

Epipactis bucegensis—A Separate Autogamous Species within the E. helleborine Alliance

 , ,
, ,
Abstract
:1. Introduction
2. Results
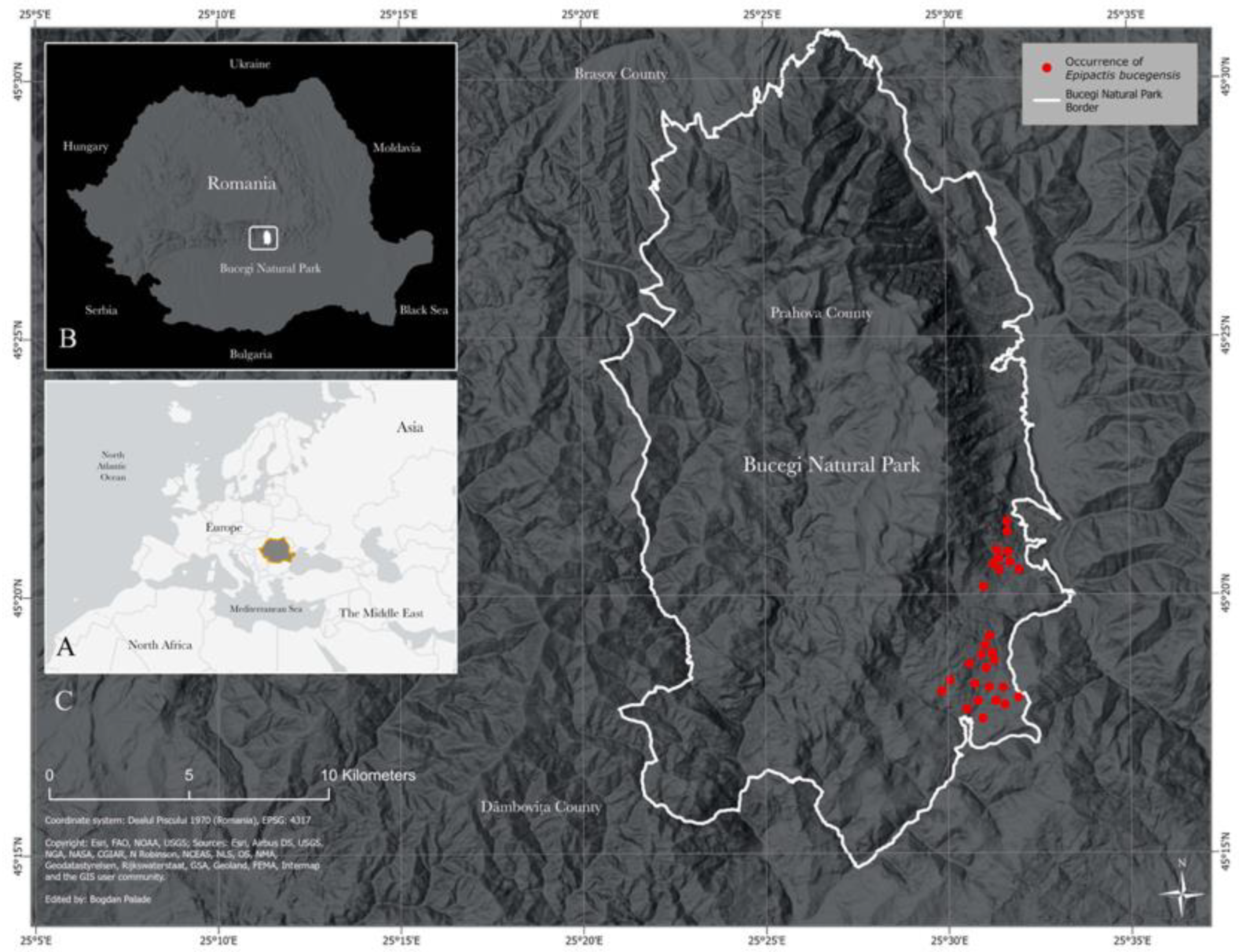
2.1. Sites Studied
2.2. Morphological Comparisons
2.3. Morphological Distinctness of Epipactis bucegensis
2.4. Morphological Changes to Autogamy
2.5. Pollination Monitoring
3. Discussion
3.1. Active Speciation within the Epipactis Genus
3.2. Inbreeding—Friend or Foe?
3.3. The Role of Cleistogamy in Active Speciation
4. Materials and Methods
4.1. Sites Studied
4.2. Populations Counts
4.3. Extent of Occurrence (EOO)
4.4. Species Studied
4.5. Study Time Frames
4.6. Morphological Comparisons
4.7. Pollination Monitoring
4.8. Digital Photographic Equipment
4.9. Maps
5. Conclusions
6. Taxonomic Treatment
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Wood, J. Epipactis. Distribution. In Genera Orchidacearum Volume 4: Epidendroideae (Part 1); Pridgeon, A.M., Cribb, P., Chase, M.W., Rasmussen, F.N., Eds.; Oxford University Press: Oxford, UK, 2005; Volume 4, pp. 501–503. ISBN 978-0-19-850712-3. [Google Scholar]
- Richards, A.J. The Influence of Minor Structural Changes in the Flower on Breeding Systems and Speciation in Epipactis Zinn (Orchidaceae). Pollinat. Evol. 1982, 47–53. [Google Scholar]
- Delforge, P. Orchids of Britain and Europe, 1st ed.; Harper Collins: London, UK, 1995; ISBN 978-0-00-220024-0. [Google Scholar]
- Delforge, P. Guide des Orchidees d’Europe, d’Afrique du Nord et du Proche-Orient, 1st ed.; Delachaux & Niestle: Librairie Le Square Grenoble, France, 1994; ISBN 978-2-603-00929-1. [Google Scholar]
- Delforge, P. Orchids of Europe, North Africa and the Middle East, 3rd ed.; Harrap, S., Ed.; A. & C. Black: London, UK, 2006; ISBN 978-0-7136-7525-2. [Google Scholar]
- Delforge, P. Guide des Orchidées d’Europe, d’Afrique du Nord et du Proche-Orient, 3rd ed.; Delachaux et Niestlé: Paris, France, 2005; ISBN 978-2-603-01323-6. [Google Scholar]
- Delforge, P. Orchidées d’Europe (Nvelle éd): dAfrique du Nord et du Proche-Orient, 4th ed.; Delachaux et Niestlé: Paris, France, 2016; ISBN 978-2-603-02407-2. [Google Scholar]
- Kühn, R.; Pedersen, H.; Cribb, P. Field Guide to the Orchids of Europe and the Mediterranean; Kew Publishing, Royal Botanic Gardens: London, UK, 2019; ISBN 978-1-84246-669-8. [Google Scholar]
- Griebl, N.; Presser, H. Orchideen-Europas; Franckh-Kosmos Verlags-GmbH & Company KG: Stuttgart, Germany, 2021; ISBN 978-3-440-50419-2. [Google Scholar]
- AHO-Bayern, e.V. Die Gattung Epipactis. Available online: https://www.aho-bayern.de/epipactis/fs_epipactis_1.html (accessed on 31 March 2023).
- Jakubska-Busse, A.; Gola, E.M. Validation of Leaf Undulation Traits in the Taxonomy of Epipactis muelleri Godfery, 1921 (Orchidaceae, Neottieae). Plant Syst. Evol. 2014, 300, 1707–1717. [Google Scholar] [CrossRef]
- Rewicz, A.; Rewers, M.; Jędrzejczyk, I.; Rewicz, T.; Kołodziejek, J.; Jakubska-Busse, A. Morphology and Genome Size of Epipactis helleborine (L.) Crantz (Orchidaceae) Growing in Anthropogenic and Natural Habitats. PeerJ 2018, 6, e5992. [Google Scholar] [CrossRef] [PubMed]
- Kolanowska, M. Niche Conservatism and the Future Potential Range of Epipactis helleborine (Orchidaceae). PLoS ONE 2013, 8, e77352. [Google Scholar] [CrossRef]
- Squirrell, J.; Hollingsworth, P.M.; Bateman, R.M.; Dickson, J.H.; Light, M.H.; Macconaill, M.; Tebbitt, M.C. Partitioning and Diversity of Nuclear and Organelle Markers in Native and Introduced Populations of Epipactis helleborine (Orchidaceae). Am. J. Bot. 2001, 88, 1409–1418. [Google Scholar] [CrossRef]
- Light, M.H.S.; MacConaill, M. Appearance and Disappearance of a Weedy Orchid, Epipactis helleborine. Folia Geobot. 2006, 41, 77–93. [Google Scholar] [CrossRef]
- Fateryga, A.V.; Fateryga, V.V. The genus Epipactis Zinn (Orchidaceae) in the flora of Russia. Turczaninowia 2018, 21, 19–34. [Google Scholar] [CrossRef]
- Molnár, A.V.; Sramkó, G. Epipactis albensis (Orchidaceae): A New Species in the Flora of Romania. Biologia 2012, 67, 883–888. [Google Scholar] [CrossRef]
- Rasmussen, H.N. Terrestrial Orchids: From Seed to Mycotrophic Plant; Cambridge University Press (Transfered to digital printing 2003): Cambridge, UK, 1995; ISBN 978-0-521-45165-9. [Google Scholar]
- De Angelli, N.; Anghelescu, D. Orchids of Romania, 1st ed.; Rotolito Romania SA: Pantelimon, Romania, 2020; ISBN 978-973-0-32586-7. [Google Scholar]
- Selosse, M.-A.; Cameron, D.D. Introduction to a Virtual Special Issue on Mycoheterotrophy: New Phytologist Sheds Light on Non-Green Plants. New Phytol. 2010, 185, 591–593. [Google Scholar] [CrossRef]
- Rasmussen, H.N.; Whigham, D.F. Phenology of Roots and Mycorrhiza in Orchid Species Differing in Phototrophic Strategy. New Phytol. 2002, 154, 797–807. [Google Scholar] [CrossRef]
- Selosse, M.-A. La Néottie, une “mangeuse” d’arbres. L′Orchidophile 2003, 155, 21–31. [Google Scholar]
- Tranchida-Lombardo, V.; Cafasso, D.; Cristaudo, A.; Cozzolino, S. Phylogeographic Patterns, Genetic Affinities and Morphological Differentiation between Epipactis helleborine and Related Lineages in a Mediterranean Glacial Refugium. Ann. Bot. 2011, 107, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Richard, A.J. Plant Breeding Systems, 1st ed.; George Allen & Unwin; Chapman and Hall: London, UK, 1986; Volume 3, ISBN 0-04-581020-6. [Google Scholar]
- Robatsch, K. Beiträge zur Kenntnis der europäischen Epipactis-Arten (Orchidaceae). Linz. Biol. Beiträge 1990, 20, 161–172. [Google Scholar]
- Robatsch, K. Beitrage zur Kenntnis des europe.aischen Epipactis-Arten (Orchidaceae) und zur Evolution der Autogamie bei europaischen und asiatischen Gattungen der Neottiodeae. J. Eur. Orch. 1995, 27, 125–177. [Google Scholar]
- Ehlers, B.K.; Pedersen, H.Æ. Genetic Variation in Three Species of Epipactis (Orchidaceae): Geographic Scale and Evolutionary Inferences. Biol. J. Linn. Soc. 2000, 69, 411–430. [Google Scholar] [CrossRef]
- Squirrell, J.; Hollingsworth, P.M.; Bateman, R.M.; Tebbitt, M.C.; Hollingsworth, M.L. Taxonomic Complexity and Breeding System Transitions: Conservation Genetics of the Epipactis leptochila Complex (Orchidaceae). Mol. Ecol. 2002, 11, 1957–1964. [Google Scholar] [CrossRef]
- Hollingsworth, P.; Squirrell, J.; Hollingsworth, M.; Richards, A.; Bateman, R. Taxonomic Complexity, Conservation and Recurrent Origins of Selfpollination in Epipactis (Orchidaceae). In Current Taxonomic Research on the British & European Flora; Bailey, J., Ellis, R.G., Eds.; BSBI: London, UK, 2006; pp. 27–44. [Google Scholar]
- Anghelescu, N.E.; Kertész, H.; Constantin, N.; Simon-Gruița, A.; Cornescu, G.D.; Pojoga, M.D.; Georgescu, M.I.; Petra, S.A.; Toma, F. New Intergeneric Orchid Hybrid Found in Romania × Pseudorhiza nieschalkii (Senghas) P.F.Hunt nothosubsp. siculorum H.Kertész & N.Anghelescu, 2020. PLoS ONE 2021, 16, e0241733. [Google Scholar] [CrossRef]
- Anghelescu, N.E.; Kertész, H.; Balogh, L.; Balogh, M.; Pataki, H.; Georgescu, M.I.; Petra, S.A.; Toma, F. First Description of Cypripedium calceolus and Cypripedium calceolus var. citrina B. Hergt: Morphology, Pollination and Conservation Status in Romania. AgroLife Sci. J. 2021, 10, 14–28. [Google Scholar] [CrossRef]
- Claessens, J.; Kleynen, J. Many Ways to Get Happy: Pollination Modes of European Epipactis Species. Orchid Dig. 2016, 80, 142–150. [Google Scholar]
- Tałałaj, I.; Brzosko, E. Selfing Potential in Epipactis palustris, E. helleborine and E. atrorubens (Orchidaceae). Plant Syst. Evol. 2008, 276, 21–29. [Google Scholar] [CrossRef]
- Dafni, A. Pollination Ecology: A Practical Approach, 1st ed.; Oxford University Press: New York, NY, USA, 1993; Volume 1, ISBN 978-0-19-963298-5. [Google Scholar]
- Jakubska-Busse, A.; Czeluśniak, I.; Kobyłka, M.J.; Hojniak, M. Why Does an Obligate Autogamous Orchid Produce Insect Attractants in Nectar?—A Case Study on Epipactis albensis (Orchidaceae). BMC Plant Biol. 2022, 22, 196. [Google Scholar] [CrossRef] [PubMed]
- Anghelescu, N.E.; Kertész, H.; Pataki, H.; Georgescu, M.I.; Petra, S.A.; Toma, F. Genus Ophrys L., 1753 in Romania—Taxonomy, Morphology and Pollination by Sexual Deception (Mimicry). Sci. Pap. Ser. B Hortic. 2021, LXV, 187–201. [Google Scholar] [CrossRef]
- Wucherpfennig, W. Epipactis albensis Nováková & Rydlo in Brandenburg. Ber. Arbeitskrs. Heim. Orchid. 1993, 10, 36–40. [Google Scholar]
- Sramkó, G.; Paun, O.; Brandrud, M.K.; Laczkó, L.; Molnár, A.V.; Bateman, R.M. Iterative Allogamy–Autogamy Transitions Drive Actual and Incipient Speciation during the Ongoing Evolutionary Radiation within the Orchid Genus Epipactis (Orchidaceae). Ann. Bot. 2019, 124, 481–497. [Google Scholar] [CrossRef] [PubMed]
- Brzosko, E.; Mirski, P. Floral Nectar Chemistry in Orchids: A Short Review and Meta-Analysis. Plants 2021, 10, 2315. [Google Scholar] [CrossRef]
- Pyke, G.H. What Does It Cost a Plant to Produce Floral Nectar? Nature 1991, 350, 58–59. [Google Scholar] [CrossRef]
- Claessens, J.; Kleynen, J. The Flower of the European Orchid. Form and Function, 1st ed.; Schrijen-Lippertz druk, print en multimedia Voerendaal: Stein, The Netherlands, 2011; ISBN 978-90-90-25556-9. [Google Scholar]
- Baumann, H.; Künkele, S.; Lorenz, R. Orchideen Europas. Mit angrenzenden Gebieten (Ulmers Naturführer), 1st ed.; Verlag Eugen Ulmer: Stuttgart, Germany, 2006; ISBN 978-3-8001-4162-3. [Google Scholar]
- Johnson, S.D.; Edwards, T.J. The Structure and Function of Orchid Pollinaria. Plant Syst. Evol. 2000, 222, 243–269. [Google Scholar] [CrossRef]
- Richards, A.J.; Porter, A.F. On the Identity of a Northumberland Epipactis. Watsonia 1982, 14, 121–128. [Google Scholar]
- Dafni, A.; Firmage, D. Pollen Viability and Longevity: Practical, Ecological and Evolutionary Implications. Plant Syst. Evol. 2000, 222, 113–132. [Google Scholar] [CrossRef]
- Kowalkowska, A.K.; Kostelecka, J.; Bohdanowicz, J.; Kapusta, M.; Rojek, J. Studies on Floral Nectary, Tepals’ Structure, and Gynostemium Morphology of Epipactis palustris (L.) Crantz (Orchidaceae). Protoplasma 2015, 252, 321–333. [Google Scholar] [CrossRef]
- Uphof, J.C.T. Cleistogamic Flowers. Bot. Rev. 1938, 4, 21–49. [Google Scholar] [CrossRef]
- Bonatti, P.M.; Sgarbi, E.; Del Prete, C. Gynostemium Micromorphology and Pollination in Epipactis microphylla (Orchidaceae). J. Plant Res. 2006, 119, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, H.Æ.; Ehlers, B.K. Local Evolution of Obligate Autogamy in Epipactis helleborine subsp. neerlandica (Orchidaceae). Plant Syst. Evol. 2000, 223, 173–183. [Google Scholar] [CrossRef]
- Catling, P.M. Auto-Pollination in the Orchidaceae (1990). In Orchid Biology: Reviews and Perspectives, X.; Kull, T., Arditti, J., Wong, S.M., Eds.; Springer Netherlands: Dordrecht, the Netherlands, 2010; Volume X, pp. 123–158. ISBN 978-1-4020-8801-8. [Google Scholar]
- Claessens, J.; Kleynen, J.; Reinhardt, J. Some Notes on the Genus Epipactis. Eurorchis 2000, 12, 75–83. [Google Scholar]
- Hagerup, O. Bud Antogamy in Some Northern Orchids. Phytomorphology Int. J. Plant Morphol. 1952, 2, 51–60. [Google Scholar]
- Robatsch, K. Beitrage zur Blütenbiologie und Autogamie der Gattung Epipactis. In Jahresberichte des Naturwissenschaftlichen Vereins in Wuppertal—Probleme der Taxonomie, Verbreitung und Vermehrung europäischer und mediterraner Orchideen; Senghas, K., Sundermann, H., Eds.; Satz und Druck: Hagemann-Druck, Hildesheim; Jahresber. Naturwiss. Ver. Wuppertal: Wuppertal, Germany, 1983; Volume 36, pp. 25–32. [Google Scholar]
- Schick, B. Zur Anatomie Und Biotechnik Des Bestaubungapparates Der Orchideen. II: Epipactis palustris (L.) Crantz Und Listera ovata (L.). R. Br. Bot. Jb. 1989, 110, 289–323. [Google Scholar]
- Yam, T.W.; Arditti, J.; Anghelescu, N.E. Predators in Orchids. Orchid. AOS—Am. Orchid Soc. 2022, 91, 920–933. [Google Scholar]
- Claessens, J.; Kleynen, J.; Wiellnga, R. Some Notes on Epipactis helleborine (L.) Crantz ssp. neerlandica (Vermeulen) Buttler and Epipactis renzii K. Robatsch. Eurorchis 1998, 10, 55–64. [Google Scholar]
- Claessens, J.; Kleynen, J. Allogamie-und Autogamie-Tendenzen bei einigen Vertretern der Gattung Epipactis. Ber. Arbeitskrs. Heim. Orchid. 1995, 12, 4–16. [Google Scholar]
- Claessens, J.; Kleynen, J. Quatre Epipactis de la flore de France 2me partie. L′Orchidophile 1997, 129, 225–229. [Google Scholar]
- Claessens, J.; Kleynen, J. Quatre Epipactis de la flore de France, 1e partie. L′Orchidophile 1997, 128, 167–172. [Google Scholar]
- Pridgeon, A.M.; Light, M.H.S. Epipactis. Pollination. In Genera Orchidacearum Volume 4: Epidendroideae (Part 1); Pridgeon, A.M., Cribb, P., Chase, M.W., Rasmussen, F.N., Eds.; Oxford University Press: Oxford, UK, 2005; Volume 4, p. 505. ISBN 978-0-19-850712-3. [Google Scholar]
- Stace, C.A. New Flora of the British Isles, 3rd ed.; Cambridge University Press: Cambridge, UK, 2010; ISBN 978-0-521-70772-5. [Google Scholar]
- Bateman, R. Evolution and Classification of European Orchids: Insights from Molecular and Morphological Characters. J. Eur. Orch. 2001, 33, 33–119. [Google Scholar]
- Bateman, R. Circumscribing Species in the European Orchid Flora: Multiple Datasets Interpreted in the Context of Speciation Mechanisms. Ber. Arbeitskrs. Heim. Orchid. 2012, 8, 160–212. [Google Scholar]
- Bateman, R. Circumscribing Genera in the European Orchid Flora: A Subjective Critique of Recent Contributions. Ber. Arbeitskrs. Heim. Orchid. 2012, 8, 92–124. [Google Scholar]
- Bateman, R. Evolutionary Classification of European Orchids: The Crucial Importance of Maximising Explicit Evidence and Minimising Authoritarian Speculation. J. Eur. Orch. 2009, 41, 243–318. [Google Scholar]
- Proctor, M.; Yeo, P. The Pollination of Flowers; Taplinger Publishing company: New York, NY, USA, 1973; ISBN 0-00-213178-1. [Google Scholar]
- Masel, J. Genetic Drift. Curr. Biol. 2011, 21, R837–R838. [Google Scholar] [CrossRef]
- Gould, S.J. Chapter 7, Section Synthesis as Hardening. In The Structure of Evolutionary Theory; Harvard University Press: Harvard, MA, USA, 2002; p. 1459. ISBN 978-0-674-41793-9. [Google Scholar]
- Jacquemyn, H.; Kort, H.D.; Broeck, A.V.; Brys, R. Immigrant and Extrinsic Hybrid Seed Inviability Contribute to Reproductive Isolation between Forest and Dune Ecotypes of Epipactis helleborine (Orchidaceae). Oikos 2018, 127, 73–84. [Google Scholar] [CrossRef]
- Reineke, D. Epipactis helleborine—ein Beispiel für die Unzulänglichkeit der heutigen Evolutionstheorien? Mitt. BI. Arb. Heim. Orchid. Baden-Württ. 1987, 19, 822–834. [Google Scholar]
- Franz, G. Epipactis helleborine lus. fIavescens auf einer Schwermetall-halde in Stolberg (Rheinland). Ber. Arbeitskrs. Heim. Orchid. 1995, 12, 102–105. [Google Scholar]
- Wucherpfennig, W. Beobachtungen an vier wenig bekannten autogamen Epipactis-Arten Mitteleuropas. Ber. Arbeitskrs. Heim. Orchid. 1993, 10, 9–24. [Google Scholar]
- Wright, S.I.; Kalisz, S.; Slotte, T. Evolutionary Consequences of Self-Fertilization in Plants. Proc. R. Soc. B Biol. Sci. 2013, 280, 20130133. [Google Scholar] [CrossRef]
- Reinhard, H.R. Autogamie bei europäischen Orchideen. Orchids 1977, 28, 178–182. [Google Scholar]
- Suetsugu, K. Autonomous Self-Pollination and Insect Visitors in Partially and Fully Mycoheterotrophic Species of Cymbidium (Orchidaceae). J. Plant Res. 2015, 128, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Stebbins, G.L. Self Fertilization and Population Variability in the Higher Plants. Am. Nat. 1957, 91, 337–354. [Google Scholar] [CrossRef]
- Igic, B.; Busch, J.W. Is Self-Fertilization an Evolutionary Dead End? New Phytol. 2013, 198, 386–397. [Google Scholar] [CrossRef] [PubMed]
- Takebayashi, N.; Morrell, P.L. Is Self-Fertilization an Evolutionary Dead End? Revisiting an Old Hypothesis with Genetic Theories and a Macroevolutionary Approach. Am. J. Bot. 2001, 88, 1143–1150. [Google Scholar] [CrossRef]
- Woodruff, D.S. Populations, Species, and Conservation Genetics. Encycl. Biodivers. 2001, 811–829. [Google Scholar] [CrossRef]
- Darwin, C. The Effects of Cross and Self Fertilisation in the Vegetable Kingdom; First published in: 1876, London: Murray; Cambridge Library Collection—Darwin, Evolution and Genetics; Cambridge University Press: Cambridge, UK, 2009; ISBN 978-0-511-69420-2. [Google Scholar]
- Darwin, C. The Different Forms of Flowers on Plants of the Same Species; First published in: 1877, London: Murray; Cambridge Library Collection—Darwin, Evolution and Genetics; Cambridge University Press: Cambridge, UK, 2010; ISBN 978-0-511-73141-9. [Google Scholar]
- Eckert, C.G. Contributions of Autogamy and Geitonogamy to Self-Fertilization in a Mass-Flowering, Clonal Plant. Ecology 2000, 81, 532–542. [Google Scholar] [CrossRef]
- Wyatt, R. The Evolution of Self-Pollination in Granite Outcrop Species of Arenaria (Caryophyllaceae) V. Artificial Crosses Within and Between Populations. Syst. Bot. 1990, 15, 363–369. [Google Scholar] [CrossRef]
- Wyatt, R. The Evolution of Self-pollination in Granite Outcrop Species of Arenaria (Caryophyllaceae). I. Morphological Correlates. Evolution 1984, 38, 804–816. [Google Scholar] [CrossRef] [PubMed]
- Jakubska-Busse, A.; Janowicz, M.; Ochnio, L.; Jackowska-Zduniak, B. Shapes of Leaves with Parallel Venation. Modelling of the Epipactis sp. (Orchidaceae) Leaves with the Help of a System of Coupled Elastic Beams. PeerJ 2016, 4, e2165. [Google Scholar] [CrossRef] [PubMed]
- Bateman, R.M. Integrating Molecular and Morphological Evidence for Evolutionary Radiations. In Molecular Systematics and Plant Evolution; Hollingsworth, P.M., Bateman, R.M., Gornall, R.J., Eds.; Taylor & Franci: London, UK, 1999; pp. 432–471. ISBN 978-0-7484-0907-5. [Google Scholar]
- Hedrén, M.; Anghelescu, N.E.; Lorenz, R. Geographic Differentiation in Nigritella nigra s.l. from the Pyrenées, the Jura, the Eastern Alps and the Southern Carpathians. J. Eur. Orch. 2022, 54, 266–328. [Google Scholar]
- Georgescu, M.I.; Peticilă, A.G.; Dobrescu, E.; Costache, N. The Importance of Botanical Survey and Proper Management in the Protection of Rare, Endangered or Vulnerable Species Outside of Protected Areas—Working Example: Hyacintella leucophaea (K. Koch) Schur. Sci. Pap. Ser. B Hortic. 2022, LXVI, 685–691. [Google Scholar]
- Burghilă, C.; Bordun, C.; Cîmpeanu, S.M.; Burghilă, D.; Badea, A. Why Mapping Ecosystems Services Is a Must in EU Biodiversity Strategy for 2020? AgroLife Sci. J. 2016, 5, 28–37. [Google Scholar]
- The IUCN Red List of Threatened Species:Version 3.1. Available online: https://www.iucnredlist.org/en (accessed on 1 April 2023).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetative and Floral Organs | Characters/Features | Epipactis bucegensis | ||
|---|---|---|---|---|
| (Millimetres) | Epipactis muelleri | Epipactis helleborine | ||
| 1. Rhizome | ||||
| Rhizome length | 40–120 (160) | NA | 50–150 (180) | |
| Rhizome diam. | 13–25 (31) | NA | 13–26 (38) | |
| 2. Adventitious Roots | ||||
| Adv. roots length | 20 (34)–100 (124) | NA | 20 (34)–100 (124) | |
| Adv. roots diam. | 1.5–2.0 (3.2) | NA | 1.5–2.0 (3.2) | |
| 3. Stem | ||||
| * Stem type | * Spindly, flexuous, yellowish-green | Spindly, greenish | Strong, robust, green | |
| Overall height | 180–300 (500) | (10) 200–350 (400) | (200) 350–900 (1300) | |
| Stem diameter | 3.0–5.0 (8.0) | 3.0–4.3 (6.9) | 5.0–9.0 (14) | |
| Stem anthocyanins | Absent | Absent | Present basally | |
| * Trichomes (Glandular hairs) | * Densely glandular–pubescent along the entire stem | Basal glandular–pubescent | Basal glabrous, downy towards the tip | |
| 4. Leaves | ||||
| * Basal sheath | * 1, acuminate–lanceolate, tapering at tip, yellowish-green | 1, acuminate–elongate, light green | 1, round–ovoidal, wide around the middle, green | |
| Basal sheath length | 15–25 | 14 (16)–22 (25) | 18–31 | |
| Basal sheath width | 9.0–20 | 7.0 (9)–15 (20) | 10–25 | |
| * Distribution of sheathing leaves on stem | * Numerous in the lower half | Numerous in the middle part | Numerous in the middle part of the stem | |
| Phyllotaxy | Alternate | Alternate | Alternate | |
| * Longest leaf posture | * Spreading to erect, to a subtended angle of c. 30° relative to the stem (angled) | Spreading horizontally to curved downwards, arched | Spreading horizontally to a subtended angle of c. 90° relative to the stem (perpendicular to the stem) | |
| * No. of basal cauline leaves | * 3–4 (8), elongate, acuminate | 1 (3), ovoid–elongate, wide, arched, acuminate | 1 basal sheath (see above) | |
| No. of median cauline leaves | 3 (5), elongate–oval, acuminate | 5 (8), ovoid–elongate, wide, arched, acuminate | 4 (10), ovoid–orbicular | |
| No. of upper cauline leaves | 1–2 (4), narrow–lanceolate acuminate | 1–2, narrow–lanceolate acuminate | 3 (4), lanceolate acuminate | |
| Length of longest leaf | 90 (120) | 33–100 (125) | 100 (140) | |
| Width of longest leaf | 35 (40) | 1.9–3 (4.3) | 60 (75) | |
| Outline shape of longest leaf | Oval to elongate–lanceolate, broadest in the middle | Ovoid–elongate, acuminate, broadest in the middle | Orbicular to ovoid–elongated, broadest in the middle | |
| * Leaf conduplicate | * Strongly keeled | Keeled | Moderately keeled | |
| * Apical hooding | * Moderate | Absent | Absent | |
| * Leaf colour | * Yellowish-green to green (aged plants) | Light- to deep-green | Green to dark-green | |
| * Leaf dorsal side | * Yellowish | Light- to deep-green | Green to dark-green | |
| * Leaf ventral side | * Shiny, yellowish-green | Shiny, light-green | Shiny, deep-green | |
| Leaf margins | Undulate, edged with fine papillae | Undulate | Entire, straight, edged with fine papillae | |
| Leaf markings | Unspotted | Unspotted | Unspotted | |
| * Leaf venation | * Strongly veined | Veined | Veined | |
| Upmost leaf | Bract-like, narrow–acuminated, yellowish | Bract-like, narrow–acuminated, greenish | Bract-like, deep- to dark-green | |
| 5. Bracts | ||||
| Length of basal bracts | 15–38 (45) exceeding the flowers | 12–33 (45) exceeding the flowers | 35–58 (102), exceeding flowers | |
| Width of basal bracts | 3.2 (4.2)–8.0 (12) | 3.2–5.4 (8.3) | 19–(24) 32 | |
| Length of floral bracts | 18–22 longer than the ovaries, shorter than the flowers | 19–25 longer than the ovaries, shorter than the flowers | 28–41 exceeding flowers | |
| Width of floral bracts | 2.0 (3.0)–4.0 (8.0) | 2.0 (2.8)–3.6 (6.5) | 12 (23)–14 (28) | |
| Texture of bracts | Robust, keeled | Membranous | Robust to membranous | |
| Bract anthocyanins | Absent, bracts yellowish | Absent, bracts light green | Absent, bracts deep-green | |
| Bract margins | Entire, undulation missing, edged with tooth-like papillae | Entire, undulation missing, edged | Entire, undulation missing, edged with tooth-like papillae | |
| 6. Inflorescence | Inflorescence length | 100–150 | 100 (200)–250 (300) | 100–200 (450) |
| Inflorescence diameter (calculated from bract-tip to bract-tip) | 35–45 (55) | 25–35 (45) | 35–55 (95) | |
| Inflorescence shape | Elongated raceme, near-one-sided, lax to dense | Elongated raceme, lax, near-one-sided | Elongated raceme, near-one-sided, dense | |
| No. of flowers | 10–30 (50/65) | 4 (10)–15 (35) | 10–30 (70/100) | |
| 7. Flowers | Flower type | Cleistogamous, half- to completely closed, pendant | Cleistogamous, half-opened, pendant | Chasmogamous, wide-opened, near-erect |
| Flower length | 8.5–11.2 | 7.2–9.0 | 8.8–14.3 | |
| Flower diameter | 4.5–6.5 | 3.3–5.5 | 4.8–7.8 | |
| Flower colour | Whitish-yellow to yellowish-green (aged individuals) | Yellowish-green | Greenish, purple, violet, pink, brownish | |
| 8. Scent | Absent | Absent | Faint to moderate, sweet to fermented | |
| 9. Sepals | ||||
| Sepal shape | Ovoid–elongated | Ovoid–elongated | Ovoid–roundish | |
| Sepal no. and colour | 3, Whitish-yellowish | 3, Yellowish-green | 3, Greenish, violet-purple tinged | |
| Sepal apex | Acuminate, tapering | Acuminate, tapering | Roundish–acuminate | |
| Sepal length | 7.0–14.2 | 5.0 (6.5)–10.2 (11) | 7.2–14.2 | |
| Sepal width | 5.5–7.1 | 3.9–4.5 | 4.5–7.5 (8.8) | |
| 10. Lateral petals | Petal no. and colour | 2, Whitish-yellowish | 3, Yellowish-green to pinkish-green | 2, Greenish, violet-purple, brownish-purple tinged |
| Lateral petal shape | Oval–lanceolate, tapering | Oval–lanceolate | Roundish–oval lanceolate | |
| Lateral petal apex | Acuminate elongated, tapering | Acuminate elongated, tapering | Acuminate elongated, tapering | |
| Lateral petal length | 6.0–9.0 | 4.9–6.2 (7.1) | 6.9–12.5 (13.5) | |
| Lateral petal width | 4.0–6.1 | 3.0–4.1 (5.50) | 4.0–6.1 (6.9) | |
| 11. * Labellum | ||||
| Outline shape | Orbicular, wide, flat, bipartite | Orbicular, tripartite | Orbicular, tripartite | |
| * Hypochile shape | * Wide, ovoid | Orbicular, cup-like | Orbicular, cup-like | |
| Hypochile length | 3.0–4.1 | 3.2–3.8 (4.2) | 2.1 (3.0)–3.9 | |
| Hypochile width | 4.0–5.2 | 3.6–4.5 (5.1) | 3.8–4.2 | |
| * Hypochile inner wall colour | * Shiny, crimson-purple to brownish-purple | Shiny, blackish-brown | Shiny, blackish-brown to blackish-green | |
| * Mesochile | * Completely absent | Well-defined, wide junction | Well-defined, narrow junction | |
| * Epichile shape | * Triangular, flat, tapering | Obtuse, heart-shaped, roundish–wide at the apex | Heart-shaped, obtuse, turned downwards | |
| Epichile length | 5.0–5.8 | 4.5–5.5 (6.1) | 5.8.0–6.7 (7.1) | |
| Epichile width | 3.0–4.5 | 3.9–4.0 | 3.1–4.2 (5.9) | |
| * Epichile colour | * White | Yellowish-green, purple tinted | Greenish-purple, brownish | |
| * Epichile lateral lobes | * Absent to mildly defined | Defined, ovoid-roundish, scalloped | Defined, ovoid–roundish | |
| * Median lobe | * Triangular, flat, tapering | Triangular, roundish, obtuse | Roundish, obtuse, downcurved | |
| 12. * Calli | * Basal calli no. and colour | * 2, pyramidal, tooth-like, prominent, wide apart, crimson-purple, non-wrinkled | 2, less-prominent, wrinkled, attenuated, greenish-yellow to pinkish, closely placed | 2, prominent, wrinkled, well-developed, variously coloured |
| Central groove | Present, well defined with a minute basal callus | Present, greenish | Present, well-defined, darker | |
| * Central lobe apex | * Narrow, acuminate/tapering, elongated | Roundish, wide, obtuse | Roundish, wide, obtuse | |
| * Base colour of median lobe | * White | Greenish-yellow | Variously coloured—green, violet, brownish | |
| * Margin colour | * White | Greenish-yellow | Variously coloured | |
| Lateral lobes | Scalloped, poorly defined to completely absent | Scalloped, moderate indentation | Entire/moderate indentation, scalloped, slightly recurved | |
| Lateral lobe margins | White, entire, scalloped | Greenish, entire, scalloped | White, entire to moderate indentation | |
| 13. Spur | ||||
| Presence | Absent | Absent | Absent | |
| 14. Nectar | ||||
| Presence | Absent-to-minute droplets in topmost flowers | Present in fair amounts | Present in abundant amounts | |
| 15. Gynostemium | ||||
| Type | Thick, cylindrical | Thick | Thick | |
| Colour | White, shiny | Creamy-white | Creamy-white | |
| 16. Staminodes | ||||
| Presence | Present, wing-like | Well-defined, wing-like, large | Well-defined, wing-like, large | |
| 17. Auricles | ||||
| Presence | Absent | Absent | Absent | |
| 18. Clinandrium | ||||
| Presence | Absent | Absent | Present, well-defined | |
| 19. Anther | ||||
| Anther colour | Translucent white | Creamy-white | Creamy-white to white | |
| Anther shape | Broad, short | Broad, short | Broad, short | |
| Type | Sessile, broad, bithecal | Sessile, broad, bithecal | Stalked, bithecal | |
| Angle relative to stigma | Angled, pushed forward | Angled, pushed forward | Erect | |
| 20. * Anther cap | ||||
| * Anther cap shape | * Elongated, fleshy, tapering | Elongated, fleshy, acuminate | Elongated, fleshy, tapering to roundish | |
| * Anther cap colour | * Whitish-creamy | Yellowish | ||
| Type | Well-developed, bithecal | Well-developed, bithecal | Well-developed, bithecal | |
| 21. Pollinia | ||||
| Shape | Elongated, ovoid, bipartite | Elongated, ovoid, bipartite | Elongated, clavate, bipartite | |
| Pollinia length | 1.5–2.3 | 1.6–2.5 | 1.8–2.9 (3.1) | |
| Pollinia width | 0.5–1.2 | 0.6–1.4 | 0.5–1.4 (1.8) | |
| Compactness | Mealy, friable, disintegrating | Moderately compact to friable | Compact, non-disintegrating | |
| Contact with stigma | Stigmatic contact before anthesis | Stigmatic contact before anthesis | No stigmatic contact | |
| 22. Viscidium | ||||
| Presence | Absent | Absent | Present, well-developed, sac-like, milky | |
| 23. * Stigma | ||||
| * Position relative to the gynostemium | * Perpendicular, roof-like, entirely flat | Perpendicular, deeply V-shaped, deeply concave | Parallel to near-parallel, slightly concave | |
| *Shape | * Rectangular, wider than longer, bilobed (rostellum absent), roof-like, entirely flat | Quadrangular, bilobed (rostellum absent), deeply V-shaped, deeply concave | Quadrangular, trilobed | |
| Stigmatic exudate | Highly abundant | Abundant | Fair amounts | |
| 24. Rostellum | ||||
| Presence | Absent | Absent | Present, well-developed (median stigmatic lobe) forming the upper stigmatic rim | |
| 25. Bursicles | ||||
| Presence | Absent | Absent | Absent | |
| 26. Ovary | ||||
| Ovary shape and colour | Pear-shaped, round apex, strongly ridged | Elongated, spindle-shaped, light-green, non-ridged | Pear-shaped, mildly ridged | |
| Ovary colour | Dark green, mildly purple pigmented (anthocyanins) | Light-green | Green to dark green | |
| Ovary length | 6.2–8.5 | 6.2 (7.3)–8.8 (9.0) | 5.2–9.5 | |
| Ovary diameter | 3.9–4.8 | 3.2–4.0 (4.5) | 3.7–5.9 | |
| * Ovary colour | * Yellowish-green to deep-green in fruiting plants | Green | Green to dark-green | |
| 27. Flower pedicel | ||||
| Pedicel length | 1.2–2.5 | 1.6–2.9 | ||
| Pedicel diam. | 1.0–1.2 | 1.0–1.4 | ||
| * Pedicel pigmentation | * Faint violet-washed at base (entirely greenish) | entirely greenish, with no basal pigmentation | Purple-pigmented at base | |
| * Pedicel colour | * Yellowish-green | Greenish | Greenish-violet basally | |
| Pedicel pubescence | Downy, covered in glandular hairs (trichomes) | Absent to mildly pubescent | Downy, covered in glandular hairs (trichomes) | |
| 28. * Fruit | ||||
| * Fruit shape | Club/*ear-shaped, swollen, ovoid with round apex, strongly ridged | Pear-shaped, elongated-ovoid, smooth | Pear-shaped, swollen middle part, mildly ridged | |
| * Fruit colour | Green to dark green (aged plants), purple washed at the base | Light-green | Green to dark green (aged plants) | |
| Fruit length | 6.9–10.5 (12.3) | 6.9–10.5 (12.3) | 5.9–10.4 (15.3) | |
| Fruit width | 4.9–5.9 (6.3) | 4.9–5.9 (6.3) | 4.7–6.1 (6.8) | |
| Fruiting | July–August | June–July | July–August | |
| Fruit set | 70–95% | 10–30% (60%) | 60–78% | |
| 29. Seed capsule | ||||
| Capsule length | 7.1–12 (13.1) | 6.8–9 (10.5) | 6.2–12.5 (14.1) | |
| Capsule width | 5.1–6.5 | 4.8–5 (6.2) | 5.5–7.2 | |
| Capsule colour | Brownish | Brownish | Brownish | |
| Capsule maturation | September–October | September–October | September–October | |
| 30. Flowering time | July | June | July–August | |
| 31. Pollination strategy | ||||
| Pollination type | Non-entomophilous | Non-entomophilous | Entomophilous | |
| Pollination mode | Self-pollinating | Self-pollinating | Cross-pollinating | |
| 32. Reproductive strategy | ||||
| * Reproductive strategy type | * Obligate autogamous | Obligate/Facultative autogamous | Obligate allogamous |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anghelescu, N.E.; Balogh, M.; Balogh, L.; Kigyossy, N.; Georgescu, M.I.; Petra, S.A.; Toma, F.; Peticilă, A.G. Epipactis bucegensis—A Separate Autogamous Species within the E. helleborine Alliance. Plants 2023, 12, 1761. https://doi.org/10.3390/plants12091761
Anghelescu NE, Balogh M, Balogh L, Kigyossy N, Georgescu MI, Petra SA, Toma F, Peticilă AG. Epipactis bucegensis—A Separate Autogamous Species within the E. helleborine Alliance. Plants. 2023; 12(9):1761. https://doi.org/10.3390/plants12091761
Chicago/Turabian StyleAnghelescu, Nora E., Mihaela Balogh, Lori Balogh, Nicoleta Kigyossy, Mihaela I. Georgescu, Sorina A. Petra, Florin Toma, and Adrian G. Peticilă. 2023. "Epipactis bucegensis—A Separate Autogamous Species within the E. helleborine Alliance" Plants 12, no. 9: 1761. https://doi.org/10.3390/plants12091761
APA StyleAnghelescu, N. E., Balogh, M., Balogh, L., Kigyossy, N., Georgescu, M. I., Petra, S. A., Toma, F., & Peticilă, A. G. (2023). Epipactis bucegensis—A Separate Autogamous Species within the E. helleborine Alliance. Plants, 12(9), 1761. https://doi.org/10.3390/plants12091761