossRef] [PubMed]
- Eskins, K. Light-quality effects on Arabidopsis development. Red, blue and far-red regulation of flowering and morphology. Physiol. Plant 1992, 86, 439–444. [Google Scholar] [CrossRef]
- Kurepin, L.V.; Walton, L.J.; Hayward, A.; Emery, R.J.N.; Pharis, R.P.; Reid, D.M. Interactions between plant hormones and light quality signaling in regulating the shoot growth of Arabidopsis thaliana seedlings. Botany 2012, 90, 237–246. [Google Scholar] [CrossRef]
- Liu, T.; Dobashi, H.; Kim, D.W.; Sagor, G.H.; Niitsu, M.; Berberich, T.; Kusano, T. Arabidopsis mutant plants with diverse defects in polyamine metabolism show unequal sensitivity to exogenous cadaverine probably based on their spermine content. Physiol. Mol. Biol. Plants 2014, 20, 151–159. [Google Scholar] [CrossRef]
- Dai, Y.R.; Galston, A.W. Simultaneous phytochrome controlled promotion and inhibition of arginine decarboxylase activity in buds and epicotyls of etiolated peas. Plant Physiol. 1981, 67, 266–269. [Google Scholar] [CrossRef]
- Yoshida, I.; Yamagata, H.; Hirasawa, E. Signal transduction controlling the blue- and red-light mediated gene expression of S-adenosylmethionine decarboxylase in Pharbitis nil. J. Exp. Bot. 2002, 53, 1525–1529. [Google Scholar]
- Yadav, A.; Singh, D.; Lingwan, M.; Yadukrishnan, P.; Masakapalli, S.K.; Datta, S. Light signaling and UV-B-mediated plant growth regulation. J. Integr. Plant Biol. 2020, 62, 1270–1292. [Google Scholar] [CrossRef]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine function in plants: Metabolism, regulation on development, and roles in abiotic stress responses. Front. Plant Sci. 2019, 9, 1945. [Google Scholar] [CrossRef]
- Sakamoto, A.; Terui, Y.; Uemura, T.; Igarashi, K.; Kashiwagi, K. Polyamines regulate gene expression by stimulating translation of histone acetyltransferase mRNAs. J. Biol. Chem. 2020, 295, 8736–8745. [Google Scholar] [CrossRef]
- Alonso, J.M.; Stepanova, A.N.; Leisse, T.J.; Kim, C.J.; Chen, H.; Shinn, P.; Stevenson, D.K.; Zimmerman, J.; Barajas, P.; Cheuk, R.; et al. Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 2003, 301, 653–657. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Jubault, M.; Hamon, C.; Gravot, A.; Lariagon, C.; Delourme, R.; Bouchereau, A.; Manzanares-Dauleux, M.J. Differential regulation of root arginine catabolism and polyamine metabolism in clubroot-susceptible and partially resistant Arabidopsis genotypes. Plant Physiol. 2008, 146, 2008–2019. [Google Scholar] [CrossRef] [PubMed]
- Kamada-Nobusada, T.; Hayashi, M.; Fukazawa, M.; Sakakibara, H.; Nishimura, M. A putative peroxisomal polyamine oxidase, AtPAO4, is involved in polyamine catabolism in Arabidopsis thaliana. Plant Cell Physiol. 2008, 49, 1272–1282. [Google Scholar] [CrossRef] [PubMed]
- Sagor, G.H.M.; Kusano, T.; Berberich, T. A polyamine oxidase from Selaginella lepidophylla (SelPAO5) can replace AtPAO5 in Arabidopsis through converting thermospermine to norspermidine instead to spermidine. Plants 2019, 8, 99. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PUT | SPD | SPM | ADC2 | SPDS2 | SPMS | CuAO1 | CuAO3 | PAO2 | PAO5 | |
|---|---|---|---|---|---|---|---|---|---|---|
| PUT | 1 | |||||||||
| SPD | −0.555 | 1 | ||||||||
| SPM | 0.915 | −0.572 | 1 | |||||||
| ADC2 | 0.688 | −0.379 | 0.617 | 1 | ||||||
| SPDS2 | −0.524 | 0.559 | −0.502 | −0.188 | 1 | |||||
| SPMS | 0.737 | −0.293 | 0.753 | 0.710 | −0.139 | 1 | ||||
| CuAO1 | 0.414 | −0.147 | 0.345 | 0.820 | −0.095 | 0.553 | 1 | |||
| CuAO3 | 0.169 | −0.124 | 0.159 | 0.413 | 0.130 | 0.261 | 0.410 | 1 | ||
| PAO2 | 0.834 | −0.593 | 0.829 | 0.688 | −0.431 | 0.666 | 0.470 | 0.399 | 1 | |
| PAO5 | 0.469 | −0.500 | 0.423 | 0.157 | −0.135 | 0.334 | −0.050 | 0.260 | 0.423 | 1 |
| Treatment | Intensity PAR (µmol m−2 s−1) | Blue µW/cm2 (400–500 nm) | Green µW/cm2 (500–600 nm) | Red µW/cm2 (600–700 nm) | Far-red µW/cm2 (700–800 nm) | Blue/ Red | Red/ Far-Red | Blue% | Green% | Red% | Far-Red% |
|---|---|---|---|---|---|---|---|---|---|---|---|
| L1 | 100 | 400 | 640 | 1020 | 30 | 0.39 | 34 | 19.14 | 30.62 | 48.8 | 1.44 |
| L2 | 100 | 450 | 680 | 930 | 240 | 0.48 | 3.88 | 19.57 | 29.57 | 40.43 | 10.43 |
| L3 | 100 | 890 | 650 | 690 | 30 | 1.29 | 23 | 39.38 | 28.76 | 30.53 | 1.33 |
| Gene Name | Primer Sequences (5′ → 3′) | Amplicon Size (bp) | References | |
|---|---|---|---|---|
| AtActin8 | forward | TTACCCGACGGACAAGTGATC | 73 | [53] |
| reverse | ATGATGGCTGGAAAAGGACTTC | |||
| AtADC2 | forward | GCGATGGACCACACAGCTTT | 64 | |
| reverse | AGAACATCCGCTGAGGACTGA | |||
| AtSPDS2 | forward | TTGCCCGTGAAGAGACCTAGA | 72 | |
| reverse | TCCACCGTTCTCTGTTTCCAT | |||
| AtSPMS | forward | TGGCTCCATACTCATCTTATTGAA | 72 | |
| reverse | CGCATAGTGAACACTTTTGAATG | |||
| AtPAO2 | forward | GGAATGCCGGAAGATCTTCCGTGATTGTGATCGG | 142 | [54] |
| reverse | CGATTCCAACACCGAGATTTGCATACTCCATGCAGC | |||
| AtPAO5 | forward | GTTGGGATGAACCAGAAGGA | 132 | [55] |
| reverse | GAGGAGCCTCGGTAAGAAGA | |||
| AtCuAO1 | forward | AGCTGGCGACATTCTGAGAT | 238 | [29] |
| reverse | GTCCAGCATCATCCTCCCTA | |||
| AtCuAO3 | forward | GTAAGTTTGTGCCACTCCCCC | 153 | |
| reverse | GCCACTCGACAAAGTACCCCC | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, A.; Tajti, J.; Majláth, I.; Janda, T.; Prerostova, S.; Ahres, M.; Pál, M. Influence of a phyA Mutation on Polyamine Metabolism in Arabidopsis Depends on Light Spectral Conditions. Plants 2023, 12, 1689. https://doi.org/10.3390/plants12081689
Rahman A, Tajti J, Majláth I, Janda T, Prerostova S, Ahres M, Pál M. Influence of a phyA Mutation on Polyamine Metabolism in Arabidopsis Depends on Light Spectral Conditions. Plants. 2023; 12(8):1689. https://doi.org/10.3390/plants12081689
Chicago/Turabian StyleRahman, Altafur, Judit Tajti, Imre Majláth, Tibor Janda, Sylva Prerostova, Mohamed Ahres, and Magda Pál. 2023. "Influence of a phyA Mutation on Polyamine Metabolism in Arabidopsis Depends on Light Spectral Conditions" Plants 12, no. 8: 1689. https://doi.org/10.3390/plants12081689
APA StyleRahman, A., Tajti, J., Majláth, I., Janda, T., Prerostova, S., Ahres, M., & Pál, M. (2023). Influence of a phyA Mutation on Polyamine Metabolism in Arabidopsis Depends on Light Spectral Conditions. Plants, 12(8), 1689. https://doi.org/10.3390/plants12081689