
Detection, Diagnosis, and Preventive Management of the Bacterial Plant Pathogen Pseudomonas syringae





Abstract
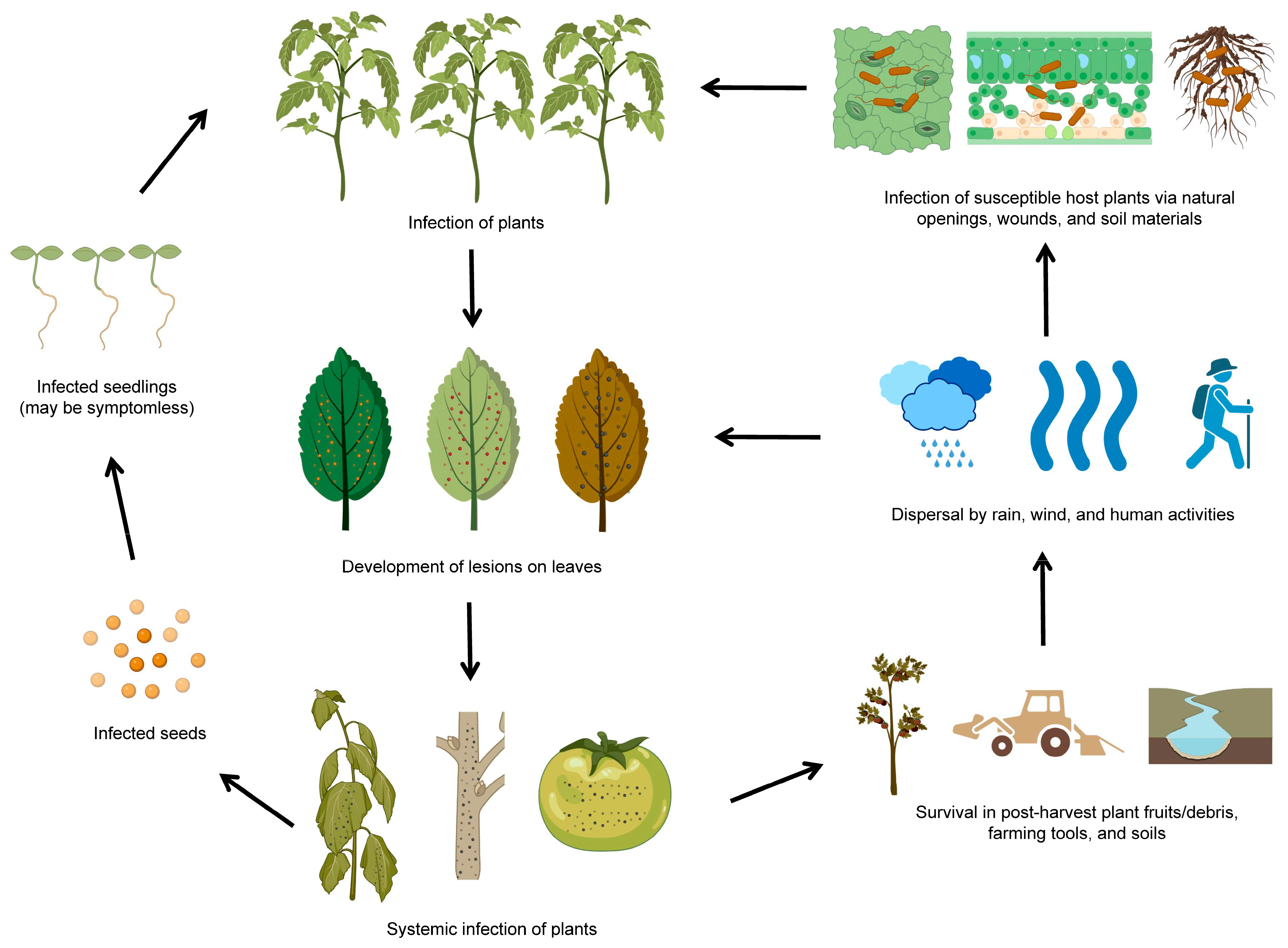
:1. Introduction
2. P. syringae as a Bacterial Plant Pathogen
3. Detection and Diagnosis of P. syringae with Conventional Methods
4. Detection and Diagnosis of P. syringae by Molecular and Genetic Methods
5. Detection and Diagnosis of P. syringae with Serological Methods
6. Detection and Diagnosis of P. syringae with Biomarker-Based Methods
7. Detection and Diagnosis of P. syringae with Vision-Based Methods
8. Detection and Diagnosis of P. syringae by AI (Artificial Intelligence) Methods
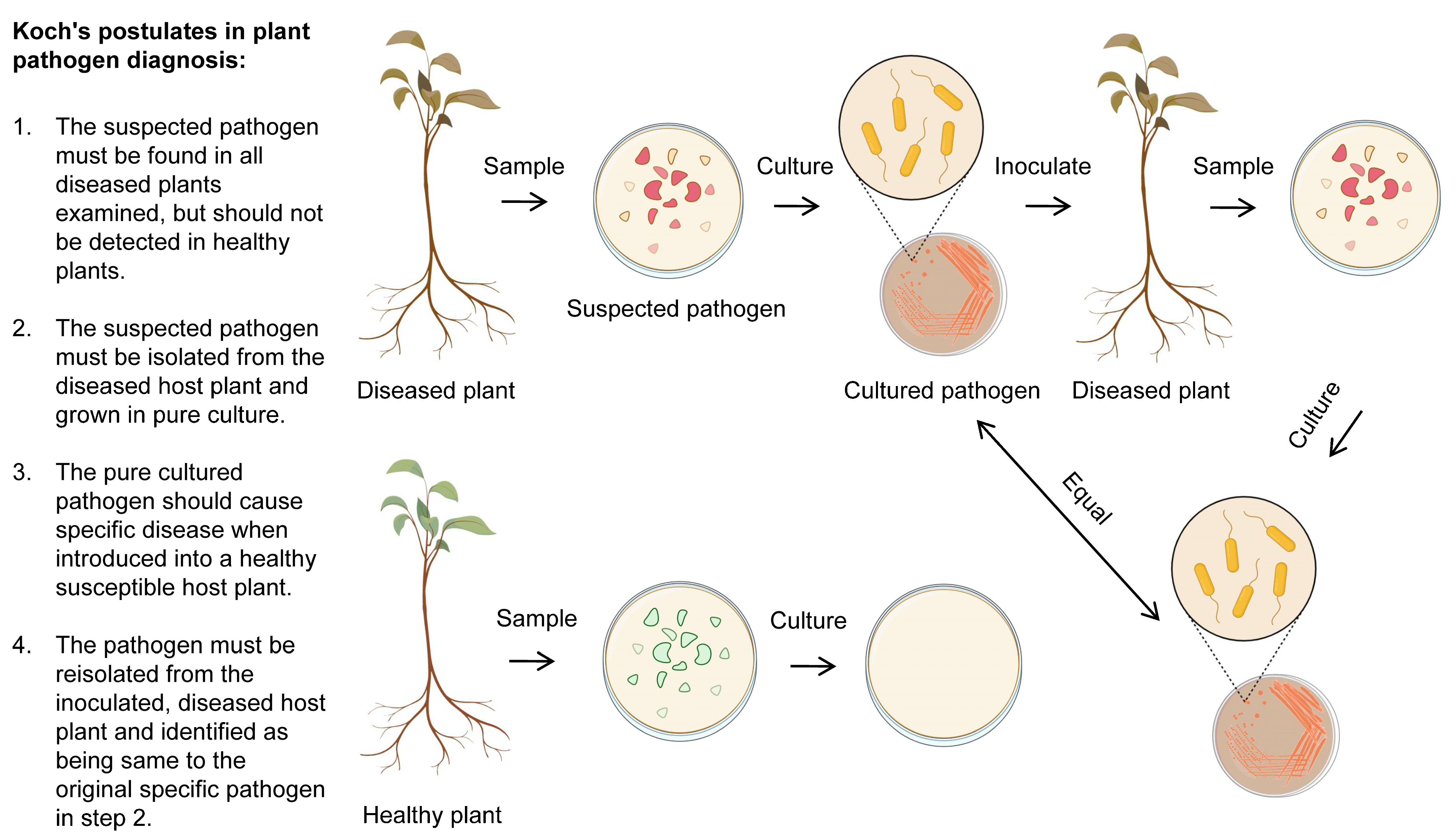
9. Koch’s Postulates as Golden Rules in the Detection and Diagnosis of P. syringae
10. Conclusions
11. Future Directions on Preventive Management of P. syringae
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mohanty, S.P.; Hughes, D.P.; Salathé, M. Using Deep Learning for Image-Based Plant Disease Detection. Front. Plant Sci. 2016, 7, 1419. [Google Scholar] [CrossRef]
- Liu, J.; Wang, X. Plant Diseases and Pests Detection Based on Deep Learning: A Review. Plant Methods 2021, 17, 22. [Google Scholar] [CrossRef] [PubMed]
- Harakannanavar, S.S.; Rudagi, J.M.; Puranikmath, V.I.; Siddiqua, A.; Pramodhini, R. Plant Leaf Disease Detection Using Computer Vision and Machine Learning Algorithms. Glob. Transit. Proc. 2022, 3, 305–310. [Google Scholar] [CrossRef]
- Plant Disease Diagnosis. Available online: https://www.apsnet.org/edcenter/disimpactmngmnt/casestudies/Pages/PlantDiseaseDiagnosis.aspx (accessed on 3 February 2023).
- Meena, A.K.; Godara, S.L.; Meena, P.N. Detection and Diagnosis of Plant Diseases; Scientific Publishers: Jodhpur, India, 2020; ISBN 978-93-89184-42-6. [Google Scholar]
- Gullino, M.L.; Bonants, P.J.M. (Eds.) Detection and Diagnostics of Plant Pathogens; Springer: Dordrecht, The Netherlands, 2014; ISBN 978-94-017-9019-2. [Google Scholar]
- FAO. The State of Food and Agriculture 2021: Making Agrifood Systems More Resilient to Shocks and Stresses; The State of Food and Agriculture (SOFA); FAO: Rome, Italy, 2021; ISBN 978-92-5-134329-6. [Google Scholar]
- Yang, P.; Chen, Y.; Wu, H.; Fang, W.; Liang, Q.; Zheng, Y.; Olsson, S.; Zhang, D.; Zhou, J.; Wang, Z.; et al. The 5-Oxoprolinase Is Required for Conidiation, Sexual Reproduction, Virulence and Deoxynivalenol Production of Fusarium graminearum. Curr. Genet. 2018, 64, 285–301. [Google Scholar] [CrossRef]
- Yang, P.; Zhao, Z.; Fan, J.; Liang, Y.; Bernier, M.C.; Gao, Y.; Zhao, L.; Opiyo, S.O.; Xia, Y. Bacillus proteolyticus OSUB18 Triggers Induced Systemic Resistance against Bacterial and Fungal Pathogens in Arabidopsis. Front. Plant Sci. 2023, 14, 1078100. [Google Scholar] [CrossRef]
- Zhao, L.; Yang, P.; Li, W.; Zhao, Z.; Xia, Y. First Report of Trichoderma crassum Causing Leaf Spot on Tomato (Solanum lycopersicum Cv. M82) in Ohio. Plant Dis. 2023, 107, 582. [Google Scholar] [CrossRef] [PubMed]
- Cavalazzi, B.; Lemelle, L.; Simionovici, A.; Cady, S.L.; Russell, M.J.; Bailo, E.; Canteri, R.; Enrico, E.; Manceau, A.; Maris, A.; et al. Cellular Remains in a ~3.42-Billion-Year-Old Subseafloor Hydrothermal Environment. Sci. Adv. 2021, 7, eabf3963. [Google Scholar] [CrossRef]
- Ehrlich, P. Address in Pathology, on Chemiotherapy. Br. Med. J. 1913, 2, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Yang, X.; Lü, S.; Fan, J.; Opiyo, S.; Yang, P.; Mangold, J.; Mackey, D.; Xia, Y. Deciphering the Novel Role of AtMIN7 in Cuticle Formation and Defense against the Bacterial Pathogen Infection. Int. J. Mol. Sci. 2020, 21, 5547. [Google Scholar] [CrossRef]
- Zhao, Z.; Fan, J.; Gao, Y.G.; Wang, Z.; Yang, P.; Liang, Y.; Opiyo, S.; Xia, Y. Arabidopsis Plasma Membrane ATPase AHA5 Is Negatively Involved in PAMP-Triggered Immunity. Int. J. Mol. Sci. 2022, 23, 3857. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Fan, J.; Yang, P.; Wang, Z.; Stephen, O.; Mackey, D.; Ye, X. Involvement of Arabidopsis Acyl Carrier Protein 1 in PAMP-Triggered Immunity. Mol. Plant-Microbe Interact. 2022, 35, 681–693. [Google Scholar] [CrossRef]
- Plant Disease Management Strategies. Available online: https://www.apsnet.org/edcenter/disimpactmngmnt/topc/EpidemiologyTemporal/Pages/ManagementStrategies.aspx (accessed on 3 February 2023).
- Buttimer, C.; McAuliffe, O.; Ross, R.P.; Hill, C.; O’Mahony, J.; Coffey, A. Bacteriophages and Bacterial Plant Diseases. Front. Microbiol. 2017, 8, 34. [Google Scholar] [CrossRef]
- Rapid and Accurate Disease Diagnosis as a Key Component to Successful Plant Disease Management. Available online: https://extension.unh.edu/blog/2022/02/rapid-accurate-disease-diagnosis-key-component-successful-plant-disease-management (accessed on 3 February 2023).
- Rizzo, D.M.; Lichtveld, M.; Mazet, J.A.K.; Togami, E.; Miller, S.A. Plant Health and Its Effects on Food Safety and Security in a One Health Framework: Four Case Studies. One Health Outlook 2021, 3, 6. [Google Scholar] [CrossRef]
- Ristaino, J.B.; Anderson, P.K.; Bebber, D.P.; Brauman, K.A.; Cunniffe, N.J.; Fedoroff, N.V.; Finegold, C.; Garrett, K.A.; Gilligan, C.A.; Jones, C.M.; et al. The Persistent Threat of Emerging Plant Disease Pandemics to Global Food Security. Proc. Natl. Acad. Sci. USA 2021, 118, e2022239118. [Google Scholar] [CrossRef]
- Mooney, H.A.; Cleland, E.E. The Evolutionary Impact of Invasive Species. Proc. Natl. Acad. Sci. USA 2001, 98, 5446–5451. [Google Scholar] [CrossRef] [PubMed]
- López, M.M.; Llop, P.; Olmos, A.; Marco-Noales, E.; Cambra, M.; Bertolini, E. Are Molecular Tools Solving the Challenges Posed by Detection of Plant Pathogenic Bacteria and Viruses? Curr. Issues Mol. Biol. 2009, 11, 13–46. [Google Scholar] [PubMed]
- Khiyami, M.A.; Almoammar, H.; Awad, Y.M.; Alghuthaymi, M.A.; Abd-Elsalam, K.A. Plant Pathogen Nanodiagnostic Techniques: Forthcoming Changes? Biotechnol. Biotechnol. Equip. 2014, 28, 775–785. [Google Scholar] [CrossRef]
- Buja, I.; Sabella, E.; Monteduro, A.G.; Chiriacò, M.S.; De Bellis, L.; Luvisi, A.; Maruccio, G. Advances in Plant Disease Detection and Monitoring: From Traditional Assays to In-Field Diagnostics. Sensors 2021, 21, 2129. [Google Scholar] [CrossRef] [PubMed]
- Pfeilmeier, S.; Caly, D.L.; Malone, J.G. Bacterial Pathogenesis of Plants: Future Challenges from a Microbial Perspective. Mol. Plant Pathol. 2016, 17, 1298–1313. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, F.; Thilmony, R.; He, S.Y. The Arabidopsis thaliana-Pseudomonas syringae Interaction. In Arabidopsis Book; The American Society of Plant Biologists: Rockville, MD, USA, 2002; Volume 2002. [Google Scholar] [CrossRef]
- Xin, X.-F.; He, S.Y. Pseudomonas syringae pv. tomato Dc3000: A Model Pathogen for Probing Disease Susceptibility and Hormone Signaling in Plants. Annu. Rev. Phytopathol. 2013, 51, 473–498. [Google Scholar] [CrossRef] [PubMed]
- Gomila, M.; Busquets, A.; Mulet, M.; García-Valdés, E.; Lalucat, J. Clarification of Taxonomic Status within the Pseudomonas syringae Species Group Based on a Phylogenomic Analysis. Front. Microbiol. 2017, 8, 2422. [Google Scholar] [CrossRef] [PubMed]
- Hirano, S.S.; Upper, C.D. Bacteria in the Leaf Ecosystem with Emphasis On Pseudomonas syringae—A Pathogen, Ice Nucleus, and Epiphyte. Microbiol. Mol. Biol. Rev. 2000, 64, 624–653. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.-F.; Kvitko, B.; He, S.Y. Pseudomonas syringae: What It Takes to Be a Pathogen. Nat. Rev. Microbiol. 2018, 16, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Gardan, L.; Shafik, H.; Belouin, S.; Broch, R.; Grimont, F.; Grimont, P.A.D.Y. 1999 DNA Relatedness among the Pathovars of Pseudomonas syringae and Description of Pseudomonas tremae sp. nov. and Pseudomonas cannabina sp. nov. (Ex Sutic and Dowson 1959). Int. J. Syst. Evol. Microbiol. 1999, 49, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Preston, G.M. Pseudomonas syringae pv. tomato: The Right Pathogen, of the Right Plant, at the Right Time. Mol. Plant Pathol. 2000, 1, 263–275. [Google Scholar] [CrossRef]
- Morris, C.E.; Sands, D.C.; Vinatzer, B.A.; Glaux, C.; Guilbaud, C.; Buffière, A.; Yan, S.; Dominguez, H.; Thompson, B.M. The Life History of the Plant Pathogen Pseudomonas syringae Is Linked to the Water Cycle. ISME J. 2008, 2, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.B.; Zitter, T.A.; Momol, T.M.; Miller, S.A. Compendium of Tomato Diseases and Pests, 2nd ed.; Jeffrey, B.J., Thomas, A.Z., Timur, M.M., Sally, A.M., Eds.; Diseases and Pests Compendium Series; The American Phytopathological Society: Saint Paul, MN, USA, 2016; ISBN 978-0-89054-434-1. [Google Scholar]
- Takikawa, Y.; Tsuyumu, S.; Goto, M. Occurrence of Bacterial Leaf Spot of Maple Incited by Pseudomonas Syrinigae pv. aceris in Japan. Jpn. J. Phytopathol. 1991, 57, 724–728. [Google Scholar] [CrossRef]
- Takikawa, Y.; Serizawa, S.; Ichikawa, T.; Tsuyumu, S.; Goto, M. Pseudomonas syringae pv. actinidiae pv. nov.: The Causal Bacterium of Canker of Kiwifruit in Japan. Ann. Phytopathol. Soc. Jpn. 1989, 55, 437–444. [Google Scholar] [CrossRef]
- Durgapal, J.C.; Singh, B. Taxonomy of Pseudomonads Pathogenic to Horse-Chestnut, Wild Fig and Wild Cherry in India. Indian Phytopathol. 1980, 33, 533–535. [Google Scholar]
- Hendson, M.; Hildebrand, D.C.; Schroth, M.N. Relatedness of Pseudomonas syringae pv. tomato, Pseudomonas syringae pv. maculicola and Pseudomonas syringae pv. antirrhini. J. Appl. Bacteriol. 1992, 73, 455–464. [Google Scholar] [CrossRef]
- Gardan, L.; Cottin, S.; Bollet, C.; Hunault, G. Phenotypic Heterogeneity of Pseudomonas syringae van Hall. Res. Microbiol. 1991, 142, 995–1003. [Google Scholar] [CrossRef]
- Maraite, H.; Weyns, J. Pseudomonas syringae pv. aptata and pv. atrofaciens, Specific Pathovars or Members of pv. Syringae? In Pseudomonas Syringae Pathovars and Related Pathogens; Rudolph, K., Burr, T.J., Mansfield, J.W., Stead, D., Vivian, A., von Kietzell, J., Eds.; Developments in Plant Pathology; Springer: Dordrecht, The Netherlands, 1997; pp. 515–520. ISBN 978-94-011-5472-7. [Google Scholar]
- Iacobellis, N.S.; Figliuolo, G.; Janse, J.; Scortichini, M.; Ciuffreda, G. Characterization of Pseudomonas syringae pv. atrofaciens. In Pseudomonas syringae Pathovars and Related Pathogens; Rudolph, K., Burr, T.J., Mansfield, J.W., Stead, D., Vivian, A., von Kietzell, J., Eds.; Developments in Plant Pathology; Springer: Dordrecht, The Netherlands, 1997; pp. 500–504. ISBN 978-94-011-5472-7. [Google Scholar]
- Sato, M.; Nishiyama, K.; Shirata, A. Involvement of Plasmid DNA in the Productivity of Coronatine by Pseudomonas syringae pv. atropurpurea. Jpn. J. Phytopathol. 1983, 49, 522–528. [Google Scholar] [CrossRef]
- Psallidas, P.G. Pseudomonas syringae pv. avellanae pathovar nov., the Bacterium Causing Canker Disease on Corylus avellana. Plant Pathol. 1993, 42, 358–363. [Google Scholar] [CrossRef]
- Ménard, M.; Sutra, L.; Luisetti, J.; Prunier, J.P.; Gardan, L. Pseudomonas syringae pv. avii (pv. nov.), the Causal Agent of Bacterial Canker of Wild Cherries (Prunus avium) in France. Eur. J. Plant Pathol. 2003, 109, 565–576. [Google Scholar] [CrossRef]
- Roberts, S.j. A Note on Pseudomonas syringae pv. berberidis Infections of Berberis: Comparative Infection Studies in Attached and Detached Leaves. J. Appl. Bacteriol. 1985, 59, 369–374. [Google Scholar] [CrossRef]
- Takahashi, K.; Nishiyama, K.; Sato, M. Pseudomonas syringae pv. broussonetiae pv. nov., the Causal Agent of Bacterial Blight of Paper Mulberry (Broussonetia kazinoki×B. papyrifera). Jpn. J. Phytopathol. 1996, 62, 17–22. [Google Scholar] [CrossRef]
- Sato, M.; Watanabe, K.; Yazawa, M.; Takikawa, Y.; Nishiyama, K. Detection of New Ethylene-Producing Bacteria, Pseudomonas syringae pvs. cannabina and sesami, by PCR Amplification of Genes for the Ethylene-Forming Enzyme. Phytopathology 1997, 87, 1192–1196. [Google Scholar] [CrossRef] [PubMed]
- Takanashi, K.; Shimizu, K. Pseudomonas syringae pv. castaneae pv. nov., causal agent of bacterial canker of chestnut (Castanea crenata Sieb. et Zucc.). Ann. Phytopathol. Soc. Jpn. 1989, 55, 397–403. [Google Scholar] [CrossRef]
- Kamiunten, H.; Nakao, T.; Oshida, S. Pseudomonas syringae pv. cerasicola, pv. nov., the Causal Agent of Bacterial Gall of Cherry Tree. J. Gen. Plant Pathol. 2000, 66, 219–224. [Google Scholar] [CrossRef]
- Lavermicocca, P.; Lonigro, S.L.; Evidente, A.; Andolfi, A. Bacteriocin Production by Pseudomonas syringae pv. ciccaronei NCPPB2355. Isolation and Partial Characterization of the Antimicrobial Compound. J. Appl. Microbiol. 1999, 86, 257–265. [Google Scholar] [CrossRef]
- Toben, H.-M.; Rudoph, K. Pseudomonas syringae pv. coriandricola, Incitant of Bacterial Umbel Blight and Seed Decay of Coriander (Coriandrum sativum L.) in Germany. J. Phytopathol. 1996, 144, 169–178. [Google Scholar] [CrossRef]
- Harper, S.; Zewdie, N.; Brown, I.R.; Mansfield, J.W. Histological, Physiological and Genetical Studies of the Responses of Leaves and Pods of Phaseolus vulgaris to Three Races of Pseudomonas syringae pv. phaseolicola and to Pseudomonas syringae pv. coronafaciens. Physiol. Mol. Plant Pathol. 1987, 31, 153–172. [Google Scholar] [CrossRef]
- Scortichini, M.; Rossi, M.P.; Loreti, S.; Bosco, A.; Fiori, M.; Jackson, R.W.; Stead, D.E.; Aspin, A.; Marchesi, U.; Zini, M.; et al. Pseudomonas syringae pv. coryli, the Causal Agent of Bacterial Twig Dieback of Corylus avellana. Phytopathology 2005, 95, 1316–1324. [Google Scholar] [CrossRef] [PubMed]
- He, X.-Y.; Goto, M. Bacterial Needle Blight of Chinese Fir (Cunninghamia lanceolate Hook) Caused by Pseudomonas syringae pv. cunninghamiae pv. nov. Jpn. J. Phytopathol. 1995, 61, 38–40. [Google Scholar] [CrossRef]
- Oginh, C.; Kubo, Y.; Higuchi, H.; Takikawa, Y. Bacterial Gall Diseases of Himeyuzuriha (Daphniphyllum teijsmanni Z.) Caused by Pseudomonas syringae pv. daphniphylli pv. nov. J. Jpn. For. Soc. 1990, 72, 17–22. [Google Scholar] [CrossRef]
- Zdorovenko, E.L.; Zatonsky, G.V.; Kocharova, N.A.; Shashkov, A.S.; Knirel, Y.A.; Ovod, V.V. Structure of the O-Polysaccharide of Pseudomonas syringae pv. delphinii NCPPB 1879T Having Side Chains of 3-Acetamido-3,6-Dideoxy-D-Galactose Residues. Biochemistry 2002, 67, 558–565. [Google Scholar] [CrossRef]
- Ogimi, C.; Higuchi, H.; Takikawa, Y. Bacterial Gall Disease of Kakuremino (Dendropanax trifidus Mak.) Caused by Pseudomonas syrigae pv. dendropanacis pv. nov. Ann. Phytopathol. Soc. Jpn. 1988, 54, 296–302. [Google Scholar] [CrossRef]
- Young, J.M.; Triggs, C.m. Evaluation of Determinative Tests for Pathovars of Pseudomonas syringae van Hall 1902. J. Appl. Bacteriol. 1994, 77, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Kamiunten, H. Loss of a Plasmid in Pseudomonas syringae pv. eriobotryae Is Correlated with Change of Symptoms. Jpn. J. Phytopathol. 1990, 56, 645–650. [Google Scholar] [CrossRef]
- Kairu, G.M. Biochemical and Pathogenic Differences between Kenyan and Brazilian Isolates of Pseudomonas syringae pv. garcae. Plant Pathol. 1997, 46, 239–246. [Google Scholar] [CrossRef]
- Osman, S.F.; Fett, W.F.; Fishman, M.L. Exopolysaccharides of the Phytopathogen Pseudomonas syringae pv. glycinea. J. Bacteriol. 1986, 166, 66–71. [Google Scholar] [CrossRef]
- Arsenijević, M.; Vennete, R.J.; Maširević, S. Pseudomonas syringae pv. helianthi (Kawamura 1934) Dye, Wilkie et Young 1978, a Pathogen of Sunflower (Helianthus annuus L.). J. Phytopathol. 1994, 142, 199–208. [Google Scholar] [CrossRef]
- Fukuda, T.; Azegami, K.; Tabei, H. Histological studies on bacterial black node of barley and wheat caused by Pseudomonas syringae pv. japonica. Ann. Phytopathol. Soc. Jpn. 1990, 56, 252–256. [Google Scholar] [CrossRef]
- Yedidia, I.; Shoresh, M.; Kerem, Z.; Benhamou, N.; Kapulnik, Y.; Chet, I. Concomitant Induction of Systemic Resistance to Pseudomonas syringae pv. lachrymans in Cucumber by Trichoderma asperellum (T-203) and Accumulation of Phytoalexins. Appl. Environ. Microbiol. 2003, 69, 7343–7353. [Google Scholar] [CrossRef]
- Cuppels, D.A.; Ainsworth, T. Molecular and Physiological Characterization of Pseudomonas syringae pv. tomato and Pseudomonas syringae pv. maculicola Strains That Produce the Phytotoxin Coronatine. Appl. Environ. Microbiol. 1995, 61, 3530–3536. [Google Scholar] [CrossRef] [PubMed]
- Kiba, A.; Imanaka, Y.; Nakano, M.; Galis, I.; Hojo, Y.; Shinya, T.; Ohnishi, K.; Hikichi, Y. Silencing of Nicotiana benthamiana SEC14 Phospholipid Transfer Protein Reduced Jasmonic Acid Dependent Defense against Pseudomonas syringae. Plant Biotechnol. 2016, 33, 111–115. [Google Scholar] [CrossRef]
- Ratanapo, S.; Ngamjunyaporn, W.; Chulavatnatol, M. Interaction of a Mulberry Leaf Lectin with a Phytopathogenic Bacterium, P. syringae Pv mori. Plant Sci. 2001, 160, 739–744. [Google Scholar] [CrossRef]
- Sundin, G.W.; Jones, A.L.; Olson, B.D. Overwintering and Population Dynamics of Pseudomonas syringae pv. syringae and P.s. pv. morsprunorum on Sweet and Sour Cherry Trees. Can. J. Plant Pathol. 1988, 10, 281–288. [Google Scholar] [CrossRef]
- Ogimi, C.; Higuchi, H. Bacterial gall of yamamomo (Myrica rubra S. et Z.) caused by Pseudomonas syringae pv. myricae pv. nov. Ann. Phytopathol. Soc. Jpn. 1981, 47, 443–448. [Google Scholar] [CrossRef]
- Kuwata, H. Pseudomonas syringae pv. oryzae pv. nov., Causal Agent of Bacterial Halo Blight of Rice. Jpn. J. Phytopathol. 1985, 51, 212–218. [Google Scholar] [CrossRef]
- Kerkoud, M.; Manceau, C.; Paulin, J.P. Rapid Diagnosis of Pseudomonas syringae pv. papulans, the Causal Agent of Blister Spot of Apple, by Polymerase Chain Reaction Using Specifically Designed HrpL Gene Primers. Phytopathology 2002, 92, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Jose, P.A.; Krishnamoorthy, R.; Kwon, S.-W.; Janahiraman, V.; Senthilkumar, M.; Gopal, N.O.; Kumutha, K.; Anandham, R. Interference in Quorum Sensing and Virulence of the Phytopathogen Pseudomonas syringae pv. passiflorae by Bacillus and Variovorax Species. BioControl 2019, 64, 423–433. [Google Scholar] [CrossRef]
- Young, J.M. Pseudomonas syringae pv. persicae from Nectarine, Peach, and Japanese Plum in New Zealand1. EPPO Bull. 1988, 18, 141–151. [Google Scholar] [CrossRef]
- Lindgren, P.B.; Peet, R.C.; Panopoulos, N.J. Gene Cluster of Pseudomonas syringae pv. “phaseolicola” Controls Pathogenicity of Bean Plants and Hypersensitivity of Nonhost Plants. J. Bacteriol. 1986, 168, 512–522. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.J. Variation within Pseudomonas syringae pv. philadelphi, the Cause of a Leaf Spot of Philadelphus spp. J. Appl. Bacteriol. 1985, 59, 283–290. [Google Scholar] [CrossRef]
- Goto, M. Pseudomonas syringae pv. photiniae pv. nov., the Causal Agent of Bacterial Leaf Spot of Photinia glabra Maxim. Jpn. J. Phytopathol. 1983, 49, 457–462. [Google Scholar] [CrossRef]
- Taylor, J.D.; Bevan, J.R.; Crute, I.R.; Reader, S.L. Genetic Relationship between Races of Pseudomonas syringae pv. pisi and Cultivars of Pisum sativum. Plant Pathol. 1989, 38, 364–375. [Google Scholar] [CrossRef]
- Samson, R.; Shafik, H.; Benjama, A.; Gardan, L. Description of the Bacterium Causing Blight of Leek as Pseudomonas syringae pv. porri (pv. nov.). Phytopathology 1998, 88, 844–850. [Google Scholar] [CrossRef] [PubMed]
- Ogimi, C.; Kawano, C.; Higuchi, H.; Takikawa, Y. Bacterial Gall Disease of Sharinbai (Rhaphiolepis umbellata MARINO) Caused by Pseudomonas syringae pv. rhaphiolepidis pv. nov. J. Jpn. For. Soc. 1992, 74, 308–313. [Google Scholar] [CrossRef]
- Ovod, V.V.; Zdorovenko, E.L.; Shashkov, A.S.; Kocharova, N.A.; Knirel, Y.A. Structure of the O Polysaccharide and Serological Classification of Pseudomonas syringae pv. ribicola NCPPB 1010. Eur. J. Biochem. 2000, 267, 2372–2379. [Google Scholar] [CrossRef]
- Janse, J.D. Pathovar Discrimination within Pseudomonas syringae subsp. Savastanoi Using Whole Cell Fatty Acids and Pathogenicity as Criteria. Syst. Appl. Microbiol. 1991, 14, 79–84. [Google Scholar] [CrossRef]
- Firdous, S.S.; Asghar, R.; Irfan-ul-Haque, M.; Waheed, A.; Afzal, S.N.; Mirza, M.Y. Pathogenesis of Pseudomonas syringae pv. sesami Associated with Sesame (Sesamum indicum L.) Bacterial Leaf Spot. Pak. J. Bot. 2009, 41, 927–934. [Google Scholar]
- SATO, M.; WATANABE, K.; SATO, Y. Pseudomonas syringae pv. solidagae pv. nov., the Causal Agent of Bacterial Leaf Spot of Tall Goldenrod Solidago altissima L. J. Gen. Plant Pathol. 2001, 67, 303–308. [Google Scholar] [CrossRef]
- Barta, T.M.; Willis, D.K. Biological and Molecular Evidence That Pseudomonas syringae Pathovars coronafaciens, striafaciens and garcae Are Likely the Same Pathovar. J. Phytopathol. 2005, 153, 492–499. [Google Scholar] [CrossRef]
- Rahimian, H. The Occurrence of Bacterial Red Streak of Sugarcane Caused by Pseudomonas syringae pv. syringae in Iran. J. Phytopathol. 1995, 143, 321–324. [Google Scholar] [CrossRef]
- Turner, J.G.; Taha, R.R. Contribution of Tabtoxin to the Pathogenicity of Pseudomonas syringae pv. tabaci. Physiol. Plant Pathol. 1984, 25, 55–69. [Google Scholar] [CrossRef]
- Scortichini, M.; Marchesi, U.; Di Prospero, P. Genetic Relatedness among Pseudomonas avellanae, P. syringae pv. theae and P.s. pv. Actinidiae, and Their Identification. Eur. J. Plant Pathol. 2002, 108, 269–278. [Google Scholar] [CrossRef]
- Ogimi, C.; Higuchi, H.; Takikawa, Y. Bacterial Gall Disease of Urajiroenoki (Trema Orientalis BLUME) Caused by Pseudomonas syringae pv. Tremae pv. nov. J. Jpn. For. Soc. 1988, 70, 441–446. [Google Scholar] [CrossRef]
- Shinohara, H.; Moriwaki, J.; Kadota, I.; Nishiyama, K. Ability of pathotype strains of Pseudomonas syringae to produce coronatine. Ann. Phytopathol. Soc. Jpn. 1999, 65, 629–634. [Google Scholar] [CrossRef]
- Stead, D.E.; Stanford, H.; Aspin, A.; Weller, S.A. First Record of Pseudomonas syringae pv. viburni in the UK. Plant Pathol. 2006, 55, 571. [Google Scholar] [CrossRef]
- Bowden, R.L.; Percich, J. Etiology of Bacterial Leaf Streak of Wild Rice. Phytopathology 1983, 73, 640–645. [Google Scholar] [CrossRef]
- Xin, X.-F.; Nomura, K.; Aung, K.; Velásquez, A.C.; Yao, J.; Boutrot, F.; Chang, J.H.; Zipfel, C.; He, S.Y. Bacteria Establish an Aqueous Living Space in Plants Crucial for Virulence. Nature 2016, 539, 524–529. [Google Scholar] [CrossRef]
- Hu, Y.; Ding, Y.; Cai, B.; Qin, X.; Wu, J.; Yuan, M.; Wan, S.; Zhao, Y.; Xin, X.-F. Bacterial Effectors Manipulate Plant Abscisic Acid Signaling for Creation of an Aqueous Apoplast. Cell Host Microbe 2022, 30, 518–529.e6. [Google Scholar] [CrossRef]
- Yuan, M.; Jiang, Z.; Bi, G.; Nomura, K.; Liu, M.; Wang, Y.; Cai, B.; Zhou, J.-M.; He, S.Y.; Xin, X.-F. Pattern-Recognition Receptors Are Required for NLR-Mediated Plant Immunity. Nature 2021, 592, 105–109. [Google Scholar] [CrossRef]
- Tyson, J.L.; Vergara, M.J.; Butler, R.C.; Seelye, J.F.; Morgan, E.R. Survival, Growth and Detection of Pseudomonas syringae pv. actinidiae in Actinidia in Vitro Cultures. N. Z. J. Crop Hortic. Sci. 2018, 46, 319–333. [Google Scholar] [CrossRef]
- Authority (EFSA), E.F.S.; Vogelaar, M.; Schenk, M.; Delbianco, A.; Graziosi, I.; Vos, S. Pest Survey Card on Pseudomonas syringae pv. actinidiae. EFSA Support. Publ. 2020, 17, 1986E. [Google Scholar] [CrossRef]
- Whalen, M.C.; Innes, R.W.; Bent, A.F.; Staskawicz, B.J. Identification of Pseudomonas syringae Pathogens of Arabidopsis and a Bacterial Locus Determining Avirulence on Both Arabidopsis and Soybean. Plant Cell 1991, 3, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Sankaran, S.; Mishra, A.; Ehsani, R.; Davis, C. A Review of Advanced Techniques for Detecting Plant Diseases. Comput. Electron. Agric. 2010, 72, 1–13. [Google Scholar] [CrossRef]
- Narayanasamy, P. Microbial Plant Pathogens-Detection and Disease Diagnosis: Bacterial and Phytoplasmal Pathogens, Vol.2; Springer: Dordrecht, The Netherlands, 2011; ISBN 978-90-481-9768-2. [Google Scholar]
- Roos, I.M.M.; Hattingh, M.J. Scanning Electron Microscopy of Pseudomonas syringae Pv, morsprunorum on Sweet Cherry Leaves. J. Phytopathol. 1983, 108, 18–25. [Google Scholar] [CrossRef]
- Silcock, D.J.; Waterhouse, R.N.; Glover, L.A.; Prosser, J.I.; Killham, K. Detection of a Single Genetically Modified Bacterial Cell in Soil by Using Charge Coupled Device-Enhanced Microscopy. Appl. Environ. Microbiol. 1992, 58, 2444–2448. [Google Scholar] [CrossRef] [PubMed]
- Van Vuurde, J.W.L.; Franken, A.A.J.M.; Birnbaum, Y.; Jochems, G. Characteristics of Immunofluorescence Microscopy and of Dilution-Plating to Detect Pseudomonas syringae pv. phaseolicola in Bean Seed Lots and for Risk Assessment of Field Incidence of Halo Blight. Neth. J. Plant Pathol. 1991, 97, 233–244. [Google Scholar] [CrossRef]
- Mohan, S.K.; Schaad, N.W. An Improved Agar Plating Assay for Detecting Pseudomonas syringae pv. syringae and P. s. pv. phaseolicola in Contaminated Bean Seed. Phytopathology 1987, 77, 1390–1395. [Google Scholar] [CrossRef]
- Schaad, N.W.; Cheong, S.S.; Tamaki, S.; Hatziloukas, E.; Panopoulos, N.J. A Combined Biological and Enzymatic Amplification (BIO-PCR) Technique to Detect Pseudomonas syringae pv. phaseolicola in Bean Seed Extracts. Phytopathology 1995, 85, 243–246. [Google Scholar] [CrossRef]
- Gitaitis, R.D.; Sasser, M.J.; Beaver, R.W.; McInnes, T.B.; Stall, R.E. Pectolytic Xanthomonads in Mixed Infections with Pseudomonas syringae pv. syringae, P. syringae pv. tomato, and Xanthomonas campestris pv. vesicatoria in Tomato and Pepper Transplants. Phytopathology 1987, 77, 611–615. [Google Scholar] [CrossRef]
- Frampton, R.A.; Taylor, C.; Holguín Moreno, A.V.; Visnovsky, S.B.; Petty, N.K.; Pitman, A.R.; Fineran, P.C. Identification of Bacteriophages for Biocontrol of the Kiwifruit Canker Phytopathogen Pseudomonas syringae pv. actinidiae. Appl. Environ. Microbiol. 2014, 80, 2216–2228. [Google Scholar] [CrossRef]
- Martino, G.; Holtappels, D.; Vallino, M.; Chiapello, M.; Turina, M.; Lavigne, R.; Wagemans, J.; Ciuffo, M. Molecular Characterization and Taxonomic Assignment of Three Phage Isolates from a Collection Infecting Pseudomonas syringae pv. actinidiae and P. syringae pv. phaseolicola from Northern Italy. Viruses 2021, 13, 2083. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-D.; Kang, H.-J.; Chai, A.-L.; Shi, Y.-X.; Xie, X.-W.; Li, L.; Li, B.-J. Development of a Loop-Mediated Isothermal Amplification (LAMP) Assay for Rapid Detection of Pseudomonas syringae pv. tomato in Planta. Eur. J. Plant Pathol. 2020, 156, 739–750. [Google Scholar] [CrossRef]
- Lobato, I.M.; O’Sullivan, C.K. Recombinase Polymerase Amplification: Basics, Applications and Recent Advances. TrAC Trends Anal. Chem. 2018, 98, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Strayer-Scherer, A.; Jones, J.B.; Paret, M.L. Paret Recombinase Polymerase Amplification Assay for Field Detection of Tomato Bacterial Spot Pathogens. Phytopathology 2019, 109, 690–700. [Google Scholar] [CrossRef] [PubMed]
- Kant, P.; Fruzangohar, M.; Mann, R.; Rodoni, B.; Hollaway, G.; Rosewarne, G. Development and Application of a Loop-Mediated Isothermal Amplification (LAMP) Assay for the Detection of Pseudomonas syringae Pathovars pisi and syringae. Agriculture 2021, 11, 875. [Google Scholar] [CrossRef]
- Díaz-Cruz, G.A.; Smith, C.M.; Wiebe, K.F.; Villanueva, S.M.; Klonowski, A.R.; Cassone, B.J. Applications of Next-Generation Sequencing for Large-Scale Pathogen Diagnoses in Soybean. Plant Dis. 2019, 103, 1075–1083. [Google Scholar] [CrossRef]
- Hu, S.; Li, X.; Yin, X.; Li, R.; Zhang, R.; Zang, J.; Liu, Y. Species-Specific Identification of Pseudomonas Based on 16S-23S RRNA Gene Internal Transcribed Spacer (ITS) and Its Combined Application with next-Generation Sequencing. BMC Microbiol. 2022, 22, 188. [Google Scholar] [CrossRef] [PubMed]
- Cuppels, D.A.; Moore, R.A.; Morris, V.L. Construction and Use of a Nonradioactive DNA Hybridization Probe for Detection of Pseudomonas syringae pv. tomato on Tomato Plants. Appl. Environ. Microbiol. 1990, 56, 1743–1749. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Yan, Q.; Chen, J.; He, Y.; Wang, J.; Zhang, H.; Yu, Z.; Li, L. Molecular Characterization of an Ice Nucleation Protein Variant (InaQ) from Pseudomonas syringae and the Analysis of Its Transmembrane Transport Activity in Escherichia coli. Int. J. Biol. Sci. 2012, 8, 1097–1108. [Google Scholar] [CrossRef]
- Muvingi, M.; Муфарo, М.; Slovareva, O.Y.; Юрьевна, С.О.; Zargar, M.; Мейсам, З. Identification of Pseudomonas fuscovaginae, Pseudomonas syringae and Xanthomonas translucens in wheat seeds using PCR. RUDN J. Agron. Anim. Ind. 2022, 17, 473–483. [Google Scholar] [CrossRef]
- Balestra, G.M.; Taratufolo, M.C.; Vinatzer, B.A.; Mazzaglia, A. A Multiplex PCR Assay for Detection of Pseudomonas syringae pv. actinidiae and Differentiation of Populations with Different Geographic Origin. Plant Dis. 2013, 97, 472–478. [Google Scholar] [CrossRef]
- Rufián, J.S.; Sánchez-Romero, M.-A.; López-Márquez, D.; Macho, A.P.; Mansfield, J.W.; Arnold, D.L.; Ruiz-Albert, J.; Casadesús, J.; Beuzón, C.R. Pseudomonas syringae Differentiates into Phenotypically Distinct Subpopulations During Colonization of a Plant Host. Environ. Microbiol. 2016, 18, 3593–3605. [Google Scholar] [CrossRef]
- Khan, M.M.T.; Pyle, B.H.; Camper, A.K. Specific and Rapid Enumeration of Viable but Nonculturable and Viable-Culturable Gram-Negative Bacteria by Using Flow Cytometry. Appl. Environ. Microbiol. 2010, 76, 5088–5096. [Google Scholar] [CrossRef]
- Rufián, J.S.; Ruiz-Albert, J.; Beuzón, C.R. Fluorescently Labeled Pseudomonas syringae DC3000 and 1449b Wild-Type Strains Constitutively Expressing Either EGFP, ECFP, or DsRED. Micropublication Biol. 2022, 2022, 595. [Google Scholar] [CrossRef]
- Cimmino, A.; Iannaccone, M.; Petriccione, M.; Masi, M.; Evidente, M.; Capparelli, R.; Scortichini, M.; Evidente, A. An ELISA Method to Identify the Phytotoxic Pseudomonas syringae pv. actinidiae Exopolysaccharides: A Tool for Rapid Immunochemical Detection of Kiwifruit Bacterial Canker. Phytochem. Lett. 2017, 19, 136–140. [Google Scholar] [CrossRef]
- Wang, W.; Feng, M.; Kong, D.; Liu, L.; Song, S.; Xu, C. Development of an Immunochromatographic Strip for the Rapid Detection of Pseudomonas syringae pv. maculicola in Broccoli and Radish Seeds. Food Agric. Immunol. 2015, 26, 738–745. [Google Scholar] [CrossRef]
- Bonants, P. Advances in Testing Seed Health. In Advances in Seed Science and Technology for More Sustainable Crop Production; Burleigh Dodds Science Publishing Limited: Cambridge, UK, 2022; pp. 233–264. [Google Scholar] [CrossRef]
- Riffaud, C.M.-H.; Morris, C.E. Detection of Pseudomonas syringae pv. aptata in Irrigation Water Retention Basins by Immunofluorescence Colony-Staining. Eur. J. Plant Pathol. 2002, 108, 539–545. [Google Scholar] [CrossRef]
- Jung, H.-C.; Lebeault, J.-M.; Pan, J.-G. Surface Display of Zymomonas Mobilis Levansucrase by Using the Ice-Nucleation Protein of Pseudomonas syringae. Nat. Biotechnol. 1998, 16, 576–580. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Hu, Y.; Qin, K.; Yang, X.; Jia, Z.; Li, Q.; Chen, H.; Yang, H. A Serological Approach for the Identification of the Effector Hopz5 of Pseudomonas syringae pv. actinidiae: A Tool for the Rapid Immunodetection of Kiwifruit Bacterial Canker. J. Plant Pathol. 2018, 100, 171–177. [Google Scholar] [CrossRef]
- Rico, A.; McCraw, S.L.; Preston, G.M. The Metabolic Interface between Pseudomonas syringae and Plant Cells. Curr. Opin. Microbiol. 2011, 14, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Purahong, W.; Orrù, L.; Donati, I.; Perpetuini, G.; Cellini, A.; Lamontanara, A.; Michelotti, V.; Tacconi, G.; Spinelli, F. Plant Microbiome and Its Link to Plant Health: Host Species, Organs and Pseudomonas syringae pv. actinidiae Infection Shaping Bacterial Phyllosphere Communities of Kiwifruit Plants. Front. Plant Sci. 2018, 9, 1563. [Google Scholar] [CrossRef]
- López-Gresa, M.P.; Torres, C.; Campos, L.; Lisón, P.; Rodrigo, I.; Bellés, J.M.; Conejero, V. Identification of Defence Metabolites in Tomato Plants Infected by the Bacterial Pathogen Pseudomonas syringae. Environ. Exp. Bot. 2011, 74, 216–228. [Google Scholar] [CrossRef]
- Scortichini, M.; Ferrante, P.; Marcelletti, S.; Petriccione, M. Omics, Epidemiology and Integrated Approach for the Coexistence with Bacterial Canker of Kiwifruit, Caused by Pseudomonas syringae pv. actinidiae. Ital. J. Agron. 2014, 9, 163–165. [Google Scholar] [CrossRef]
- Nobori, T.; Wang, Y.; Wu, J.; Stolze, S.C.; Tsuda, Y.; Finkemeier, I.; Nakagami, H.; Tsuda, K. In Planta Bacterial Multi-Omics Analysis Illuminates Regulatory Principles Underlying Plant-Pathogen Interactions. bioRxiv 2019. [Google Scholar] [CrossRef]
- Guilbaud, C.; Morris, C.E.; Barakat, M.; Ortet, P.; Berge, O. Isolation and Identification of Pseudomonas syringae Facilitated by a PCR Targeting the Whole P. syringae Group. FEMS Microbiol. Ecol. 2016, 92, fiv146. [Google Scholar] [CrossRef]
- Bultreys, A.; Gheysen, I.; Maraite, H.; de Hoffmann, E. Characterization of Fluorescent and Nonfluorescent Peptide Siderophores Produced by Pseudomonas syringae Strains and Their Potential Use in Strain Identification. Appl. Environ. Microbiol. 2001, 67, 1718–1727. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Liu, Y.; Wang, N.R.; Haney, C.H. Mechanisms in Plant–Microbiome Interactions: Lessons from Model Systems. Curr. Opin. Plant Biol. 2021, 62, 102003. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Moreno, L.; Pineda, M.; Soukupová, J.; Macho, A.P.; Beuzón, C.R.; Barón, M.; Ramos, C. Early Detection of Bean Infection by Pseudomonas syringae in Asymptomatic Leaf Areas Using Chlorophyll Fluorescence Imaging. Photosynth. Res. 2008, 96, 27–35. [Google Scholar] [CrossRef]
- Iqbal, M.J.; Goodwin, P.H.; Leonardos, E.D.; Grodzinski, B. Spatial and Temporal Changes in Chlorophyll Fluorescence Images of Nicotiana benthamiana Leaves Following Inoculation with Pseudomonas syringae pv. tabaci. Plant Pathol. 2012, 61, 1052–1062. [Google Scholar] [CrossRef]
- Huang, L. Pseudomonas syringae pv. actinidiae Chemotaxis and the Kiwifruit Phyllosphere. Ph.D. Thesis, Open Access Te Herenga Waka-Victoria University of Wellington, Wellington, New Zealand, 2022. [Google Scholar]
- Hupp, S.; Rosenkranz, M.; Bonfig, K.; Pandey, C.; Roitsch, T. Noninvasive Phenotyping of Plant–Pathogen Interaction: Consecutive In Situ Imaging of Fluorescing Pseudomonas syringae, Plant Phenolic Fluorescence, and Chlorophyll Fluorescence in Arabidopsis Leaves. Front. Plant Sci. 2019, 10, 1239. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.; Mowat, A.; Bollen, A.; Whelan, B. Early Season Detection and Mapping of Pseudomonas syringae pv. actinidae Infected Kiwifruit (Actinidia sp.) Orchards. N. Z. J. Crop Hortic. Sci. 2014, 42, 303–311. [Google Scholar] [CrossRef]
- Cebula, Z.; Żołędowska, S.; Dziąbowska, K.; Skwarecka, M.; Malinowska, N.; Białobrzeska, W.; Czaczyk, E.; Siuzdak, K.; Sawczak, M.; Bogdanowicz, R.; et al. Detection of the Plant Pathogen Pseudomonas syringae pv. lachrymans on Antibody-Modified Gold Electrodes by Electrochemical Impedance Spectroscopy. Sensors 2019, 19, 5411. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Hulin, M.T.; Brain, P.; Mansfield, J.W.; Jackson, R.W.; Harrison, R.J. Rapid, Automated Detection of Stem Canker Symptoms in Woody Perennials Using Artificial Neural Network Analysis. Plant Methods 2015, 11, 57. [Google Scholar] [CrossRef]
- Furci, L.; Pascual-Pardo, D.; Ton, J. A Rapid and Non-Destructive Method for Spatial–Temporal Quantification of Colonization by Pseudomonas syringae pv. tomato DC3000 in Arabidopsis and Tomato. Plant Methods 2021, 17, 126. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Q.; Cui, H.; Li, Y.; Xu, N.; Lu, T.; Chen, J.; Penuelas, J.; Hu, B.; Qian, H. Composition Identification and Functional Verification of Bacterial Community in Disease-Suppressive Soils by Machine Learning. Environ. Microbiol. 2022, 24, 3405–3419. [Google Scholar] [CrossRef]
- Almeida, R.N.D.; Greenberg, M.; Bundalovic-Torma, C.; Martel, A.; Wang, P.W.; Middleton, M.A.; Chatterton, S.; Desveaux, D.; Guttman, D.S. Predictive Modeling of Pseudomonas syringae Virulence on Bean Using Gradient Boosted Decision Trees. PLoS Pathog. 2022, 18, e1010716. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.; Zhang, J.E.; Jiang, Y.P.; Wang, R.L.; Wu, R.S. Predicting the Potential Distribution of Pseudomonas syringae pv. actinidiae in China Using Ensemble Models. Plant Pathol. 2020, 69, 120–131. [Google Scholar] [CrossRef]
- Misas-Villamil, J.C.; Kolodziejek, I.; van der Hoorn, R.A.L. Pseudomonas syringae Colonizes Distant Tissues in Nicotiana benthamiana through Xylem Vessels. Plant J. 2011, 67, 774–782. [Google Scholar] [CrossRef] [PubMed]
- Farias, G.A.; Olmedilla, A.; Gallegos, M. Visualization and Characterization of Pseudomonas syringae pv. tomato DC3000 Pellicles. Microb. Biotechnol. 2019, 12, 688–702. [Google Scholar] [CrossRef] [PubMed]
- Dorati, F.; Barrett, G.A.; Sanchez-Contreras, M.; Arseneault, T.; José, M.S.; Studholme, D.J.; Murillo, J.; Caballero, P.; Waterfield, N.R.; Arnold, D.L.; et al. Coping with Environmental Eukaryotes; Identification of Pseudomonas syringae Genes during the Interaction with Alternative Hosts or Predators. Microorganisms 2018, 6, 32. [Google Scholar] [CrossRef]
- Daher, R.K.; Stewart, G.; Boissinot, M.; Bergeron, M.G. Recombinase Polymerase Amplification for Diagnostic Applications. Clin. Chem. 2016, 62, 947–958. [Google Scholar] [CrossRef]
- Rees-George, J.; Vanneste, J.L.; Cornish, D.A.; Pushparajah, I.P.S.; Yu, J.; Templeton, M.D.; Everett, K.R. Detection of Pseudomonas syringae pv. actinidiae Using Polymerase Chain Reaction (PCR) Primers Based on the 16S–23S RDNA Intertranscribed Spacer Region and Comparison with PCR Primers Based on Other Gene Regions. Plant Pathol. 2010, 59, 453–464. [Google Scholar] [CrossRef]
- Suzaki, K.; Sawada, H.; Kisaki, G. Loop-Mediated Isothermal Amplification of Bacterial Effector Genes to Detect Pseudomonas syringae pv. actinidiae Biovars 1 and 3. J. Gen. Plant Pathol. 2022, 88, 2–9. [Google Scholar] [CrossRef]
- Li, G.-R.; Huang, G.-M.; Zhu, L.-H.; Lv, D.; Cao, B.; Liao, F.; Luo, J.-F. Loop-Mediated Isothermal Amplification (LAMP) Detection of Phytophthora hibernalis, P. syringae and P. cambivora. J. Plant Pathol. 2019, 101, 51–57. [Google Scholar] [CrossRef]
- Huber, D.; Voith von Voithenberg, L.; Kaigala, G.V. Fluorescence in Situ Hybridization (FISH): History, Limitations and What to Expect from Micro-Scale FISH? Micro Nano Eng. 2018, 1, 15–24. [Google Scholar] [CrossRef]
- Kanegane, H.; Hoshino, A.; Okano, T.; Yasumi, T.; Wada, T.; Takada, H.; Okada, S.; Yamashita, M.; Yeh, T.-W.; Nishikomori, R.; et al. Flow Cytometry-Based Diagnosis of Primary Immunodeficiency Diseases. Allergol. Int. 2018, 67, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Stepaniak, L.; Sørhaug, T.; Jedrychowski, L. ANALYSIS | Immunochemical. In Encyclopedia of Dairy Sciences; Roginski, H., Ed.; Elsevier: Oxford, UK, 2002; pp. 62–67. ISBN 978-0-12-227235-6. [Google Scholar]
- Gonzalez, A.; Gaines, M.; Gallegos, L.Y.; Guevara, R.; Gomez, F.A. Enzyme-Linked Immunosorbent Assays (ELISA) Based on Thread, Paper, and Fabric. Electrophoresis 2018, 39, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Chan, H.S.L.; Haddad, G.; Zheng, L.; Bradley, G.; Dalton, W.S.; Ling, V. Sensitive Immunofluorescence Detection of the Expression of P-Glycoprotein in Malignant Cells. Cytometry 1997, 29, 65–75. [Google Scholar] [CrossRef]
- Tian, X.; Zhao, Z.; Wang, H.; Qin, Y.; Wang, D.; Li, Y.; Wang, X.; Wang, L. An Immunochromatographic Test Strip for On-Site Detection of Ralstonia solanacearum in Tobacco Leaves and Soil. Anal. Biochem. 2022, 641, 114561. [Google Scholar] [CrossRef]
- Strimbu, K.; Tavel, J.A. What Are Biomarkers? Curr. Opin. HIV AIDS 2010, 5, 463–466. [Google Scholar] [CrossRef]
- Bakare, O.O.; Keyster, M.; Pretorius, A. Identification of Biomarkers for the Accurate and Sensitive Diagnosis of Three Bacterial Pneumonia Pathogens Using in Silico Approaches. BMC Mol. Cell Biol. 2020, 21, 82. [Google Scholar] [CrossRef]
- Dyussembayev, K.; Sambasivam, P.; Bar, I.; Brownlie, J.C.; Shiddiky, M.J.A.; Ford, R. Biosensor Technologies for Early Detection and Quantification of Plant Pathogens. Front. Chem. 2021, 9, 636245. [Google Scholar] [CrossRef]
- Steinhauser, D.; Kopka, J. Methods, Applications and Concepts of Metabolite Profiling: Primary Metabolism. In Plant Systems Biology; Baginsky, S., Fernie, A.R., Eds.; Experientia Supplementum; Birkhäuser: Basel, Switzerland, 2007; pp. 171–194. ISBN 978-3-7643-7439-6. [Google Scholar]
- Shah, S.H.; Kraus, W.E.; Newgard, C.B. Metabolomic Profiling for Identification of Novel Biomarkers and Mechanisms Related to Common Cardiovascular Diseases: Form and Function. Circulation 2012, 126, 1110–1120. [Google Scholar] [CrossRef]
- Hazrati, H.; Kudsk, P.; Ding, L.; Uthe, H.; Fomsgaard, I.S. Integrated LC–MS and GC–MS-Based Metabolomics Reveal the Effects of Plant Competition on the Rye Metabolome. J. Agric. Food Chem. 2022, 70, 3056–3066. [Google Scholar] [CrossRef]
- Ferentinos, K.P. Deep Learning Models for Plant Disease Detection and Diagnosis. Comput. Electron. Agric. 2018, 145, 311–318. [Google Scholar] [CrossRef]
- Reis-Pereira, M.; Tosin, R.; Martins, R.; Neves Dos Santos, F.; Tavares, F.; Cunha, M. Kiwi Plant Canker Diagnosis Using Hyperspectral Signal Processing and Machine Learning: Detecting Symptoms Caused by Pseudomonas syringae pv. actinidiae. Plants 2022, 11, 2154. [Google Scholar] [CrossRef] [PubMed]
- Byrd, A.L.; Segre, J.A. Adapting Koch’s Postulates. Science 2016, 351, 224–226. [Google Scholar] [CrossRef] [PubMed]
- Agrios, G.N. Plant Pathology, 5th ed.; Elsevier Academic Press: Amsterdam, The Netherland; Boston, MA, USA, 2005; ISBN 978-0-12-044565-3. [Google Scholar]
- Segre, J.A. What Does It Take to Satisfy Koch’s Postulates Two Centuries Later? Microbial Genomics and Propionibacteria Acnes. J. Investig. Dermatol. 2013, 133, 2141–2142. [Google Scholar] [CrossRef]
- Bhunjun, C.S.; Phillips, A.J.L.; Jayawardena, R.S.; Promputtha, I.; Hyde, K.D. Importance of Molecular Data to Identify Fungal Plant Pathogens and Guidelines for Pathogenicity Testing Based on Koch’s Postulates. Pathogens 2021, 10, 1096. [Google Scholar] [CrossRef]
- Scholthof, K.-B.G. The Disease Triangle: Pathogens, the Environment and Society. Nat. Rev. Microbiol. 2007, 5, 152–156. [Google Scholar] [CrossRef]
- The Disease Triangle: A Plant Pathological Paradigm Revisited. Available online: https://www.apsnet.org/edcenter/foreducators/TeachingNotes/Pages/DiseaseTriangle.aspx (accessed on 6 January 2023).
- Fry, W.E. Principles of Plant Disease Management; Academic Press: Cambridge, MA, USA, 2012; ISBN 978-0-08-091830-3. [Google Scholar]
- Yang, P.; Bokros, N.; Debolt, S.; Zhao, Z.; Xia, Y. Genome Sequence Source of Bacillus amyloliquefaciens Strain GD4a, a Bacterial Endophyte Associated with Switchgrass Plants. Phytobiomes J. 2022, 6, 354–357. [Google Scholar] [CrossRef]
- Koskey, G.; Mburu, S.W.; Awino, R.; Njeru, E.M.; Maingi, J.M. Potential Use of Beneficial Microorganisms for Soil Amelioration, Phytopathogen Biocontrol, and Sustainable Crop Production in Smallholder Agroecosystems. Front. Sustain. Food Syst. 2021, 5, 606308. [Google Scholar] [CrossRef]
- Romano, I.; Ventorino, V.; Pepe, O. Effectiveness of Plant Beneficial Microbes: Overview of the Methodological Approaches for the Assessment of Root Colonization and Persistence. Front. Plant Sci. 2020, 11, 6. [Google Scholar] [CrossRef]
- Sahib, M.R.; Yang, P.; Bokros, N.; Shapiro, N.; Woyke, T.; Kyrpides, N.C.; Xia, Y.; DeBolt, S. Improved Draft Genome Sequence of Microbacterium sp. Strain LKL04, a Bacterial Endophyte Associated with Switchgrass Plants. Microbiol. Resour. Announc. 2019, 8, e00927-19. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Bokros, N.; DeBolt, S.; Yang, P.; Xia, Y. Genome Sequence Resource of Bacillus sp. RRD69, a Beneficial Bacterial Endophyte Isolated from Switchgrass Plants. Mol. Plant-Microbe Interact. 2021, 34, 1320–1323. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced Systemic Resistance by Beneficial Microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef]
- Dellagi, A.; Quillere, I.; Hirel, B. Beneficial Soil-Borne Bacteria and Fungi: A Promising Way to Improve Plant Nitrogen Acquisition. J. Exp. Bot. 2020, 71, 4469–4479. [Google Scholar] [CrossRef]
- Rosenblueth, M.; Ormeño-Orrillo, E.; López-López, A.; Rogel, M.A.; Reyes-Hernández, B.J.; Martínez-Romero, J.C.; Reddy, P.M.; Martínez-Romero, E. Nitrogen Fixation in Cereals. Front. Microbiol. 2018, 9, 1794. [Google Scholar] [CrossRef] [PubMed]
- Fox, A.; Kwapinski, W.; Griffiths, B.S.; Schmalenberger, A. The Role of Sulfur- and Phosphorus-Mobilizing Bacteria in Biochar-Induced Growth Promotion of Lolium perenne. FEMS Microbiol. Ecol. 2014, 90, 78–91. [Google Scholar] [CrossRef]
- Gahan, J.; Schmalenberger, A. The Role of Bacteria and Mycorrhiza in Plant Sulfur Supply. Front. Plant Sci. 2014, 5, 723. [Google Scholar] [CrossRef] [PubMed]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial Phosphorus Solubilization and Its Potential for Use in Sustainable Agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef]
- Moretti, L.G.; Crusciol, C.A.C.; Bossolani, J.W.; Calonego, J.C.; Moreira, A.; Garcia, A.; Momesso, L.; Kuramae, E.E.; Hungria, M. Beneficial Microbial Species and Metabolites Alleviate Soybean Oxidative Damage and Increase Grain Yield during Short Dry Spells. Eur. J. Agron. 2021, 127, 126293. [Google Scholar] [CrossRef]
- Abdelaal, K.; AlKahtani, M.; Attia, K.; Hafez, Y.; Király, L.; Künstler, A. The Role of Plant Growth-Promoting Bacteria in Alleviating the Adverse Effects of Drought on Plants. Biology 2021, 10, 520. [Google Scholar] [CrossRef]
- Munir, N.; Hanif, M.; Abideen, Z.; Sohail, M.; El-Keblawy, A.; Radicetti, E.; Mancinelli, R.; Haider, G. Mechanisms and Strategies of Plant Microbiome Interactions to Mitigate Abiotic Stresses. Agronomy 2022, 12, 2069. [Google Scholar] [CrossRef]
- Porter, S.S.; Bantay, R.; Friel, C.A.; Garoutte, A.; Gdanetz, K.; Ibarreta, K.; Moore, B.M.; Shetty, P.; Siler, E.; Friesen, M.L. Beneficial Microbes Ameliorate Abiotic and Biotic Sources of Stress on Plants. Funct. Ecol. 2020, 34, 2075–2086. [Google Scholar] [CrossRef]
- Jain, A.; Sarsaiya, S.; Wu, Q.; Lu, Y.; Shi, J. A Review of Plant Leaf Fungal Diseases and Its Environment Speciation. Bioengineered 2019, 10, 409–424. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Selvaraj, P.; Lin, L.; Fang, W.; Wu, C.; Yang, P.; Zhang, J.; Abubakar, Y.S.; Yang, F.; Lu, G.; et al. Rab7/Retromer-Based Endolysosomal Trafficking Facilitates Effector Secretion and Host Invasion in Rice Blast. bioRxiv 2022. [Google Scholar] [CrossRef]
- Gilligan, C.A. An Epidemiological Framework for Disease Management. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 2002; Volume 38, pp. 1–64. [Google Scholar]
- Newhall, A.G. Herbert Hice Whetzel: Pioneer American Plant Pathologist. Annu. Rev. Phytopathol. 1980, 18, 27–36. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathovar | Country | Host | Plant Symptoms | Year | References |
|---|---|---|---|---|---|
| pv. aceris | Japan | Acer buergerianum Miq. ‘Tohkaeda’(trident maple) | Necrotic leaf spots | 1990 | NCPPB, [35] |
| pv. actinidiae | Japan | Actinidia chinensis (kiwifruit) | Stem cankers | 1990 | NCPPB, [36] |
| pv. aesculi | India | Aesculus indica (horse chestnut) | Stem cankers | 1990 | NCPPB, [37] |
| pv. antirrhini | UK | Antirrhinum majus (snapdragon) | Necrotic leaf spots | 1966 | NCPPB, [38] |
| pv. apii | USA | Apium graveolens var. dulce (celery) | Necrotic leaf spots | 1964 | NCPPB, [39] |
| pv. aptata | USA | Beta vulgaris (sugar beet) | Tissue blights | 1961 | NCPPB, [40] |
| pv. atrofaciens | New Zealand | Triticum aestivum (bread wheat) | Glume rots | 1974 | NCPPB, [41] |
| pv. atropurpurea | Japan | Lolium multiflorum (ryegrass) | Shoot-tip diebacks | 1971 | NCPPB, [42] |
| pv. avellanae | Greece | Corylus avellana (hazel) | Stem cankers | 1987 | NCPPB, [43] |
| pv. avii | UK | Prunus avium (wild cherry) | Necrotic leaf spots | 1959 | NCPPB, [44] |
| pv. berberidis | New Zealand | Berberis sp. (barberry) | Necrotic leaf spots | 1975 | NCPPB, [45] |
| pv. broussonetiae | Japan | Broussonetia kazinoki (kozo) | Shoot blights | 2008 | NCPPB, [46] |
| pv. cannabina | Hungary | Cannabis sativa (hemp) | Leaf and stem rots | 1960 | NCPPB, [47] |
| pv. castaneae | Japan | Castanea crenata (chestnut) | Leaf blights | 2008 | NCPPB, [48] |
| pv. cerasicola | Japan | Prunus × yedoensis (cherry tree) | Galls on trunks and twigs. | 2008 | NCPPB, [49] |
| pv. ciccaronei | Italy | Ceratonia siliqua (carob) | Stem cankers | 1971 | NCPPB, [50] |
| pv. coriandricola | Germany | Coriandrum sativum var. micocarpur (coriander) | Necrotic leaf spots | 1991 | NCPPB, [51] |
| pv. coronafaciens | UK | Avena sativa (oat) | Leaf blights | 1958 | NCPPB, [52] |
| pv. coryli | Italy | Corylus avellena (hazel) | Stem cankers | 2001 | NCPPB, [53] |
| pv. cunninghamiae | China | Cunninghamia lanceolata (Chinese fir) | Small brown spots with yellow halos on needles (leaves) | 2008 | NCPPB, [54] |
| pv. daphniphylli | Japan | Daphniphyllum teijsmanni (himeyuzuriha) | Galls on trunks and twigs. | 1989 | NCPPB, [55] |
| pv. delphinii | New Zealand | Delphinium sp. (candle larspur) | Stem cankers | 1966 | NCPPB, [56] |
| pv. dendropanacis | Japan | Dendropanax trifidus (ivy) | Stem cankers | 1986 | NCPPB, [57] |
| pv. dysoxyli | New Zealand | Dysoxylum sp. (kohekohe) | Frost damages | 1966 | NCPPB, [58] |
| pv. eriobotryae | USA | Eriobotrya japonica (loquat) | Spots and blisters on fruit | 1970 | NCPPB, [59] |
| pv. garcae | Brazil | Coffea arabica (coffee) | Leaf and stem rots | 1958 | NCPPB, [60] |
| pv. glycinea | New Zealand | Glycine max (soybean) | Leaf blights | 1971 | NCPPB, [61] |
| pv. helianthi | Mexico | Helianthus annuus (sunflower) | Necrotic leaf spots | 1974 | NCPPB, [62] |
| pv. hibisci | USA | Hibiscus rosa seinensis (hibiscus) | Necrotic leaf spots | 1990 | NCPPB, [31] |
| pv. japonica | Japan | Hordeum vulgare (barley) | Leaf blights | 1979 | NCPPB, [63] |
| pv. lachrymans | Hungary | Cucumis sativus (cucumber) | Necrotic leaf spots | 1960 | NCPPB, [64] |
| pv. maculicola | New Zealand | Brassica oleracea var. botrytis (cauliflower) | Necrotic leaf spots | 1967 | NCPPB, [65] |
| pv. mellea | Japan | Nicotiana tabacum (tobacco) | Necrotic leaf spots | 1971 | NCPPB, [66] |
| pv. mori | Hungary | Morus alba (mulberry) | Necrotic leaf spots | 1961 | NCPPB, [67] |
| pv. morsprunorum | Switzerland | Prunus armeniaca (apricot) | Dead dormant buds | 1971 | NCPPB, [68] |
| pv. myricae | Japan | Myrica rubra (yumberry) | Necrotic leaf spots | 1981 | NCPPB, [69] |
| pv. oryzae | Japan | Oryza sativa (rice) | Sheath brown rots | 1990 | NCPPB, [70] |
| pv. papulans | Canada | Malus sylvestris (forest apple) | Blister spots | 1975 | NCPPB, [71] |
| pv. passiflorae | New Zealand | Passiflora edulis (passion fruit) | Necrotic leaf spots | 1963 | NCPPB, [72] |
| pv. persicae | France | Prunus persica (peach) | Stem cankers | 1975 | NCPPB, [73] |
| pv. phaseolicola | Canada | Phaseolus vulgaris (bean) | Necrotic leaf spots | 1941 | NCPPB, [74] |
| pv. philadelphi | UK | Philadelphus coronarius (dogwood) | Necrotic leaf spots | 1983 | NCPPB, [75] |
| pv. photiniae | Japan | Photinia glabra (Japanese photinia) | Necrotic leaf spots | 1990 | NCPPB, [76] |
| pv. pisi | New Zealand | Pisum sativum (pea) | Necrotic leaf spots | 1974 | NCPPB, [77] |
| pv. porri | France | Allium porrum (leek) | Leaf blights | 1985 | NCPPB, [78] |
| pv. primulae | USA | Primula sp. (primrose) | Necrotic leaf spots | 1939 | NCPPB, [31] |
| pv. rhaphiolepidis | Japan | Raphiolepis umbellata (yeddo hawthorne) | Necrotic leaf spots | 1989 | NCPPB, [79] |
| pv. ribicola | USA | Ribes aureum (golden currant) | Necrotic leaf spots | 1961 | NCPPB, [80] |
| pv. nerii | Spain | Nerium oleander (oleander) | Brown leaf galls | 1983 | NCPPB, [81] |
| pv. sesami | Greece | Sesamum indicum (sesame) | Necrotic leaf spots | 1961 | NCPPB, [82] |
| pv. solidagae | Japan | Solidago altissima (goldenrod) | Defoliation and terminal diebacks | 2009 | NCPPB, [83] |
| pv. striafaciens | USA | Avena sp. (oats) | Stripe blights | 1966 | NCPPB, [84] |
| pv. syringae | Japan | Hordeum vulgare (barley) | Leaf blights | 1979 | NCPPB, [85] |
| pv. tabaci | Australia | Glycine max (soybean) | Necrotic leaf spots | 1975 | NCPPB, [86] |
| pv. tagetis | Zimbabwe | Tagetes erecta (marigold) | Necrotic leaf spots | 1972 | NCPPB, [1] |
| pv. theae | Japan | Thea sinensis (tea plant) | Shoot blights | 1974 | NCPPB, [87] |
| pv. tomato | UK | Lycopersicon esculentum (tomato) | Necrotic leaf spots | 1961 | NCPPB, [32] |
| pv. tremae | Japan | Trema orientalis (charcoal-tree) | Necrotic leaf spots | 1986 | NCPPB, [88] |
| pv. ulmi | Yugoslavia | Ulmus sp. (elm) | Necrotic leaf spots | 1959 | NCPPB, [89] |
| pv. viburni | USA | Viburnum sp. (cranberry bush) | Leaf and stem spots | 1966 | NCPPB, [90] |
| pv. zizaniae | USA | Zizania aquatica (wild rice) | Leaf streaks | 1990 | NCPPB, [91] |
| Method Type | Method | Advantages | Limitations | References |
|---|---|---|---|---|
| Conventional | Visual examination | Quick and easy to perform, onsite disease detection and diagnosis | Subjective, not sensitive enough at early stages | [96,97] |
| Conventional | Microscopy | High resolution, versatility | Sample preparation, sample size, shallow depth of field | [100,101,102] |
| Conventional | Culture plate | Relatively inexpensive, easy to use, isolation of individual bacterial species | Prone to contamination, not suitable for unculturable bacteria | [103,104] |
| Conventional | Phage typing | High specificity | Limited to certain bacteria, limited resolution, risk of contamination | [105,106,107] |
| Molecular | RPA (recombinase polymerase amplification) | High sensitivity, high specificity, rapid turnaround time, onsite disease detection and diagnosis | Limited multiplexing, low throughput, poor stability, high cost | [108,109,110] |
| Molecular | LAMP (loop-mediated isothermal amplification) | Onsite disease detection and diagnosis, simplicity | Limited multiplexing, limited commercial availability | [108,111] |
| Molecular | NGS (next-generation sequencing) | High throughput, large-scale, high resolution, versatility | Technical expertise, sample quality, data analysis, limited access | [112,113] |
| Molecular | FISH (fluorescence in-situ hybridization) | High sensitivity, high specificity, rapid, Easy to visualize | Photobleaching, autofluorescence, limited to specific sequences | [114,115] |
| Molecular | PCR (polymerase chain reaction) | Ease of use, quantitation possible, sensitivity, specificity, speed, versatility | PCR system affects the effectiveness, complexity, false positives | [104,116,117] |
| Serological | FCM (flow cytometry) | High throughput, multiple parameters, sensitivity, accuracy | Complexity, sample preparation, limited cell types, interference | [118,119,120] |
| Serological | ELISA (enzyme-linked immunosorbent assay) | Speed, ease of use, testing seed health, sensitivity, specificity, versatility | Expensive, complexity, limited scope, false positives | [121,122,123] |
| Serological | IF (immunofluorescence) | Sensitive and visualizable, multiplexing, ease of use | Photobleaching, requirement for specific antibodies, limited to surface-exposed molecules, need for specialized equipment | [102,124,125] |
| Serological | ImmunoStrip | Rapid, sensitive, specific, portable, easy to use | Specificity varies among products, expensive, a qualitative test, | [122,126] |
| Biomarker-based | Plant metabolite profiling | High specificity, early detection, high sensitivity, multiplexing capability, High throughput | Expensive, incomplete database, data analysis skills, complexity | [127,128,129] |
| Biomarker-based | Pathogen metabolite profiling | High specificity, early detection, high sensitivity, multiplexing capability, high throughput | Expensive, incomplete database, data analysis skills, limited to specific stages of infection, limited to specific pathogens | [130,131,132,133] |
| Biomarker-based | Microbiome analysis | High throughput, early detection, broad coverage | Expensive, incomplete database, data analysis skills, complexity, limited understanding, technical challenges | [128,134] |
| Vision-based | Hyperspectral imaging | Early-detection, can be used to study the effects of various factors, such as environmental conditions or treatment, on plant growth and development | Expensive, can be affected by factors such as light intensity and wavelength, may require the use of specialized equipment and software | [135,136,137,138] |
| Vision-based | Spectroscopic imaging | Early-detection, non-destructive, high spatial resolution, high sensitivity, rapid analysis | Expensive, limited depth penetration, complex sample preparation, need for calibration, interference from external factors | [139,140] |
| AI (artificial intelligence) | Machine learning | Speed, accuracy, cost-effectiveness | Lack of database, lack of algorithms, lack of understanding, dependence on high-quality data, lack of interpretability, bias | [141,142,143,144,145] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, P.; Zhao, L.; Gao, Y.G.; Xia, Y. Detection, Diagnosis, and Preventive Management of the Bacterial Plant Pathogen Pseudomonas syringae. Plants 2023, 12, 1765. https://doi.org/10.3390/plants12091765
Yang P, Zhao L, Gao YG, Xia Y. Detection, Diagnosis, and Preventive Management of the Bacterial Plant Pathogen Pseudomonas syringae. Plants. 2023; 12(9):1765. https://doi.org/10.3390/plants12091765
Chicago/Turabian StyleYang, Piao, Lijing Zhao, Yu Gary Gao, and Ye Xia. 2023. "Detection, Diagnosis, and Preventive Management of the Bacterial Plant Pathogen Pseudomonas syringae" Plants 12, no. 9: 1765. https://doi.org/10.3390/plants12091765
APA StyleYang, P., Zhao, L., Gao, Y. G., & Xia, Y. (2023). Detection, Diagnosis, and Preventive Management of the Bacterial Plant Pathogen Pseudomonas syringae. Plants, 12(9), 1765. https://doi.org/10.3390/plants12091765