



Shading Stress at Different Grain Filling Stages Affects Dry Matter and Nitrogen Accumulation and Remobilization in Fresh Waxy Maize



Abstract
:1. Introduction
2. Results
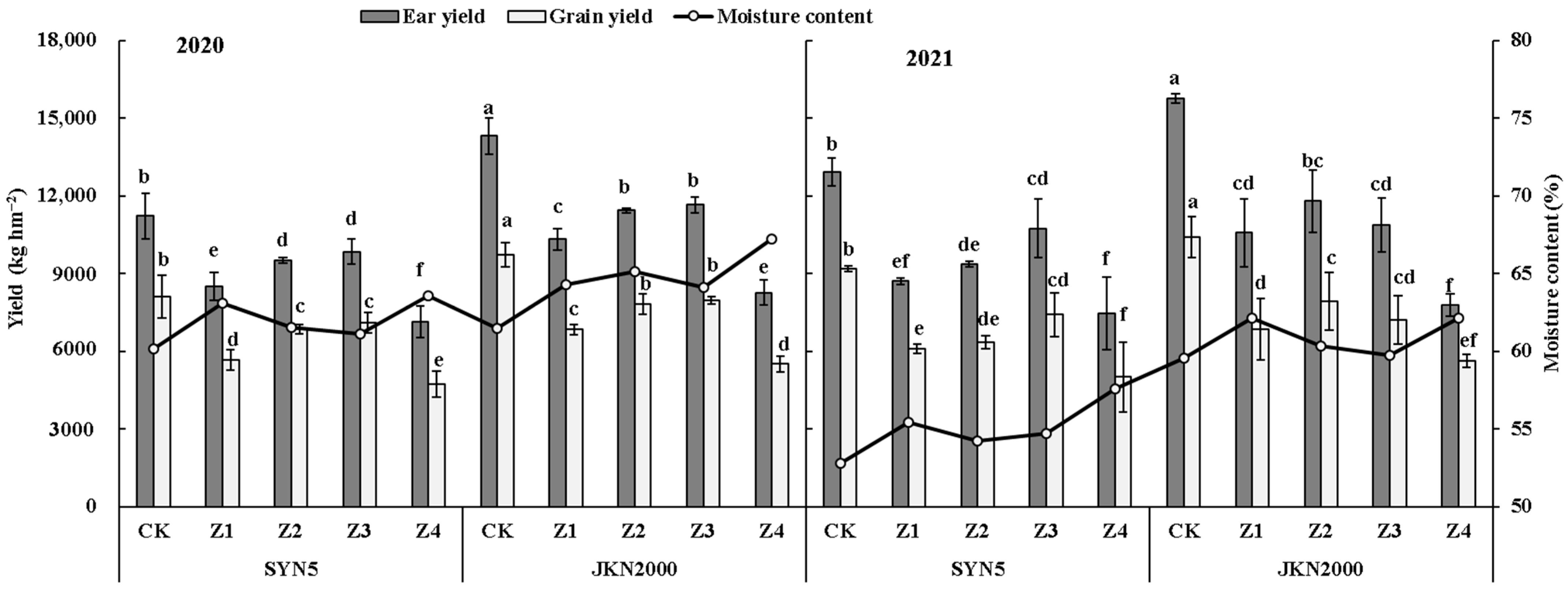
2.1. Yield
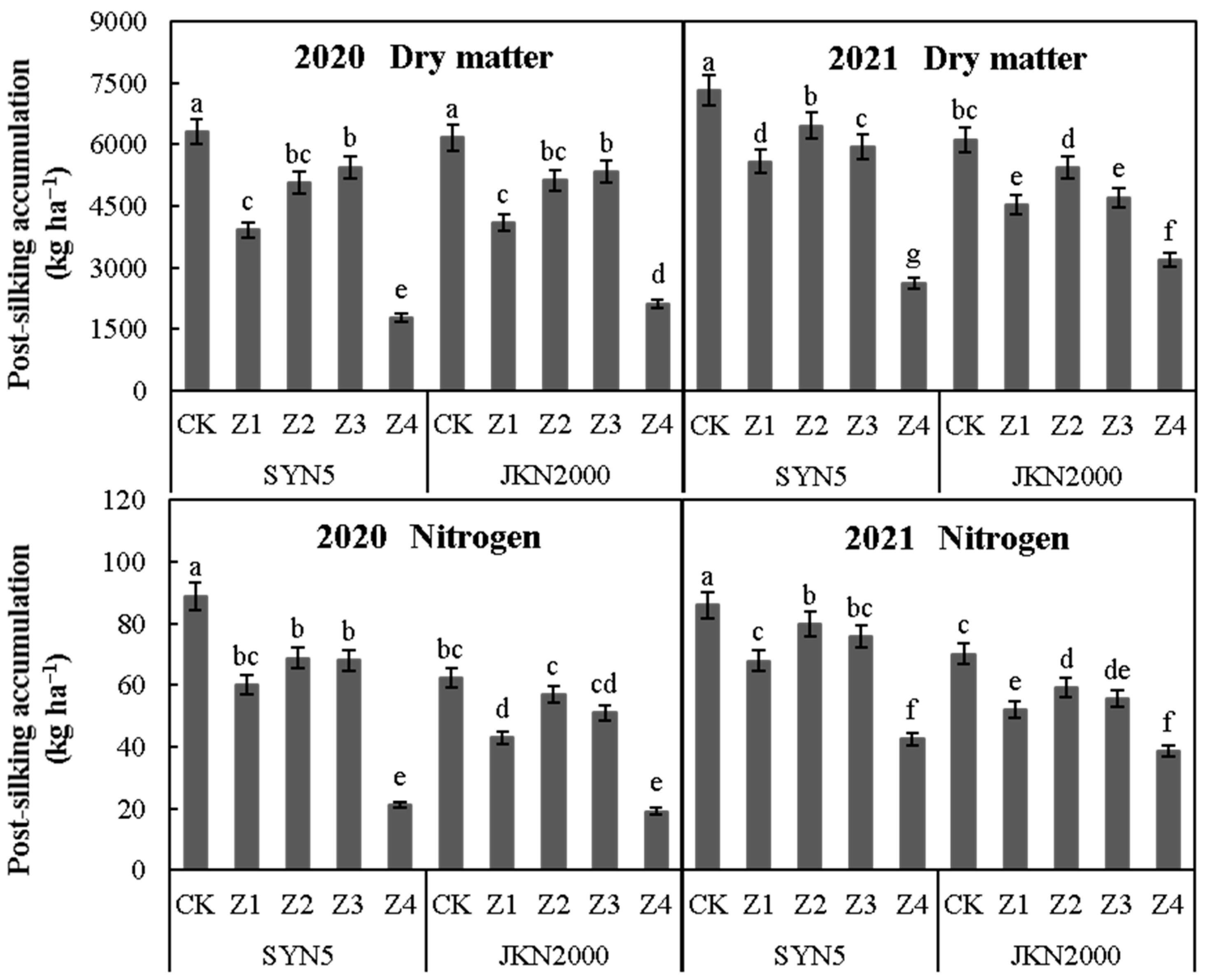
2.2. Dry Matter and Nitrogen Accumulation at Post-Silking
2.3. Post-Silking Remobilization of Dry Matter and Nitrogen
2.4. Pre-Silking Remobilization of Dry Matter and Nitrogen
2.5. Harvest Index of Dry Matter and Nitrogen
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Sampling and Measurements
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, I.; Swaminathan, K.; Hudson, K.; Hudson, M. Evolutionary divergence of phytochrome protein function in Zea mays PIF3 signaling. J. Exp. Bot. 2016, 67, 4231–4240. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.X.; Wang, L.C.; Niu, Z.G.; Zhang, M.; Li, C.A.; Li, J.R. The effects of projected climate change and extreme climate on maize and rice in the Yangtze River Basin, China. Agric. For. Meteorol. 2020, 282–283, 107867. [Google Scholar] [CrossRef]
- Shi, K.; Gu, X.; Lu, W.; Lu, D. Effects of weak-light stress during grain filling on the physicochemical properties of normal maize starch. Carbohyd. Polym. 2018, 202, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.S.; Guo, X.X.; Liu, G.Z.; Liu, W.M.; Xue, J.; Ming, B.; Xie, R.Z.; Wang, K.R.; Hou, P.; Li, S.K. Solar radiation effects on dry matter accumulations and transfer in maize. Front. Plant Sci. 2021, 12, 727134. [Google Scholar] [CrossRef]
- Gerakis, P.; Papakosta-Tasopoulou, D. Effects of dense planting and artificial shading on five maize hybrids. Agric. Meteorol. 1980, 2, 129–137. [Google Scholar] [CrossRef]
- Mbewe, D.; Hunter, R.B. The effect of shade stress on the performance of corn for silage versus grain. Can. J. Plant Sci. 1986, 1, 53–60. [Google Scholar] [CrossRef]
- Jia, S.f.; Li, C.; Dong, S.T.; Zhang, J.W. Effects of shading at different stages after anthesis on maize grain weight and quality at cytology level. J. Integr. Agric. 2011, 10, 58–69. [Google Scholar] [CrossRef]
- Gao, J.; Zhao, B.; Dong, S.T.; Liu, P.; Ren, B.Z.; Zhang, J.W. Response of summer maize photosynthate accumulation and distribution to shading stress assessed by using 13CO2 stable isotope tracer in the field. Front. Plant Sci. 2017, 8, 1821. [Google Scholar] [CrossRef]
- Cui, H.Y.; Camberato, J.; Jin, L.B.; Zhang, J.W. Effects of shading on spike differentiation and grain yield formation of summer maize in the field. Int. J. Biometeorol. 2015, 59, 1189–1200. [Google Scholar] [CrossRef]
- Wen, Z.; Shi, K.; Lu, W.P.; Lu, D.L. Effects of postsilking weak-light stress on the flour quality of spring maize. Cereal Chem. 2019, 96, 742–753. [Google Scholar] [CrossRef]
- Lu, D.; Cai, X.M.; Xu, R.; Zhao, J.; Lu, W.P. Effects of shading after pollination on starch physico-chemical properties of waxy maize. Res. Crops 2014, 15, 49–54. [Google Scholar] [CrossRef]
- Bellasio, C.; Griffiths, H. Acclimation of C4 metabolism to low light in mature maize leaves could limit energetic losses during progressive shading in a crop canopy. J. Exp. Bot. 2014, 13, 3725–3736. [Google Scholar] [CrossRef] [PubMed]
- Ubierna, N.; Sun, W.; Kramer, D.; Cousins, A. The efficiency of C4 photosynthesis under low light conditions in Zea mays L, Miscanthus×giganteus and Flaveria bidentis. Plant Cell Environ. 2013, 36, 365–381. [Google Scholar] [CrossRef]
- Ren, B.; Cui, H.Y.; Camberato, J.; Dong, S.T.; Liu, P.; Zhao, B.; Zhang, J.W. Effects of shading on the photosynthetic characteristics and mesophyll cell ultrastructure of summer maize. Sci. Nat. 2016, 103, 67. [Google Scholar] [CrossRef] [PubMed]
- Sharwood, R.E.; Sonawane, B.; Ghannoum, O. Photosynthetic flexibility in maize exposed to salinity and shade. J. Exp. Bot. 2014, 65, 3715–3724. [Google Scholar] [CrossRef]
- Zhong, X.; Shi, Z.; Li, F.; Huang, H. Photosynthesis and chlorophyll fluorescence of infertile and fertile stalks of paired near-isogenic lines in maize (Zea mays L.) under shade conditions. Photosynthetica 2014, 52, 597–603. [Google Scholar] [CrossRef]
- Impa, S.M.; Sunoj, V.S.J.; Krassovskaya, I.; Bheemanahalli, R.; Obata, T.; Jagadish, S.V.K. Carbon balance and source-sink metabolic changes in winter wheat exposed to high night-time temperature. Plant Cell Environ. 2019, 42, 1233–1246. [Google Scholar] [CrossRef]
- Tang, W.; Cheng, W.X.; Zeng, H.; Zhu, B. Light intensity controls rhizosphere respiration rate and rhizosphere priming effect of soybean and sunflower. Rhizosphere 2019, 9, 97–105. [Google Scholar] [CrossRef]
- Merah, O.; Monneveux, P. Contribution of different organs to grain filling in durum wheat under mediterranean conditions I. contribution of post-anthesis photosynthesis and remobilization. J. Agron. Crop Sci. 2015, 201, 344–352. [Google Scholar] [CrossRef]
- Sun, Z.C.; Zhang, J.W. Physiological mechanism and regulation effect of low light on maize yield formation. Acta Agron. Sin. 2023, 49, 12–23, (In Chinese with English Abstract). [Google Scholar]
- Yuan, L.Z.; Tang, J.H.; Liu, J.Y.; Song, H.; Zhang, M.B.; Li, H.P.; Li, C.H. Differential miRNA expression in maize ear subjected to shading tolerance. Acta Physiol. Plant 2016, 38, 80. [Google Scholar] [CrossRef]
- Deng, F.; Zeng, Y.L.; Li, Q.P.; He, C.Y.; Li, B.; Zhu, Y.Y.; Zhou, X.; Yang, F.; Zhong, X.Y.; Wang, L.; et al. Decreased anther dehiscence contributes to a lower fertilization rate of rice subjected to shading stress. Field Crops Res. 2021, 273, 108291. [Google Scholar] [CrossRef]
- Gao, J.; Shi, J.G.; Dong, S.T.; Liu, P.; Zhao, B.; Zhang, J.W. Grain development and endogenous hormones in summer maize (Zea mays L.) submitted to different light conditions. Int. J. Biometeorol. 2018, 62, 2131–2138. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; An, P.P.; Li, C.H. Ovary abortion induced by combined waterlogging and shading stress at the flowering stage involves amino acids and flavonoid metabolism in maize. Front. Plant Sci. 2021, 12, 778717. [Google Scholar]
- Barnabás, B.; Jager, K.; Fehér, A. The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ. 2008, 31, 11–38. [Google Scholar] [CrossRef]
- Yang, H.; Shi, Y.; Xu, R.; Lu, D.; Lu, W. Effects of shading after pollination on kernel filling and physicochemical quality traits of waxy maize. Crop J. 2016, 4, 235–245. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Li, G.H.; Yang, H.; Lu, D.L. Foliar brassinolide sprays ameliorate post-silking heat stress on the accumulation and remobilization of biomass and nitrogen in fresh waxy maize. Agronomy 2022, 12, 1363. [Google Scholar] [CrossRef]
- Li, G.H.; Li, W.; Zhang, S.B.; Lu, W.P.; Lu, D.L. Optimized fertilization practices improved rhizosphere soil chemical and bacterial properties and fresh waxy maize yield. Metabolites 2022, 12, 935. [Google Scholar] [CrossRef]
- Hannachi, L.; Deléens, E.; Gate, P. Nitrogen and carbon isotope composition of wheat grain: Alteration due to sink-source modifications at flowering. Rapid Commun. Mass Spectrom. 1996, 19, 979–986. [Google Scholar] [CrossRef]
- Yang, J.C.; Zhang, J.H. Grain filling of cereals under soil drying. New Phytol. 2006, 169, 223–236. [Google Scholar] [CrossRef]
- Xu, Y.M.; Zhang, X.Y.; Yang, H.; Lu, D.L. Effects of exogenous brassinolide application at the silking stage on nutrient accumulation, translocation and remobilization of waxy maize under post-silking heat stress. Agriculture 2022, 12, 572. [Google Scholar] [CrossRef]
- Takayuki, Y.; Osamu, U. Structural and photosynthetic reacclimation to low light in C4 maize leaves that developed under high light. Ann. Bot. 2019, 124, 437–445. [Google Scholar]
- Lu, D.L.; Sun, X.L.; Wang, X.; Yan, F.B.; Lu, W.P. Effect of shading during grain filling on the physicochemical properties of fresh waxy maize. J. Integr. Agric. 2013, 12, 1560–1567. [Google Scholar] [CrossRef]
- Wang, J.; Shi, K.; Lu, W.P.; Lu, D.L. Effects of post-silking shading stress on enzymatic activities and phytohormone contents during grain development in spring Maize. J. Plant Growth Regul. 2021, 40, 1060–1073. [Google Scholar] [CrossRef]
- Gao, Z.; Sun, L.; Ren, J.H.; Liang, X.G.; Shen, S.; Lin, S.; Zhao, X.; Chen, X.M.; Wu, G.; Zhou, S.L. Detasseling increases kernel number in maize under shade stress. Agric. For. Meteorol. 2020, 280, 107811. [Google Scholar] [CrossRef]
- Chen, Y.L.; Xiao, C.X.; Chen, X.C.; Li, Q.; Zhang, J.; Chen, F.J.; Yuan, L.X.; Mi, G.H. Characterization of the plant traits contributed to high grain yield and high grain nitrogen concentration in maize. Field Crops Res. 2014, 159, 1–9. [Google Scholar] [CrossRef]
- Zhou, T.; Wang, L.; Sun, X.; Wang, X.C.; Chen, Y.L.; Rengel, Z.; Liu, W.G.; Yang, W.Y. Light intensity influence maize adaptation to low P stress by altering root morphology. Plant Soil 2020, 447, 183–197. [Google Scholar] [CrossRef]
- Yu, P.; He, X.M.; Baer, M.; Beirinckx, S.; Tian, T.; Moya, Y.A.; Zhang, X.C.; Deichmann, M.; Frey, F.P.; Bresgen, V.; et al. Plant flavones enrich rhizosphere oxalobacteraceae to improve maize performance under nitrogen deprivation. Nat. Plants 2021, 7, 481–499. [Google Scholar] [CrossRef]
- Wang, J.; Shi, K.; Lu, W.P.; Lu, D.L. Post-silking shading stress affects leaf nitrogen metabolism of spring maize in southern China. Plants 2020, 9, 210. [Google Scholar] [CrossRef]
- Majlath, I.; Darko, E.; Palla, B.; Nagy, Z.; Janda, T.; Szalai, G. Reduced light and moderate water deficiency sustain nitrogen assimilation and sucrose degradation at low temperature in durum wheat. J. Plant Physiol. 2016, 191, 149–158. [Google Scholar] [CrossRef]
- Yu, S.; Wang, Y.; Zhou, Z.; Lv, F.; Liu, J.; Ma, Y.; Chen, J.; Abudukeyoumu, A. Effect of shade on nitrogen metabolism and its mechanism in cotton plant at flowering andboll-forming stage. Acta Agron. Sin. 2011, 37, 1879–1887. [Google Scholar] [CrossRef]
- Shu, S.; Tang, Y.Y.; Yuan, Y.H.; Sun, J.; Zhong, M.; Guo, S.R. The role of 24-epibrassinolide in the regulation of photosynthetic characteristics and nitrogen metabolism of tomato seedlings under a combined low temperature and weak light stress. Plant Physiol. Biochem. 2016, 107, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, T.R.; Pinter, P.J.; Kimball, B.A.; Adamsen, F.J.; LaMorte, R.L.; Wall, G.W.; Hunsaker, D.J.; Adam, N.; Brooks, T.J.; Garcia, R.L.; et al. Leaf nitrogen concentration of wheat subjected to elevated CO2 and either water or N deficits. Agric. Ecosyst. Environ. 2000, 79, 53–60. [Google Scholar] [CrossRef]
- Chen, K.; Kumudini, S.V.; Tollenaar, M.; Vyn, T.J. Plant biomass and nitrogen partitioning changes between silking and maturity in newer versus older maize hybrids. Field Crops Res. 2015, 183, 315–328. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Variety | Shading Treatment | Stem | Leaf | Total | |||
|---|---|---|---|---|---|---|---|---|
| REP (kg ha−1) | REE (%) | REP (kg ha−1) | REE (%) | REP (kg ha−1) | REE (%) | |||
| 2020 | SYN5 | CK | −458.8 h | −13.6 f | −591.6 h | −49.8 f | −1050.4 j | −23.1 f |
| Z1 | 332.8 c | 9.9 c | −359.0 f | −30.2 d | −26.2 f | −0.6 c | ||
| Z2 | 106.2 d | 3.2 d | −481.2 g | −40.5 e | −375.0 h | −8.2 d | ||
| Z3 | −38.6 g | −1.1 e | −601.5 h | −50.6 f | −640.1 i | −14.0 e | ||
| Z4 | 1149.4 a | 34.1 a | −274.8 e | −23.1 c | 874.6 b | 19.2 a | ||
| JKN2000 | CK | 4.6 f | 0.1 e | −103.6 d | −4.9 b | −99.0 g | −1.6 c | |
| Z1 | 372.2 c | 9.0 c | 102.4 a | 4.9 a | 474.6 c | 7.6 b | ||
| Z2 | 72.8 e | 1.8 de | −76.2 c | −3.6 b | −3.4 e | −0.1 c | ||
| Z3 | 332.4 c | 8.1 c | −95.6 d | −4.5 b | 236.8 d | 3.8bc | ||
| Z4 | 962.2 b | 23.4 b | 20.6 b | 1.0 ab | 982.8 a | 15.8 a | ||
| 2021 | SYN5 | CK | −1435.6 h | −52.0 e | −535.8 g | −38.0 de | −1971.4 j | −47.3 f |
| Z1 | −564.6 f | −20.5 d | −377.4 e | −26.7 cd | −942.0 g | −22.6 d | ||
| Z2 | −697.2 g | −25.3 d | −660.2 h | −46.8 e | −1357.4 i | −32.6 e | ||
| Z3 | −705.0 g | −25.6 d | −458.2 f | −32.5 d | −1163.2 h | −27.9 de | ||
| Z4 | 378.2 b | 13.7 a | −174.8 b | −12.4 b | 203.4 b | 4.9ab | ||
| JKN2000 | CK | −46.4 d | −1.2 bc | −275.8 d | −13.3 b | −322.2 e | −5.3 bc | |
| Z1 | −26.2 d | −0.7 bc | −238.6 c | −11.5 b | −264.8 d | −4.4 bc | ||
| Z2 | −139.0 e | −3.5 c | −423.2 f | −20.4 c | −562.2 f | −9.3 c | ||
| Z3 | 123.0 c | 3.1 b | −252.4 cd | −12.2 b | −129.4 c | −2.1 b | ||
| Z4 | 495.2 a | 12.5 a | −67.6 a | −3.3 a | 427.6 a | 7.1 a | ||
| ANOVA | ||||||||
| Year (Y) | 301.2 ** | 357.2 ** | 307.7 ** | 20.5 ** | 115.6 ** | 106.4 ** | ||
| Variety (V) | 162.7 ** | 205.1 ** | 296.8 ** | 191.9 ** | 145.3 ** | 137.1 ** | ||
| Shading (S) | 222.5 ** | 478.4 ** | 674.2 ** | 147.5 ** | 374.3 ** | 282.9 ** | ||
| Y × V | 84.8 ** | 149.3 ** | 47.4 ** | 28.2 ** | 68.4 ** | 184.3 ** | ||
| Y × S | 25.7 ** | 40.5 ** | 165.4 ** | 27.4 ** | 77.2 ** | 45.0 ** | ||
| V × S | 40.3 ** | 127.4 ** | 54.7 ** | 31.7 ** | 53.8 ** | 68.7 ** | ||
| Y × V × S | 29.4 ** | 167.1 ** | 12.8 ** | 39.5 ** | 40.6 ** | 49.9 ** | ||
| Year | Variety | Shading Treatment | Stem | Leaf | Total | |||
|---|---|---|---|---|---|---|---|---|
| REP (kg ha−1) | REE (%) | REP (kg ha−1) | REE (%) | REP (kg ha−1) | REE (%) | |||
| 2020 | SYN5 | CK | −17.1 f | −72.1 g | −8.3 bc | −37.5 f | −25.4 e | −55.5 f |
| Z1 | −8.0 e | −33.5 f | −8.1 bc | −36.7 f | −16.0 d | −35.0 e | ||
| Z2 | −8.7 e | −36.5 f | −8.1 bc | −37.0 f | −16.8 d | −36.7 e | ||
| Z3 | −4.3 d | −18.2 e | −9.7 c | −44.2 g | −14.1 d | −30.7 d | ||
| Z4 | 11.7 ab | 49.1 a | −5.2 b | −23.6 e | 6.5 b | 14.1 ab | ||
| JKN2000 | CK | 11.3 ab | 39.0 b | −3.7 b | −9.5 c | 7.7 b | 11.2 b | |
| Z1 | 5.8 c | 19.9 d | 2.8 a | 7.1 a | 8.6 b | 12.6 b | ||
| Z2 | 5.3 c | 18.3 d | −5.6 b | −14.4 d | −0.3 c | −0.5 c | ||
| Z3 | 8.4 b | 28.9 c | 0.4 ab | 1.1 b | 8.8 b | 13.0 b | ||
| Z4 | 13.5 a | 46.5 ab | −0.6 ab | −1.5 b | 12.9 a | 19.0 a | ||
| 2021 | SYN5 | CK | −13.1 de | −63.7 f | −7.1 b | −26.9 d | −20.2 e | −43.0 f |
| Z1 | −7.6 cd | −37.1 d | −7.3 b | −27.7 d | −15.0 d | −31.8 e | ||
| Z2 | −10.0 d | −48.6 e | −10.5 c | −39.8 e | −20.5 e | −43.7 f | ||
| Z3 | −14.8 e | −71.8 g | −10.0 c | −37.7 e | −24.8 f | −52.6 g | ||
| Z4 | −4.9 c | −23.6 c | −3.4 ab | −13.0 b | −8.3 c | −17.7 d | ||
| JKN2000 | CK | 7.7 a | 26.1 a | −7.9 b | −20.3 c | −0.2 b | −0.3 b | |
| Z1 | 5.3 b | 17.8 b | −4.7 ab | −12.0 b | 0.6 b | 0.9 b | ||
| Z2 | 5.9 ab | 20.1 ab | −7.9 b | −20.4 c | −2.0 bc | −2.9 c | ||
| Z3 | 7.0 a | 23.7 a | −7.5 b | −19.3 c | −0.5 b | −0.8 b | ||
| Z4 | 7.5 a | 25.4 a | 1.0 a | 2.5 a | 8.5 a | 12.4 a | ||
| ANOVA | ||||||||
| Year (Y) | 61.9 ** | 66.6 ** | 261.8 ** | 7.9 * | 120.1 ** | 100.7 ** | ||
| Variety (V) | 117.9 ** | 83.3 ** | 127.9 ** | 138.7 ** | 153.9 ** | 171.6 ** | ||
| Shading (S) | 213.8 ** | 180.9 ** | 239.0 ** | 237.4 ** | 415.1 ** | 349.5 ** | ||
| Y × V | 8.1 * | 19.3 ** | 30.9 ** | 196.7 ** | 2.6 | 0.1 | ||
| Y × S | 84.9 ** | 61.8 ** | 48.3 ** | 33.4 ** | 48.4 ** | 49.5 ** | ||
| V × S | 130.9 ** | 134.9 ** | 52.3 ** | 46.7 ** | 97.3 ** | 121.4 ** | ||
| Y × V × S | 45.3 ** | 35.3 ** | 42.9 ** | 34.0 ** | 65.3 ** | 64.0 ** | ||
| Date (Month/Date) | Light Intensity (Lux) | Photosynthetically Active Radiation (μmol m−2 s−1) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CK | Z1 | Z2 | Z3 | Z4 | CK | Z1 | Z2 | Z3 | Z4 | |
| 6.9 | 100,800 | 49,236 | 101,100 | 100,456 | 49,736 | 1355 | 670 | 1286 | 1397 | 580 |
| 6.10 | 85,312 | 48,536 | 84,312 | 82,312 | 49,577 | 1059 | 486 | 1100 | 1065 | 512 |
| 6.11 | 84,352 | 54,562 | 83,352 | 82,352 | 53,562 | 1137 | 523 | 1037 | 1237 | 557 |
| 6.12 | 123,008 | 66,637 | 122,008 | 121,008 | 65,637 | 1897 | 845 | 1765 | 1812 | 893 |
| 6.13 | 92,148 | 60,054 | 91,148 | 93,148 | 59,054 | 1391 | 712 | 1223 | 1308 | 693 |
| 6.14 | 63,008 | 29,617 | 62,008 | 64,008 | 27,617 | 906 | 357 | 880 | 912 | 445 |
| 6.15 | 121,512 | 57,617 | 120,512 | 122,512 | 56,617 | 1900 | 865 | 1800 | 185 | 905 |
| 6.16 | 67,637 | 66,637 | 21,562 | 65,637 | 24,562 | 945 | 913 | 435 | 925 | 392 |
| 6.17 | 66,066 | 65,066 | 30,916 | 64,066 | 33,916 | 1020 | 920 | 535 | 1107 | 525 |
| 6.18 | 76,719 | 75,719 | 41,047 | 74,719 | 42,912 | 1213 | 1113 | 639 | 1013 | 607 |
| 6.19 | 73,706 | 72,706 | 34,772 | 71,706 | 33,701 | 1186 | 1086 | 506 | 1114 | 570 |
| 6.20 | 49,636 | 48,636 | 19,899 | 47,636 | 19,074 | 776 | 758 | 341 | 726 | 328 |
| 6.21 | 60,181 | 59,181 | 21,621 | 58,181 | 20,621 | 955 | 946 | 396 | 925 | 402 |
| 6.22 | 81,378 | 80,378 | 42,374 | 79,378 | 43,558 | 1297 | 1182 | 587 | 1306 | 640 |
| 6.23 | 2582 | 2475 | 2260 | 1323 | 1681 | 35 | 36 | 38 | 7 | 6 |
| 6.24 | 83,099 | 82,099 | 81,099 | 42,299 | 41,015 | 1315 | 1150 | 1320 | 620 | 632 |
| 6.25 | 84,713 | 83,713 | 82,713 | 41,976 | 40,763 | 1275 | 1391 | 1296 | 621 | 585 |
| 6.26 | 88,878 | 89,878 | 87,878 | 43,020 | 48,005 | 1390 | 1253 | 1274 | 611 | 578 |
| 6.27 | 39,629 | 38,629 | 40,629 | 17,887 | 18,608 | 687 | 703 | 673 | 272 | 295 |
| 6.28 | 90,492 | 89,492 | 88,492 | 36,431 | 38,794 | 1597 | 1471 | 1538 | 714 | 740 |
| 6.29 | 99,271 | 98,271 | 97,271 | 38,970 | 37,579 | 1478 | 1434 | 1326 | 660 | 671 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, H.; Li, W.; Liang, Y.; Li, G. Shading Stress at Different Grain Filling Stages Affects Dry Matter and Nitrogen Accumulation and Remobilization in Fresh Waxy Maize. Plants 2023, 12, 1742. https://doi.org/10.3390/plants12091742
Sun H, Li W, Liang Y, Li G. Shading Stress at Different Grain Filling Stages Affects Dry Matter and Nitrogen Accumulation and Remobilization in Fresh Waxy Maize. Plants. 2023; 12(9):1742. https://doi.org/10.3390/plants12091742
Chicago/Turabian StyleSun, Haohan, Wei Li, Yuwen Liang, and Guanghao Li. 2023. "Shading Stress at Different Grain Filling Stages Affects Dry Matter and Nitrogen Accumulation and Remobilization in Fresh Waxy Maize" Plants 12, no. 9: 1742. https://doi.org/10.3390/plants12091742
APA StyleSun, H., Li, W., Liang, Y., & Li, G. (2023). Shading Stress at Different Grain Filling Stages Affects Dry Matter and Nitrogen Accumulation and Remobilization in Fresh Waxy Maize. Plants, 12(9), 1742. https://doi.org/10.3390/plants12091742