
Bruchus rufimanus Boh. Effect on Broad Bean Seed Quality and the Infection Level of Seed-Borne Fungal Pathogens



Abstract
:1. Introduction
2. Results
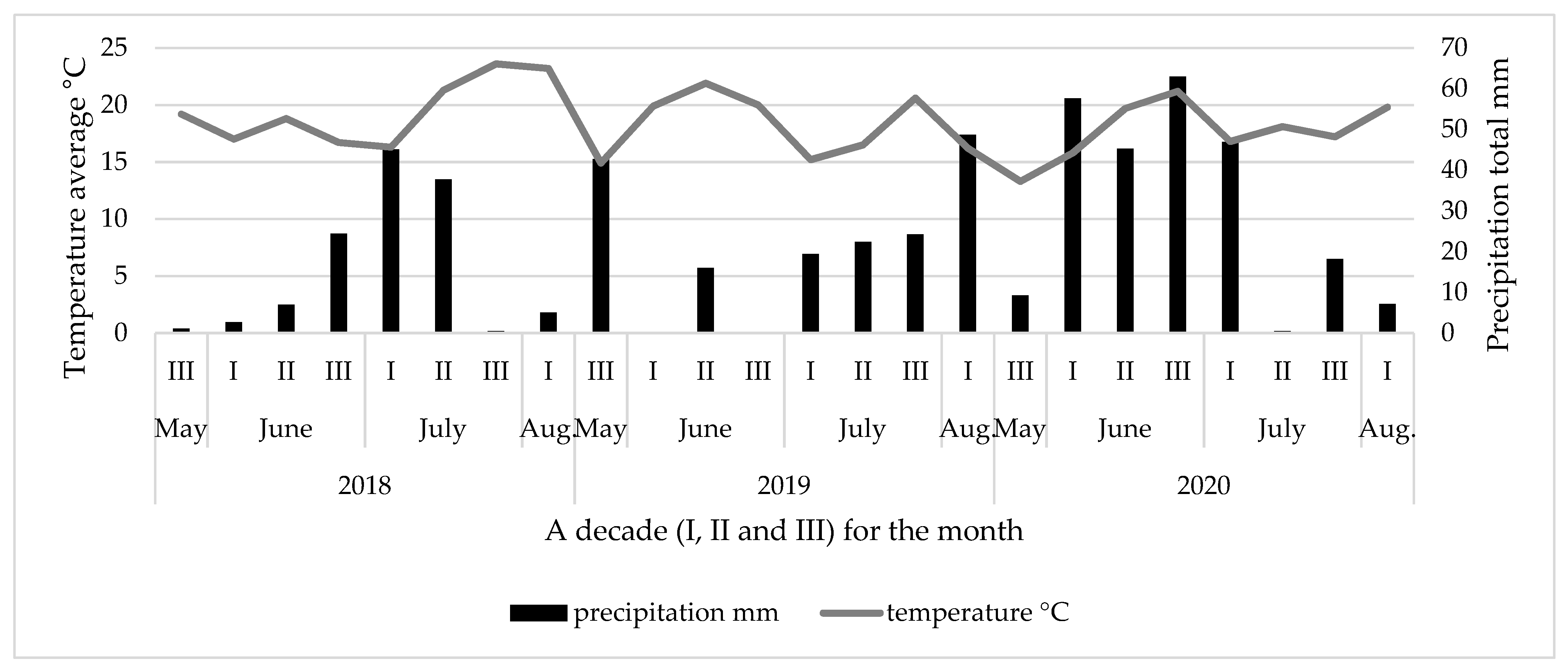
2.1. Weather Data
2.2. Seeds Germination and Seedling Vigor
2.3. Fungal Contamination on Seed
3. Discussion
4. Materials and Methods
4.1. Field Experiment
4.2. Seed Quality
4.3. Seed-Borne Pathogen Identification
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vitti, A.; Bevilacqua, V.; Logozzo, G.; Marzario, S.; Gioia, T.; Nuzzaci, M. Response of Two Local Common Bean Ecotypes of “Fagioli Di Sarconi” PGI (Phaseolus vulgaris L.) to Seed-Borne Pathogens and Environmental Change. Agronomy 2021, 11, 1924. [Google Scholar] [CrossRef]
- Dhull, S.B.; Kidwai, M.K.; Siddiq, M.; Sidhu, J.S. Faba (Broad) Bean Production, Processing, and Nutritional Profile. In Dry Beans and Pulses; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2022; pp. 359–381. ISBN 978-1-119-77680-2. [Google Scholar]
- Singh, D.V.; Nath, D.S.; Singh, S.P.; Mishra, U.; Singh, S. Character Association Correlation and Path Coefficient Analysis in Faba Bean (Vicia faba L.). Pharma Innov. 2021, 10, 517–520. [Google Scholar] [CrossRef]
- Bilalis, D.; Sidiras, N.; Economou, G.; Vakali, C. Effect of Different Levels of Wheat Straw Soil Surface Coverage on Weed Flora in Vicia faba Crops. J. Agron. Crop Sci. 2003, 189, 233–241. [Google Scholar] [CrossRef]
- Bangar, S.P.; Kajla, P. Introduction: Global Status and Production of Faba-Bean. In Faba Bean: Chemistry, Properties and Functionality; Punia Bangar, S., Bala Dhull, S., Eds.; Springer International Publishing: New York, NY, USA, 2022; pp. 1–15. ISBN 978-3-031-14587-2. [Google Scholar]
- OSP Official Statistics Portal. Sown Area of Agricultural Crops. Available online: https://osp.stat.gov.lt/statistiniu-rodikliu-analize?indicator=S9R086# (accessed on 5 January 2022).
- Prabhu, S.; Rajeswari, D. Nutritional and Biological Properties of Vicia faba L.: A Perspective Review. Int. Food Res. J. 2018, 25, 1332–1340. [Google Scholar]
- Dhull, S.B.; Kidwai, M.K.; Noor, R.; Chawla, P.; Rose, P.K. A Review of Nutritional Profile and Processing of Faba Bean (Vicia faba L.). Legume Sci. 2022, 4, e129. [Google Scholar] [CrossRef]
- Multari, S.; Stewart, D.; Russell, W.R. Potential of Fava Bean as Future Protein Supply to Partially Replace Meat Intake in the Human Diet. Compr. Rev. Food Sci. Food Saf. 2015, 14, 511–522. [Google Scholar] [CrossRef]
- Kusvuran, A.; Parlak, E.L.; SağlamtiMur, T. Biomass Yield of Faba Bean (Vicia faba L.) and Its Mixture with Some Grasses (Poaceae). Turk. J. Agric. Nat. Sci. 2015, 2, 178–184. [Google Scholar]
- Nohwar, N.; Khandare, R.V.; Desai, N.S. Isolation and Characterization of Salinity Tolerant Nitrogen Fixing Bacteria from Sesbania sesban (L) Root Nodules. Biocatal. Agric. Biotechnol. 2019, 21, 101325. [Google Scholar] [CrossRef]
- Jensen, E.S.; Peoples, M.B.; Hauggaard-Nielsen, H. Faba Bean in Cropping Systems. Field Crops Res. 2010, 115, 203–216. [Google Scholar] [CrossRef]
- Stoddard, F.L.; Nicholas, A.H.; Rubiales, D.; Thomas, J.; Villegas-Fernández, A.M. Integrated Pest Management in Faba Bean. Field Crops Res. 2010, 115, 308–318. [Google Scholar] [CrossRef]
- Ward, R. The Biology and Ecology of Bruchus Rufimanus (Bean Seed Beetle). Ph.D. Thesis, Newcastle University, Newcastle, UK, 2018. [Google Scholar]
- Seidenglanz, M.; Huňady, I. Effects of Faba Bean (Vicia faba) Varieties on the Development of Bruchus Rufimanus. Czech J. Genet. Plant Breed. 2016, 52, 22–29. [Google Scholar] [CrossRef]
- Pölitz, B.; Reike, H.P. Studies on biology and infestation dynamics of the bean seed beetle (Coleoptera, Bruchidae: Bruchus rufimanus) in Saxony. Gesunde Pflanz. 2019, 71, 79–85. [Google Scholar] [CrossRef]
- Khelfane-Goucem, K.; Medjdoub-Bensaad, F. Impact of Bruchus Rufimanus Infestation upon Broad Bean Seeds Germination. Adv. Environ. Biol. 2016, 10, 144–152. [Google Scholar]
- Epperlein, K. Investigations of the Damage of Broad Bean Weevil Bruchus rufimanus Bohem on Broad Bean Seed. Anz. Fuer Schaedlingskunde Pflanzenschutz Umweltschutz 1992, 65, 147–150. [Google Scholar] [CrossRef]
- Kaniuczak, Z. Seed Damage of Field Bean (Vicia faba L. Var. Minor Harz.) Caused by Bean Weevils (Bruchus rufimanus Boh.) (Coleoptera: Bruchidae). J. Plant Prot. Res. 2004, 44, 125–129. [Google Scholar]
- Jones FG, W.; Jones, M.G. Seed Beetles: Bruchus rufimanus Boh. In Pests of Field Crops; St. Martin’s Press: New York, NY, USA; London, UK, 1964; pp. 141–142. [Google Scholar]
- Roubinet, E. Management of the Broad Bean Weevil (Bruchus rufimanus Boh.) in Faba Bean (Vicia faba L.); Swedish University of Agricultural Sciences: Uppsala, Sweden, 2016; p. 23. [Google Scholar]
- Abdulwehab, S.A.; El-Nagerabi, S.a.F.; Elshafie, A.E. Leguminicolous Fungi Associated with Some Seeds of Sudanese Legumes. Biodivers. J. Biol. Divers. 2015, 16, 269–280. [Google Scholar] [CrossRef]
- Saleem, A.; Ebrahim, M.K.H. Production of Amylase by Fungi Isolated from Legume Seeds Collected in Almadinah Almunawwarah, Saudi Arabia. J. Taibah Univ. Sci. 2014, 8, 90–97. [Google Scholar] [CrossRef]
- Ahmed, S.; Imtiaz, M.; Kumar, S.; Malhotra, R.; Maalouf, F. Impact of Climate Change and Variability on Diseases of Food Legumes in the Dry Areas. In Proceedings of the Food Security and Climate Change in Dry Areas, Amman, Jordan, 1–4 February 2011; pp. 157–165. [Google Scholar]
- Elwakil, M.A.; El-Refai, I.M.; Awadallah, O.A.; El-Metwally, M.A.; Mohammed, M.S. Seed-Borne Pathogens of Faba Bean in Egypt: Detection and Pathogencity. Plant Pathol. J. 2009, 8, 90–97. [Google Scholar] [CrossRef]
- Biddle, A.; Cattlin, N. Pests, Diseases and Disorders of Peas and Beans. In A Colour Handbook; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Embaby, E.M.; Abdel-Galil, M.M. Seed Borne Fungi and Mycotoxins Associated with Some Legume Seeds in Egypt. J. Appl. Sci. Res. 2006, 2, 1064–1071. [Google Scholar]
- Singh, D.; Sangle, U.; Kumar, B.; Tripathi, H.; Singh, K.; Gupta, A. Integrated Disease Management of Faba Bean (Vicia faba L.). In Faba Bean (Vicia faba L.): A Potential Leguminous Crop of India; Indian Council of Agricultural Research: New Delhi, India, 2012; pp. 279–301. [Google Scholar]
- Rasiukeviciute, N.; Kelpsiene, J. The Impact of Fusarium Graminearum Infection on Different Plant Seeds. Res. Rural. Dev. 2018, 2, 114–118. [Google Scholar] [CrossRef]
- Kaiser, W.J. Fungi Associated with the Seeds of Commercial Lentils from the U.S. Pacific Northwest. Plant Dis. 1992, 76, 605–610. [Google Scholar] [CrossRef]
- Martín, I.; Gálvez, L.; Guasch, L.; Palmero, D. Fungal Pathogens and Seed Storage in the Dry State. Plants 2022, 11, 3167. [Google Scholar] [CrossRef] [PubMed]
- Nikolova, I.; Georgieva, N. Evaluation of Damage Caused by Bruchus pisorum L. on Some Parameters Related to Seed Quality of Pea Cultivars. J. Mt. Agric. Balk. 2015, 18, 452–477. [Google Scholar]
- Boughdad, A.; Laugé, G. Infestation et Pertes Des Graines de Vicia faba L. Dues à Bruchus rufimanus Boh. (Coleoptera, Bruchidae) Au Maroc | INRA. Al Awamia 1997, 97, 27–39. [Google Scholar]
- Titouhi, F.; Amri, M.; Jemâa, J. Susceptibility of Some Tunisian Broad Bean Varieties to Injuries Caused by the Broad Bean Weevil Bruchus Rufimanus in North Tunisia. Tunis. J. Plant Prot. 2015, 10, 55–61. [Google Scholar]
- Fox, C.W.; Wallin, W.G.; Bush, M.L.; Czesak, M.E.; Messina, F.J. Effects of Seed Beetles on the Performance of Desert Legumes Depend on Host Species, Plant Stage, and Beetle Density. J. Arid. Environ. 2012, 80, 10–16. [Google Scholar] [CrossRef]
- Beck, C.W.; Blumer, L. A Handbook on Bean Beetles; National Science Foundation: Alexandria, VA, USA, 2014; p. 17. [Google Scholar]
- Sinha, R.N. Effects of Stored-Product Beetle Infestation on Fat Acidity, Seed Germination, and Microflora of Wheat1. J. Econ. Entomol. 1983, 76, 813–817. [Google Scholar] [CrossRef]
- Jha, A.N.; Srivastava, C.; Dwivedi, V. Effect of Infestation of Rice Weevil (Sitophilus oryzae L.). Indian J. Entomol. 2014, 76, 74–76. [Google Scholar]
- Nikolova, I.M. Response of Pea Varieties to Damage Degree of Pea Weevil, Bruchus pisorum L. Scientifica 2016, 2016, e8053860. [Google Scholar] [CrossRef]
- Mateus, C.; Mexia, A.; Duarte, I.; Pereira, G.; Tavares de Sousa, M. Evaluation of Damage Caused by Bruchids (Coleoptera: Bruchidae) on Peas (Pisum sativum L.). Int. Symp. Plant 2010, 917, 125–132. [Google Scholar] [CrossRef]
- Segers, A.; Caparros Megido, R.; Lognay, G.; Francis, F. Overview of Bruchus Rufimanus Boheman 1833 (Coleoptera: Chrysomelidae): Biology, Chemical Ecology and Semiochemical Opportunities in Integrated Pest Management Programs. Crop Prot. 2021, 140, 105411. [Google Scholar] [CrossRef]
- Bonal, R.; Muñoz, A.; Díaz, M. Satiation of Predispersal Seed Predators: The Importance of Considering Both Plant and Seed Levels. Evol. Ecol. 2007, 21, 367–380. [Google Scholar] [CrossRef]
- Dubey, S.C.; Patel, B. Mode of Perpetuation and Spread of Alternaria Blight of Broad Bean. Indian Phytopathol. 2000, 53, 175–177. [Google Scholar]
- Bhutta, A.R. Seed-borne disease problems of legume crops in pakistan. Biol. Sci.-PJSIR 2000, 43, 249–254. [Google Scholar]
- Alves da Silva, S.; Fonseca Alvarenga Pereira, R.G.; de Azevedo Lira, N.; Micotti da Glória, E.; Chalfoun, S.M.; Batista, L.R. Fungi Associated to Beans Infested with Coffee Berry Borer and the Risk of Ochratoxin A. Food Control 2020, 113, 107204. [Google Scholar] [CrossRef]
- Zingales, V.; Taroncher, M.; Martino, P.A.; Ruiz, M.-J.; Caloni, F. Climate Change and Effects on Molds and Mycotoxins. Toxins 2022, 14, 445. [Google Scholar] [CrossRef]
- Paterson, R.R.M.; Lima, N.; Taniwaki, M.H. Coffee, Mycotoxins and Climate Change. Food Res. Int. 2014, 61, 1–15. [Google Scholar] [CrossRef]
- Pszczółkowska, A.; Okorski, A.; Fordoński, G.; Kotecki, A.; Kozak, M.; Dzienis, G. Effect of Weather Conditions on Yield and Health Status of Faba Bean Seeds in Poland. Agronomy 2020, 10, 48. [Google Scholar] [CrossRef]
- Ghareib, O.A.E.; Aly, A.Z.; Zaid, M.I.A.; Zaki, K.I. Isolation, Identification and Frequency Occurrence of Faba Bean Chocolate Spotdisease and Its Associated Fungi in Different Governorates of Egypt. Plant Arch. 2021, 21, 465–473. [Google Scholar]
- Haddoudi, I.; Mhadhbi, H.; Gargouri, M.; Barhoumi, F.; Romdhane, S.B.; Mrabet, M. Occurrence of Fungal Diseases in Faba Bean (Vicia faba L.) under Salt and Drought Stress. Eur. J. Plant Pathol. 2021, 159, 385–398. [Google Scholar] [CrossRef]
- Honda, Y.; Rahman, M.Z.; Islam, S.Z.; Muroguchi, N. Leaf Spot Disease of Broad Bean Caused by Alternaria Tenuissima in Japan. Plant Dis. 2001, 85, 95. [Google Scholar] [CrossRef] [PubMed]
- Pérez, J.; Infante, F.; Vega, F.E.; Holguín, F.; MacÍAs, J.; Valle, J.; Nieto, G.; Peterson, S.W.; Kurtzman, C.P.; O’donnell, K. Mycobiota Associated with the Coffee Berry Borer (Hypothenemus hampei) in Mexico. Mycol. Res. 2003, 107, 879–887. [Google Scholar] [CrossRef]
- Czerniakowski, Z.W.; Błażej, J.; Olbrycht, T. Increased Incidence of Major Diseases and Pests on Horse Bean (Vicia faba L. Var. Minor Harz.). Plant Breed. Seed Sci. 1996, 40, 79–86. [Google Scholar]
- Tack, A.J.M.; Dicke, M. Plant Pathogens Structure Arthropod Communities across Multiple Spatial and Temporal Scales. Funct. Ecol. 2013, 27, 633–645. [Google Scholar] [CrossRef]
- Stout, M.J.; Thaler, J.S.; Thomma, B.P.H.J. Plant-Mediated Interactions between Pathogenic Microorganisms and Herbivorous Arthropods. Annu. Rev. Entomol. 2006, 51, 663–689. [Google Scholar] [CrossRef]
- ISTA, I. International Seed Testing Association; International Rules for Seed Testing: Bassersdorf, Switzerland, 2005. [Google Scholar]
- Ruan, S.; Xue, Q.; Tylkowska, K. The Influence of Priming on Germination of Rice (Oryza sativa L.) Seeds and Seedling Emergence and Performance in Flooded Soil. Seed Sci. Technol. 2002, 30, 61–67. [Google Scholar]
- Javaid, M.M.; Florentine, S.; Ali, H.H.; Weller, S. Effect of Environmental Factors on the Germination and Emergence of Salvia verbenaca L. Cultivars (Verbenaca and Vernalis): An Invasive Species in Semi-Arid and Arid Rangeland Regions. PLoS ONE 2018, 13, e0194319. [Google Scholar] [CrossRef]
- Abdul-Baki, A.A.; Anderson, J.D. Vigor Determination in Soybean Seed by Multiple Criteria. Crop Sci. 1973, 13, 630–633. [Google Scholar] [CrossRef]
- Sheppard, J.W.; Cockerell, V. ISTA Handbook of Method Validation for the Detection of Seed-Borne Pathogens; International Seed Testing Association: Zurich, Switzerland, 2000; p. 84. [Google Scholar]
- Mathur, S.B.; Kongsdal, O. Common Laboratory Seed Health Testing Methods for Detecting Fungi; International Seed Testing Association: Bassersdorf, Switzerland, 2003; p. 425. [Google Scholar]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Wiley-Blackwell: Hoboken, NJ, USA, 2006; p. 416. [Google Scholar]
- Johnston, A.; Booth, C. Plant Pathologist’s Pocket Book. Common Wealth Mycological Institute; Commonwealth Agricultural Bureaux: Wallingford, UK, 1983; p. 527. [Google Scholar]


{kind=link}
| Year | Germination Rate (%) | Germination Index | Thousand Grain Weight (g) | |||
|---|---|---|---|---|---|---|
| Non-Damaged Seeds | Damaged Seeds | Non-Damaged Seeds | Damaged Seeds | Non-Damaged Seeds | Damaged Seeds | |
| 2018 | 99.0 ± 0.37 a | 89.5 ± 1.67 b | 13.2 ± 0.13 a | 12.3 ± 0.60 b | 647.9 ± 16.53 a | 539.5 ± 9.69 b |
| 2019 | 94.5 ± 1.05 a | 69.0 ± 3.72 b | 12.5 ± 0.39 a | 9.2 ± 0.67 b | 609.3 ± 8.63 a | 472.8 ± 2.39 b |
| 2020 | 98.2 ± 2.20 a | 85.7 ± 3.91 b | 12.8 ± 0.47 a | 12.4 ± 0.60 a | 637.7 ± 7.06 a | 511.0 ± 4.85 b |
| Average | 97.2 | 81.4 | 12.8 | 11.3 | 631.7 | 507.7 |
| Year | Length (cm) | Weight (g) | Vigor Index | |||
|---|---|---|---|---|---|---|
| Non-Damaged Seeds | Damaged Seeds | Non-Damaged Seeds | Damaged Seeds | Non-Damaged Seeds | Damaged Seeds | |
| Shoots | ||||||
| 2018 | 26.1 ± 0.5 a | 25.5 ± 0.7 a | 17.8 ± 0.7 a | 15.5 ± 0.8 a | 2581.4 ± 57.6 a | 2276.8 ± 49.2 b |
| 2019 | 24.2 ± 0.7 a | 18.2 ± 1.4 b | 20.3 ± 0.5 a | 15.4 ± 1.3 b | 2289.7 ± 79.4 a | 1265.2 ± 121.2 b |
| 2020 | 23.4 ± 1.2 a | 17.9 ± 1.0 b | 17.3 ± 1.4 a | 11.6 ± 0.9 b | 2260.5 ± 137.7 a | 1676.1 ± 112.1 b |
| Average | 24.6 | 20.53 | 18.5 | 14.2 | 2377.2 | 1739.4 |
| Roots | ||||||
| 2018 | 17.2 ± 0.4 a | 13.6 ± 0.4 b | 5.0 ± 0.6 a | 4.4 ± 0.4 a | 1699.5 ± 39.3 a | 1222.6 ± 49 b |
| 2019 | 11.9 ± 0.7 a | 10.5 ± 0.5 a | 12.3 ± 0.3 a | 8.8 ± 0.6 b | 1114.6 ± 64.4 a | 752.1 ± 47.5 b |
| 2020 | 17.9 ± 0.6 a | 17.1 ± 0.6 a | 11.2 ± 0.6 a | 5.6 ± 0.3 b | 1680.2 ± 66.2 a | 1543.1 ± 65.9 a |
| Average | 15.7 | 13.7 | 9.5 | 6.3 | 1498.1 | 1172.6 |
| Year | Fungal Pathogen | Non-Sterilized Seeds | Sterilized Seeds | ||
|---|---|---|---|---|---|
| Damaged Seeds | Non-Damaged Seeds | Damaged Seeds | Non-Damaged Seeds | ||
| 2018 | Fusarium spp. | 5.8 ± 4.3 CD a | 1.5 ± 0.6 C ab | 3.0 ± 2.4 D ab | 0.0 ± 0.0 B b |
| n = 400 * | Alternaria spp. | 37.3 ± 5.9 B a | 27.0 ± 8.6 B a | 24.3 ± 9.8 B a | 1.8 ± 1.0 B b |
| Aspergillus spp. | 6.0 ± 1.4 CD a | 7.0 ± 0.8 C a | 0.3 ± 0.5 D b | 0.0 ± 0.0 B b | |
| Penicillium spp. | 44.0 ± 5.4 B a | 30.0 ± 0.8 B b | 13.3 ± 4.0 C c | 0.5 ± 0.6 B d | |
| Cladosporium spp. | 96.3 ± 2.6 A a | 64.5 ± 7.5 A b | 53.3 ± 7.2 A b | 24.0 ± 2.2 A c | |
| Mucor spp. | 14.3 ± 13.7 C a | 9.3 ± 6.8 C a | 0.0 ± 0.0 D a | 0.3 ± 0.5 B a | |
| Botrytis spp. | 0.5 ± 1.0 D ab | 0.0 ± 0.0 C b | 1.8 ± 1.0 D a | 0.0 ± 0.0 B b | |
| 2019 | Fusarium spp. | 1.3 ± 1.7 C ab | 0.3 ± 0.7 C b | 1.8 ± 1.7 C a | 0.3 ± 0.5 D b |
| n = 1200 * | Alternaria spp. | 50.7 ± 17.2 B a | 19.6 ± 13.4 C b | 59.1 ± 12.8 A a | 11.0 ± 9.3 A b |
| Aspergillus spp. | 3.3 ± 2.1 C a | 0.6 ± 0.9 C b | 0.3 ± 0.7 C b | 0.9 ± 1.1 CD b | |
| Penicillium spp. | 88.3 ± 10.1 A a | 75.9 ± 25.9 A a | 23.3 ± 18.2 B b | 5.8 ± 5.1 BC b | |
| Cladosporium spp. | 39.2 ± 22.1 B a | 48.2 ± 33.5 B a | 14.3 ± 5.7 B b | 8.4 ± 3.8 AB b | |
| Mucor spp. | 8.2 ± 10.8 C a | 3.3 ± 4.6 C a | 3.3 ± 3.3 C a | 2.7 ± 3.1 CD a | |
| Botrytis spp. | 0.1 ± 0.3 C b | 0.0 ± 0.0 C b | 0.8 ± 1.2 C a | 0.0 ± 0.0 D b | |
| 2020 | Fusarium spp. | 28.1 ± 10.1 C a | 0.8 ± 1.0 C c | 17.3 ± 6.0 C b | 0.4 ± 0.5 B c |
| n = 800 * | Alternaria spp. | 35.3 ± 6.2 BC b | 9.8 ± 3.5 C c | 52.1 ± 12.4 A a | 5.0 ± 4.1 B c |
| Aspergillus spp. | 1.0 ± 1.2 D a | 0.1 ± 0.4 C a | 0.3 ± 0.7 E a | 0.1 ± 0.4 B a | |
| Penicillium spp. | 43.6 ± 18.3 B a | 32.0 ± 33.2 B ab | 11.8 ± 9.2 CD bc | 1.9 ± 3.4 B c | |
| Cladosporium spp. | 91.1 ± 4.2 A a | 86.8 ± 13.6 A a | 30.6 ± 10.2 B b | 22.5 ± 19.5 A b | |
| Mucor spp. | 0.8 ± 1.4 D a | 0.1 ± 0.4 C a | 0.5 ± 0.8 E a | 0.0 ± 0.0 B a | |
| Botrytis spp. | 7.8 ± 3.3 D a | 0.6 ± 0.9 C b | 3.6 ± 4.2 DE b | 0.5 ± 0.8 B b | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almogdad, M.; Jonavičienė, A.; Semaškienė, R. Bruchus rufimanus Boh. Effect on Broad Bean Seed Quality and the Infection Level of Seed-Borne Fungal Pathogens. Plants 2023, 12, 1825. https://doi.org/10.3390/plants12091825
Almogdad M, Jonavičienė A, Semaškienė R. Bruchus rufimanus Boh. Effect on Broad Bean Seed Quality and the Infection Level of Seed-Borne Fungal Pathogens. Plants. 2023; 12(9):1825. https://doi.org/10.3390/plants12091825
Chicago/Turabian StyleAlmogdad, Mohammad, Akvilė Jonavičienė, and Roma Semaškienė. 2023. "Bruchus rufimanus Boh. Effect on Broad Bean Seed Quality and the Infection Level of Seed-Borne Fungal Pathogens" Plants 12, no. 9: 1825. https://doi.org/10.3390/plants12091825
APA StyleAlmogdad, M., Jonavičienė, A., & Semaškienė, R. (2023). Bruchus rufimanus Boh. Effect on Broad Bean Seed Quality and the Infection Level of Seed-Borne Fungal Pathogens. Plants, 12(9), 1825. https://doi.org/10.3390/plants12091825