
Rhizosphere Microbiota Promotes the Growth of Soybeans in a Saline–Alkali Environment under Plastic Film Mulching

Abstract
:1. Introduction
2. Results
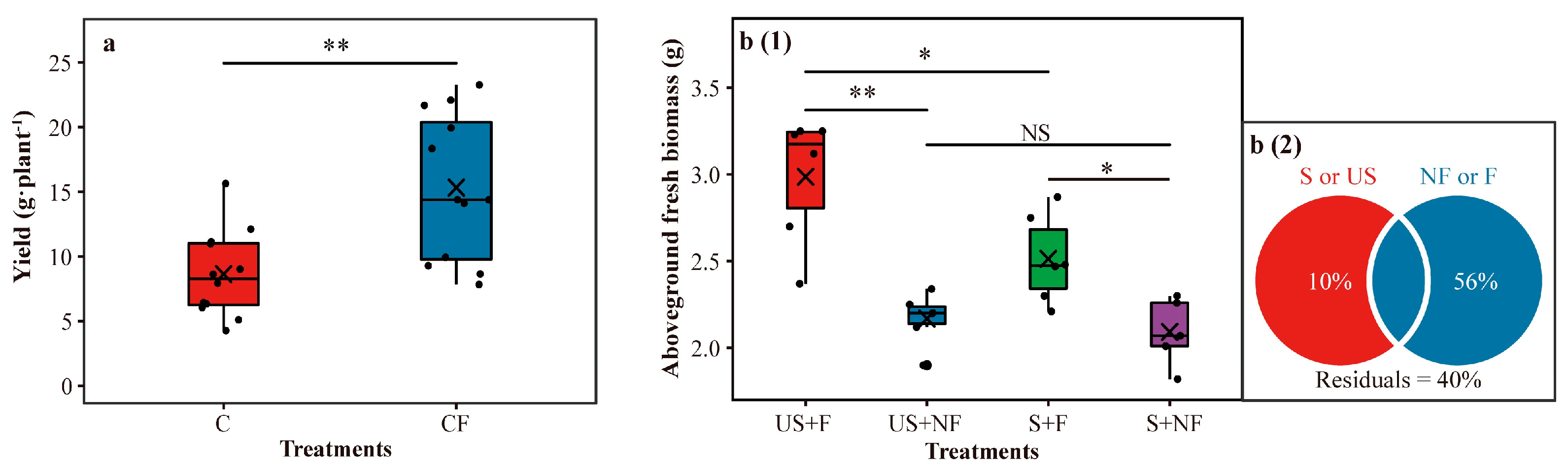
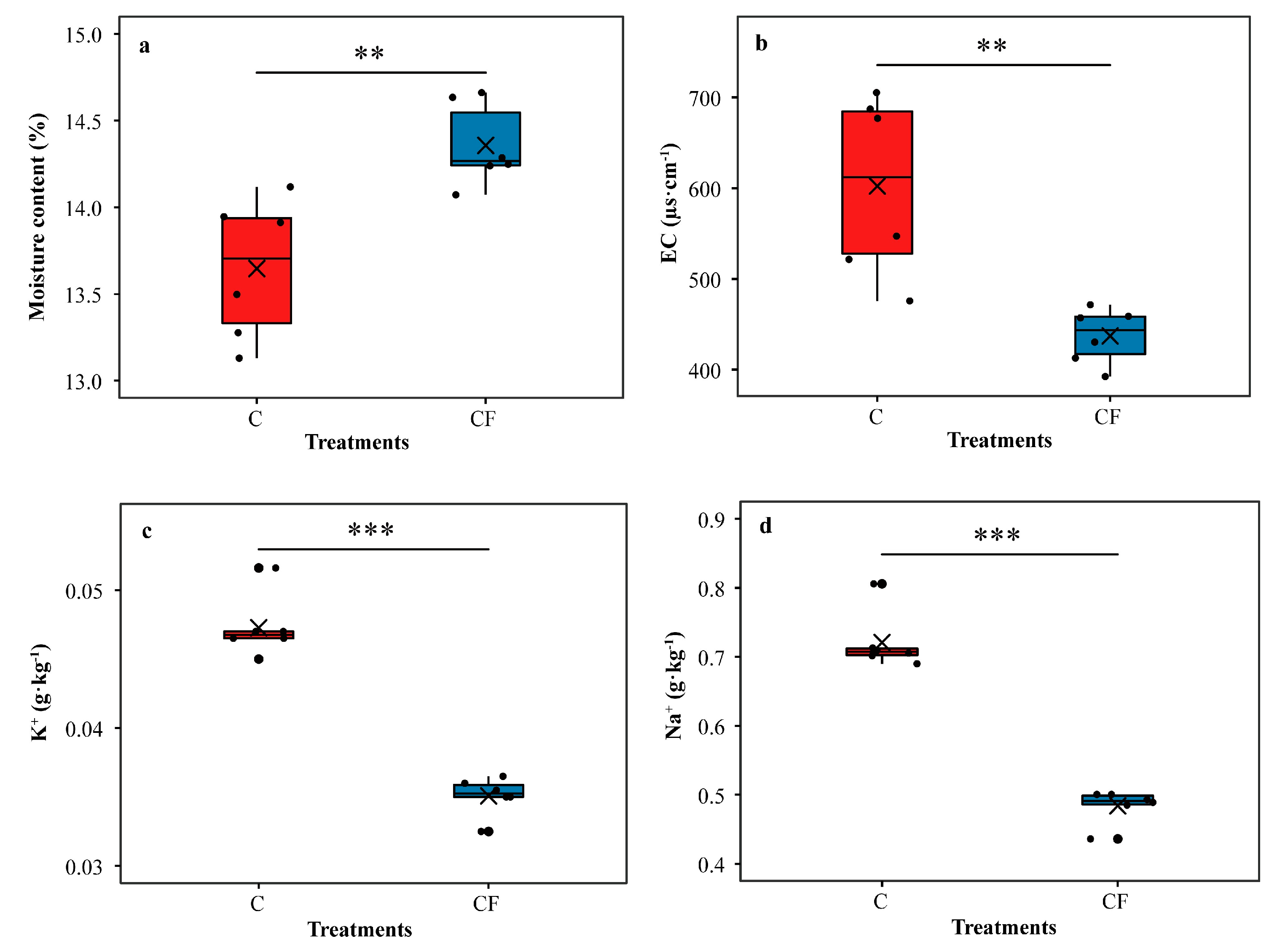
2.1. Soybean Yield and Bulk Soil Properties in the Field Survey
2.2. Results of the Greenhouse Mesocosm Experiment
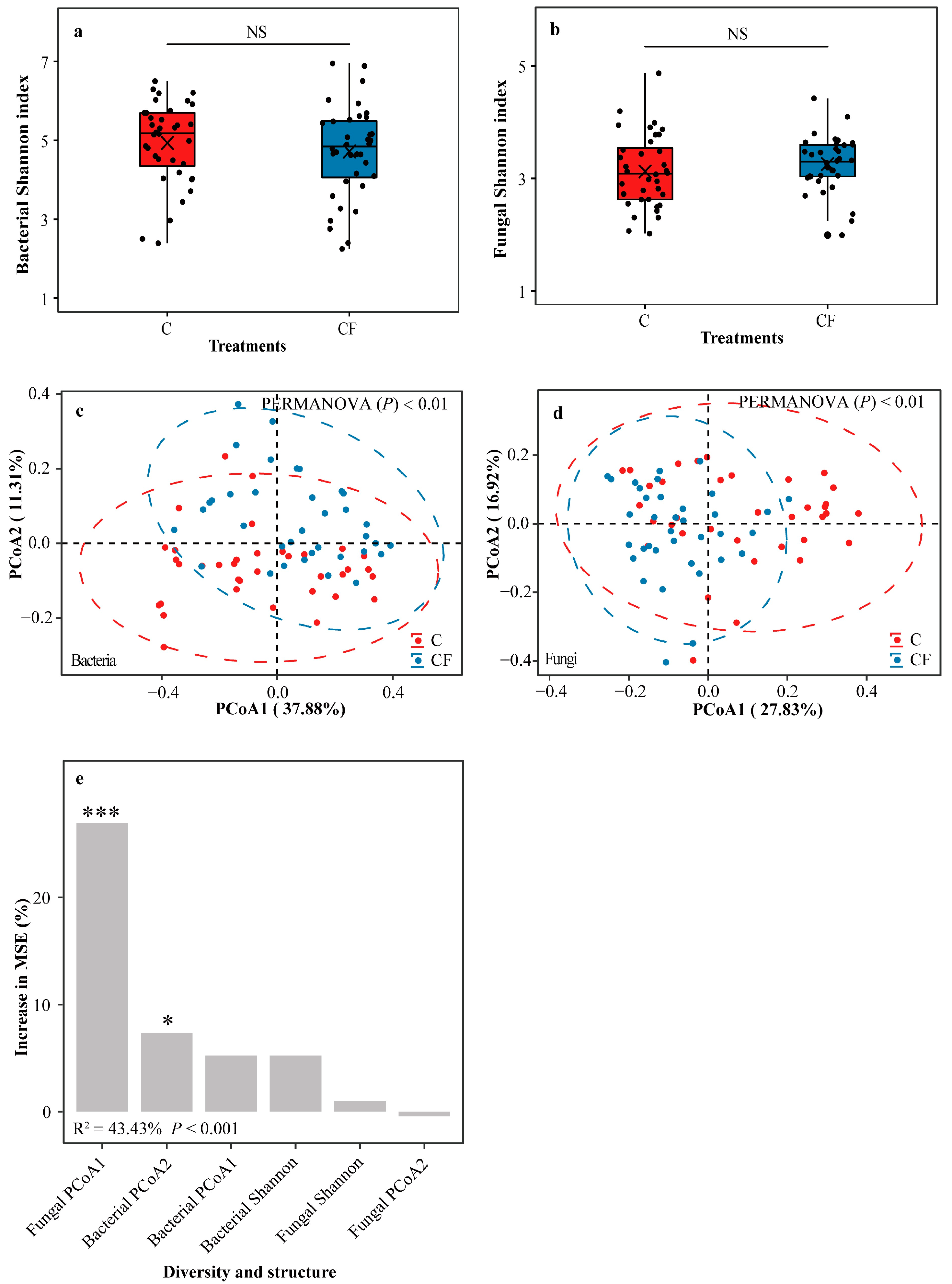
2.3. Diversity and Structure of Soybean Rhizosphere Microbiota
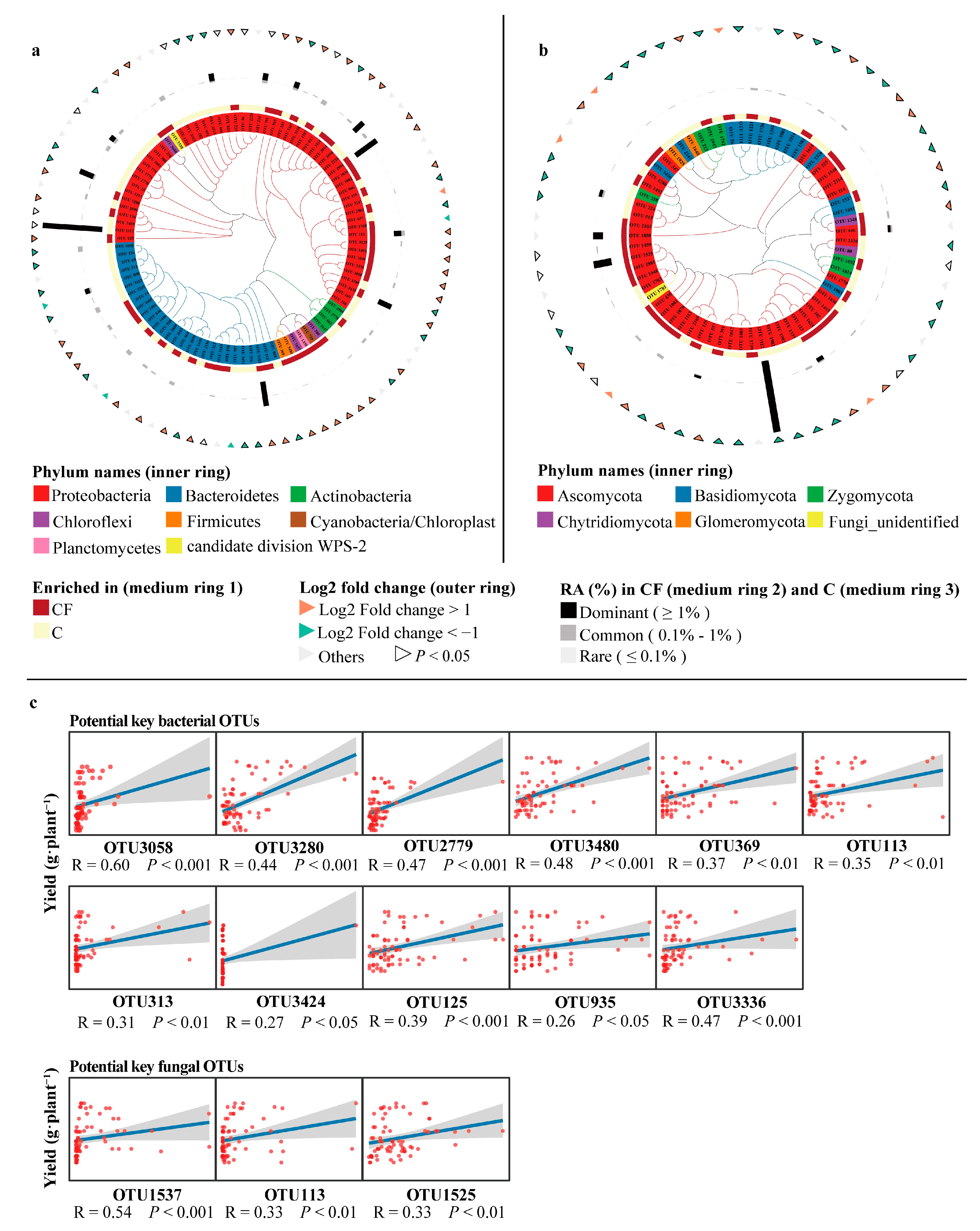
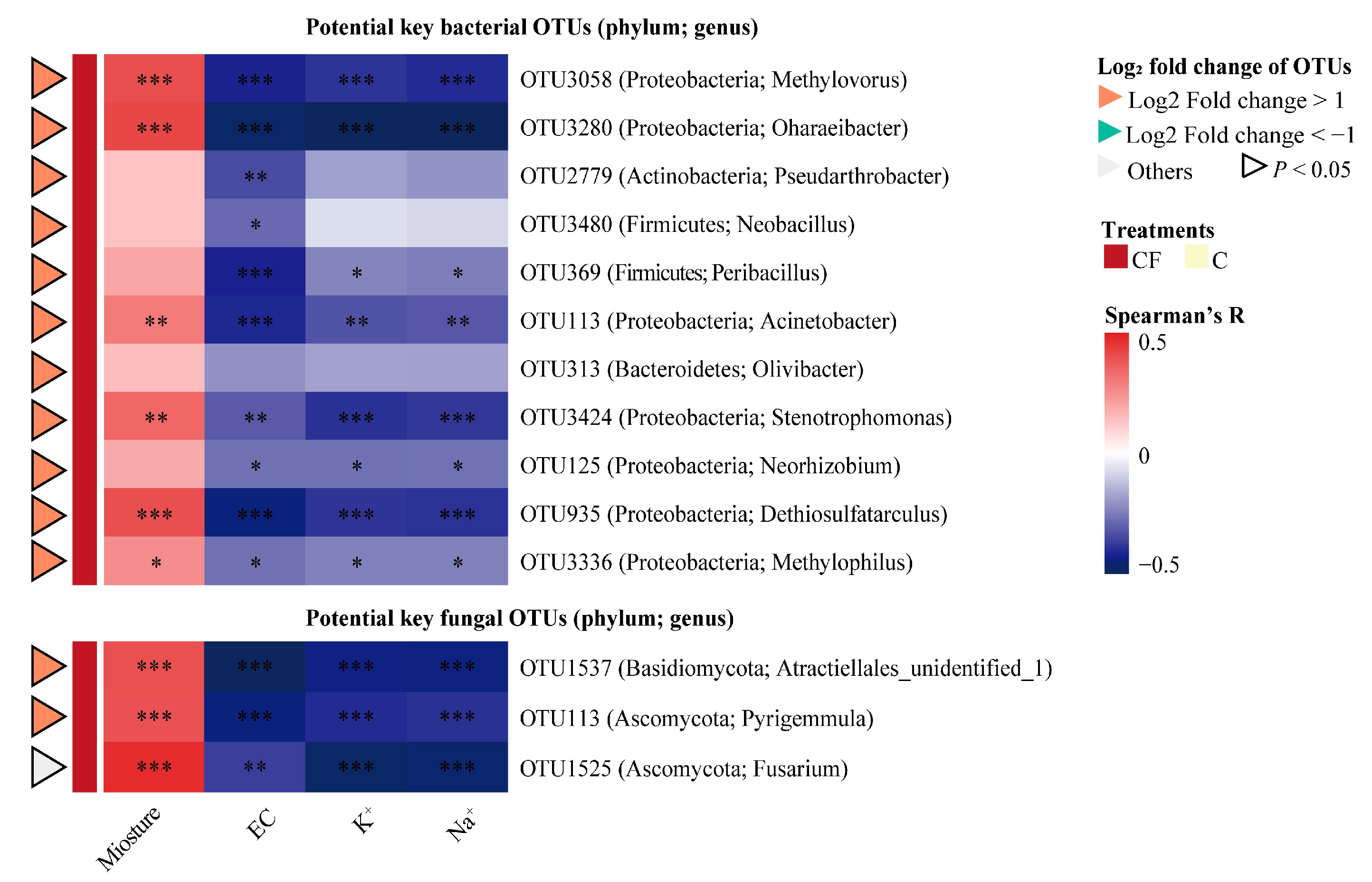
2.4. Key Microbial Taxa Associated with Yield and Soil Characteristics
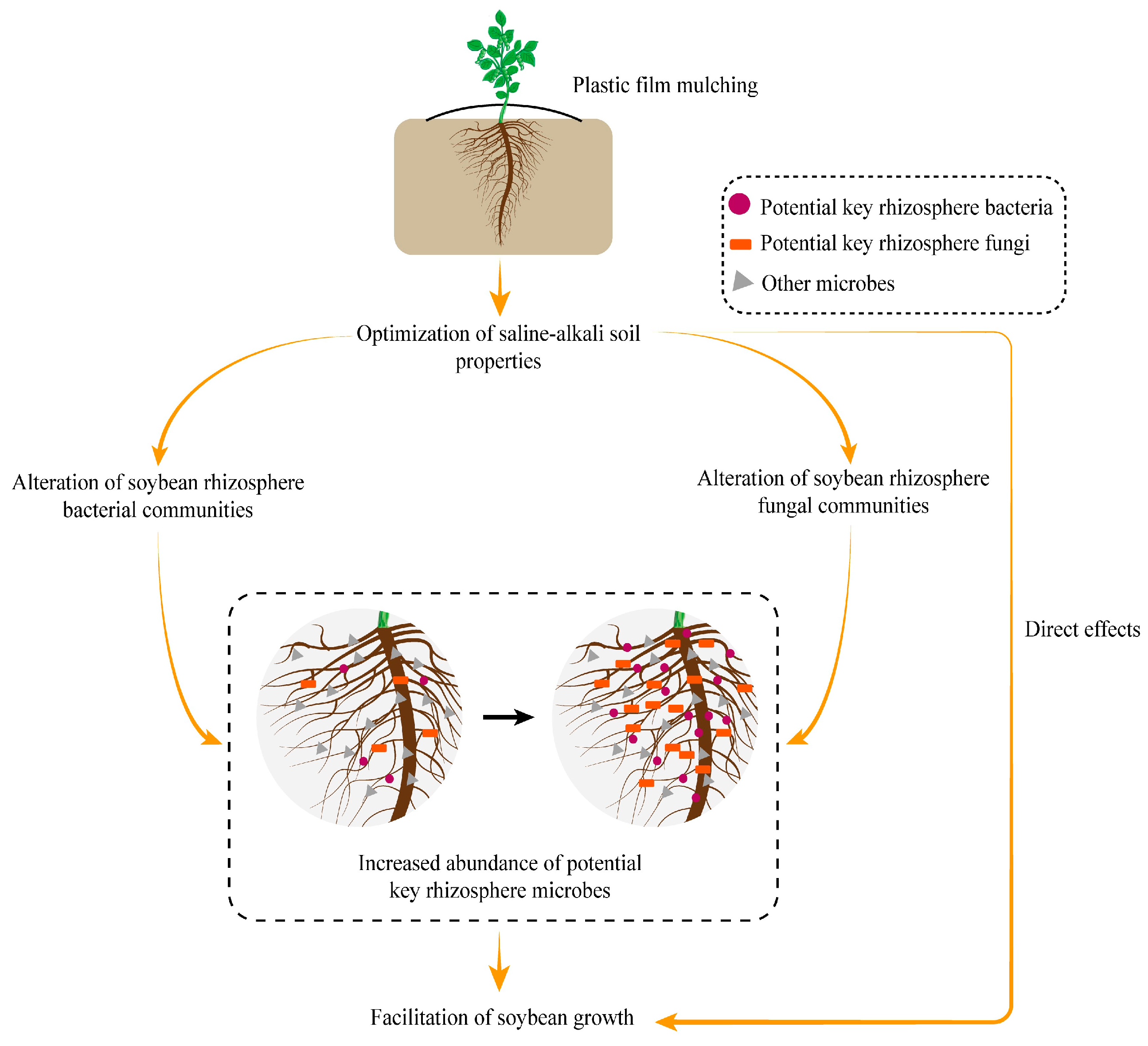
3. Discussion
4. Materials and Methods
4.1. Root and Soil Samplings
4.2. DNA Extraction, Sequencing and qPCR
4.3. Greenhouse Mesocosm Experiment
4.4. Bioinformatics and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Julkowska, M.M.; Testerink, C. Tuning plant signaling and growth to survive salt. Trends Plant Sci. 2015, 20, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Cuenca, M.-R.; Iglesias, D.J.; Forner-Giner, M.A.; Primo-Millo, E.; Legaz, F. The effect of sodium bicarbonate on plant performance and iron acquisition system of FA-5 (Forner-Alcaide 5) citrus seedlings. Acta Physiol. Plant. 2013, 35, 2833–2845. [Google Scholar] [CrossRef]
- Jin, H.; Plaha, P.; Park, J.; Hong, C.; Lee, I.; Yang, Z.; Jiang, G.; Kwak, S.; Liu, S.; Lee, J.; et al. Comparative EST profiles of leaf and root of Leymus chinensis, a xerophilous grass adapted to high pH sodic soil. Plant Sci. 2006, 170, 1081–1086. [Google Scholar] [CrossRef]
- Li, H.; Zhao, Q.; Huang, H. Current states and challenges of salt-affected soil remediation by cyanobacteria. Sci. Total Environ. 2019, 669, 258–272. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; La, S.; Zhang, X.; Gao, L.; Tian, Y. Salt-induced recruitment of specific root-associated bacterial consortium capable of enhancing plant adaptability to salt stress. ISME J. 2021, 15, 2865–2882. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.-M.; Wang, H.; Svetla, S.; Zhu, Y.-F.; Hu, Y.; Cheng, L.; Zhao, T.; Wang, Y.-X. Comparative physiological responses and adaptive strategies of apple Malus halliana to salt, alkali and saline-alkali stress. Sci. Hortic. 2019, 245, 154–162. [Google Scholar] [CrossRef]
- Wang, J.; Wang, X.; Wang, J. Profile distribution of CO2 in an arid saline-alkali soil with gypsum and wheat straw amendments: A two-year incubation experiment. Sci. Rep. 2018, 8, 11939. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, Y. Elucidating the molecular mechanisms mediating plant salt-stress responses. New Phytol. 2017, 217, 523–539. [Google Scholar] [CrossRef]
- Meena, M.D.; Joshi, P.K.; Narjary, B.; Sheoran, P.; Jat, H.S.; Chinchmalatpure, A.R.; Yadav, R.K.; Sharma, D.K. Effects of municipal solid waste compost, rice-straw compost and mineral fertilisers on biological and chemical properties of a saline soil and yields in a mustard-pearl millet cropping system. Soil Res. 2016, 54, 958–969. [Google Scholar] [CrossRef]
- Jesus, J.M.; Danko, A.S.; Fiúza, A.; Borges, M.-T. Phytoremediation of salt-affected soils: A review of processes, applicability, and the impact of climate change. Environ. Sci. Pollut. Res. 2015, 22, 6511–6525. [Google Scholar] [CrossRef]
- Luo, S.; Tian, L.; Chang, C.; Wang, S.; Zhang, J.; Zhou, X.; Li, X.; Tran, L.-S.P.; Tian, C. Grass and maize vegetation systems restore saline-sodic soils in the Songnen Plain of northeast China. Land Degrad. Dev. 2018, 29, 1107–1119. [Google Scholar] [CrossRef]
- Bandopadhyay, S.; Martin-Closas, L.; Pelacho, A.; Debruyn, J.M. Biodegradable Plastic Mulch Films: Impacts on Soil Microbial Communities and Ecosystem Functions. Front. Microbiol. 2018, 9, 819. [Google Scholar] [CrossRef] [PubMed]
- Steinmetz, Z.; Wollmann, C.; Schaefer, M.; Buchmann, C.; David, J.; Tröger, J.; Muñoz, K.; Frör, O.; Schaumann, G.E. Plastic mulching in agriculture. Trading short-term agronomic benefits for long-term soil degradation? Sci. Total Environ. 2016, 550, 690–705. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Ng, E.L.; Hu, W.; Wang, H.; Galaviz, P.; Yang, H.; Sun, W.; Li, C.; Ma, X.; Fu, B.; et al. Plastic pollution in croplands threatens long-term food security. Glob. Chang. Biol. 2020, 26, 3356–3367. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lv, S.; Zhang, M.; Chen, G.; Zhu, T.; Zhang, S.; Teng, Y.; Christie, P.; Luo, Y. Effects of plastic film residues on occurrence of phthalates and microbial activity in soils. Chemosphere 2016, 151, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Sun, Z.-X.; Feng, L.-S.; Zheng, M.-Z.; Chi, D.-C.; Meng, W.-Z.; Hou, Z.-Y.; Bai, W.; Li, K.-Y. Plastic Film Mulching for Water-Efficient Agricultural Applications and Degradable Films Materials Development Research. Mater. Manuf. Process. 2015, 30, 143–154. [Google Scholar] [CrossRef]
- Kader, M.A.; Senge, M.; Mojid, M.A.; Ito, K. Recent advances in mulching materials and methods for modifying soil environment. Soil Tillage Res. 2017, 168, 155–166. [Google Scholar] [CrossRef]
- Sun, D.; Li, H.; Wang, E.; He, W.; Hao, W.; Yan, C.; Li, Y.; Mei, X.; Zhang, Y.; Sun, Z.; et al. An overview of the use of plastic-film mulching in China to increase crop yield and water-use efficiency. Natl. Sci. Rev. 2020, 7, 1523–1526. [Google Scholar] [CrossRef]
- Kumar, M.; Etesami, H.; Kumar, V. (Eds.) Saline Soil-Based Agriculture by Halotolerant Microorganisms, 1st ed.; Springer Nature Singapore Pte Ltd.: Singapore, 2019. [Google Scholar]
- Bandopadhyay, S.; Sintim, H.Y.; Debruyn, J.M. Effects of biodegradable plastic film mulching on soil microbial communities in two agroecosystems. PeerJ 2020, 8, e9015. [Google Scholar] [CrossRef]
- Qian, H.; Zhang, M.; Liu, G.; Lu, T.; Qu, Q.; Du, B.; Pan, X. Effects of Soil Residual Plastic Film on Soil Microbial Community Structure and Fertility. Water Air Soil Pollut. 2018, 229, 261. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Qu, Q.; Zhang, Z.; Peijnenburg, W.J.G.M.; Liu, W.; Lu, T.; Hu, B.; Chen, J.-M.; Chen, J.; Lin, Z.; Qian, H. Rhizosphere Microbiome Assembly and Its Impact on Plant Growth. J. Agric. Food Chem. 2020, 68, 5024–5038. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.N.; Rastegari, A.A.; Yadav, N.; Kour, D. (Eds.) Advances in Plant Microbiome and Sustainable Agriculture, 1st ed.; Springer Nature Singapore Pte Ltd.: Singapore, 2020. [Google Scholar]
- Hartmann, A.; Rothballer, M.; Schmid, M. Lorenz Hiltner, a pioneer in rhizosphere microbial ecology and soil bacteriology research. Plant Soil 2007, 312, 7–14. [Google Scholar] [CrossRef]
- Fiorentino, N.; Ventorino, V.; Woo, S.L.; Pepe, O.; De Rosa, A.; Gioia, L.; Romano, I.; Lombardi, N.; Napolitano, M.; Colla, G.; et al. Trichoderma-Based Biostimulants Modulate Rhizosphere Microbial Populations and Improve N Uptake Efficiency, Yield, and Nutritional Quality of Leafy Vegetables. Front. Plant Sci. 2018, 9, 743. [Google Scholar] [CrossRef]
- Qiao, C.; Penton, C.R.; Xiong, W.; Liu, C.; Wang, R.; Liu, Z.; Xu, X.; Li, R.; Shen, Q. Reshaping the rhizosphere microbiome by bio-organic amendment to enhance crop yield in a maize-cabbage rotation system. Appl. Soil Ecol. 2019, 142, 136–146. [Google Scholar] [CrossRef]
- Kwak, M.-J.; Kong, H.G.; Choi, K.; Kwon, S.-K.; Song, J.Y.; Lee, J.; Lee, P.A.; Choi, S.Y.; Seo, M.; Lee, H.J.; et al. Rhizosphere microbiome structure alters to enable wilt resistance in tomato. Nat. Biotechnol. 2018, 36, 1100–1109. [Google Scholar] [CrossRef]
- Yuan, X.; Hong, S.; Xiong, W.; Raza, W.; Shen, Z.; Wang, B.; Li, R.; Ruan, Y.; Shen, Q.; Dini-Andreote, F. Development of fungal-mediated soil suppressiveness against Fusarium wilt disease via plant residue manipulation. Microbiome 2021, 9, 200. [Google Scholar] [CrossRef]
- Xu, L.; Dong, Z.; Chiniquy, D.; Pierroz, G.; Deng, S.; Gao, C.; Diamond, S.; Simmons, T.; Wipf, H.M.-L.; Caddell, D.; et al. Genome-resolved metagenomics reveals role of iron metabolism in drought-induced rhizosphere microbiome dynamics. Nat. Commun. 2021, 12, 3209. [Google Scholar] [CrossRef]
- Mukhtar, S.; Mehnaz, S.; Malik, K.A. Microbial diversity in the rhizosphere of plants growing under extreme environments and its impact on crop improvement. Environ. Sustain. 2019, 2, 329–338. [Google Scholar] [CrossRef]
- Bal, H.B.; Nayak, L.; Das, S.; Adhya, T.K. Isolation of ACC deaminase producing PGPR from rice rhizosphere and evaluating their plant growth promoting activity under salt stress. Plant Soil 2012, 366, 93–105. [Google Scholar] [CrossRef]
- Bharti, N.; Pandey, S.S.; Barnawal, D.; Patel, V.K.; Kalra, A. Plant growth promoting rhizobacteria Dietzia natronolimnaea modulates the expression of stress responsive genes providing protection of wheat from salinity stress. Sci. Rep. 2016, 6, 34768. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Brunel, C.; van Kleunen, M.; Li, J.; Jin, Z. Salinity-induced changes in the rhizosphere microbiome improve salt tolerance of Hibiscus hamabo. Plant Soil 2019, 443, 525–537. [Google Scholar] [CrossRef]
- Fu, L.; Penton, C.R.; Ruan, Y.; Shen, Z.; Xue, C.; Li, R.; Shen, Q. Inducing the rhizosphere microbiome by biofertilizer application to suppress banana Fusarium wilt disease. Soil Biol. Biochem. 2017, 104, 39–48. [Google Scholar] [CrossRef]
- Kinkel, L.L.; Bakker, M.G.; Schlatter, D.C. A Coevolutionary Framework for Managing Disease-Suppressive Soils. Annu. Rev. Phytopathol. 2011, 49, 47–67. [Google Scholar] [CrossRef]
- Mendes, R.; Kruijt, M.; de Bruijn, I.; Dekkers, E.; van der Voort, M.; Schneider, J.H.M.; Piceno, Y.M.; DeSantis, T.Z.; Andersen, G.L.; Bakker, P.A.H.M.; et al. Deciphering the Rhizosphere Microbiome for Disease-Suppressive Bacteria. Science 2011, 332, 1097–1100. [Google Scholar] [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Babalola, O.O. The rhizosphere microbial complex in plant health: A review of interaction dynamics. J. Integr. Agric. 2022, 21, 2168–2182. [Google Scholar] [CrossRef]
- Qiao, Q.; Wang, F.; Zhang, J.; Chen, Y.; Zhang, C.; Liu, G.; Zhang, H.; Ma, C.; Zhang, J. The Variation in the Rhizosphere Microbiome of Cotton with Soil Type, Genotype and Developmental Stage. Sci. Rep. 2017, 7, 3940. [Google Scholar] [CrossRef]
- Pii, Y.; Borruso, L.; Brusetti, L.; Crecchio, C.; Cesco, S.; Mimmo, T. The interaction between iron nutrition, plant species and soil type shapes the rhizosphere microbiome. Plant Physiol. Biochem. 2016, 99, 39–48. [Google Scholar] [CrossRef]
- Panpatte, D.G.; Jhala, Y.K. (Eds.) Soil Fertility Management for Sustainable Development, 1st ed.; Springer Nature Singapore Pte Ltd.: Singapore, 2019. [Google Scholar]
- Cucci, G.; Lacolla, G.; Mastro, M.A.; Caranfa, G. Leaching effect of rainfall on soil under four-year saline water irrigation. Soil Water Res. 2016, 11, 181–189. [Google Scholar] [CrossRef]
- Wang, W.; Gong, Y.; Xing, X. Groundwater Evaporation for Salt-Affected Soil Under Plastic Film-Covered Cultivation Condition: A Review. J. Soil Sci. Plant Nutr. 2020, 20, 1229–1237. [Google Scholar] [CrossRef]
- Hueso, S.; Hernández, T.; García, C. Resistance and resilience of the soil microbial biomass to severe drought in semiarid soils: The importance of organic amendments. Appl. Soil Ecol. 2011, 50, 27–36. [Google Scholar] [CrossRef]
- Khodadad, C.L.M.; Zimmerman, A.R.; Green, S.J.; Uthandi, S.; Foster, J.S. Taxa-specific changes in soil microbial community composition induced by pyrogenic carbon amendments. Soil Biol. Biochem. 2011, 43, 385–392. [Google Scholar] [CrossRef]
- Zhu, Y.; Chen, Y.; Gong, X.; Peng, Y.; Wang, Z.; Ji, B. Plastic film mulching improved rhizosphere microbes and yield of rainfed spring wheat. Agric. For. Meteorol. 2018, 263, 130–136. [Google Scholar] [CrossRef]
- Giri, D.D.; Kumar, A.; Shukla, P.N.; Singh, R.; Singh, P.K.; Pandey, K.D. Salt stress Tolerance of Methylotrophic Bacteria Methylophilus sp. and Methylobacterium sp. Isolated from Coal Mine Spoils. Pol. J. Microbiol. 2013, 62, 273–280. [Google Scholar] [CrossRef]
- Kalyaeva, M.A.; Zacharchenko, N.S.; Doronina, N.V.; Rukavtsova, E.B.; Ivanova, E.G.; Alekseeva, V.V.; Trotsenko, Y.A.; Bur’Yanov, Y.I. Plant Growth and Morphogenesis in vitro Is Promoted by Associative Methylotrophic Bacteria. Russ. J. Plant Physiol. 2001, 48, 514–517. [Google Scholar] [CrossRef]
- Pigoleva, S.V.; Zakharchenko, N.S.; Furs, O.V.; Tarlachkov, S.V.; Funtikova, T.V.; Filonov, A.E.; Aripovskii, A.V.; Dyachenko, O.V.; Buryanov, Y.I.; Shevchuk, T.V. Effects of Associative Microorganisms on Plant Growth and Resistance to Xenobiotics and Phytopathogens. Appl. Biochem. Microbiol. 2020, 56, 473–482. [Google Scholar] [CrossRef]
- Wang, W.; Liu, A.; Fu, W.; Peng, D.; Wang, G.; Ji, J.; Jin, C.; Guan, C. Tobacco-associated with Methylophilus sp. FP-6 enhances phytoremediation of benzophenone-3 through regulating soil microbial community, increasing photosynthetic capacity and maintaining redox homeostasis of plant. J. Hazard. Mater. 2022, 431, 128588. [Google Scholar] [CrossRef]
- Lv, H.; Masuda, S.; Fujitani, Y.; Sahin, N.; Tani, A. Oharaeibacter diazotrophicus gen. nov., sp. nov., a diazotrophic and facultatively methylotrophic bacterium, isolated from rice rhizosphere. Int. J. Syst. Evol. Microbiol. 2017, 67, 576–582. [Google Scholar] [CrossRef]
- Lv, H.; Tani, A. Genomic characterization of methylotrophy of Oharaeibacter diazotrophicus strain SM30T. J. Biosci. Bioeng. 2018, 126, 667–675. [Google Scholar] [CrossRef]
- Ham, S.; Yoon, H.; Park, J.-M.; Park, Y.G. Optimization of Fermentation Medium for Indole Acetic Acid Production by Pseudarthrobacter sp. NIBRBAC000502770. Appl. Biochem. Biotechnol. 2021, 193, 2567–2579. [Google Scholar] [CrossRef] [PubMed]
- Park, M.K.; Park, Y.J.; Kim, M.; Kim, M.C.; Ibal, J.C.; Kang, G.U.; Lee, G.D.; Tagele, S.B.; Kwon, H.J.; Kang, M.S.; et al. Complete genome sequence of a plant growth-promoting bacterium Pseudarthrobacter sp. NIBRBAC000502772, isolated from shooting range soil in the Republic of Korea. Korean J. Microbiol. 2020, 56, 390–393. [Google Scholar]
- Patel, S.; Gupta, R.S. A phylogenomic and comparative genomic framework for resolving the polyphyly of the genus Bacillus: Proposal for six new genera of Bacillus species, Peribacillus gen. nov., Cytobacillus gen. nov., Mesobacillus gen. nov., Neobacillus gen. nov., Metabacillus gen. nov. and Alkalihalobacillus gen. nov. Int. J. Syst. Evol. Microbiol. 2020, 70, 406–438. [Google Scholar] [CrossRef]
- Peng, J.; Ma, J.; Wei, X.; Zhang, C.; Jia, N.; Wang, X.; Wang, E.T.; Hu, D.; Wang, Z. Accumulation of beneficial bacteria in the rhizosphere of maize (Zea mays L.) grown in a saline soil in responding to a consortium of plant growth promoting rhizobacteria. Ann. Microbiol. 2021, 71, 40. [Google Scholar] [CrossRef]
- Jiang, L.; Lee, M.H.; Jeong, J.C.; Kim, D.H.; Kim, C.Y.; Kim, S.W.; Lee, J. Neobacillus endophyticus sp nov an endophytic bacterium isolated from Selaginella involvens roots. Int. J. Syst. Evol. Microbiol. 2021, 71, 004581. [Google Scholar] [CrossRef]
- Zhan, P.-C.; Li, C.-J.; Zhang, Z.; Mao, R.-F.; Liu, J.-R.; Jiang, X.-W.; Zhi, X.-Y.; Yang, L.-L. Neobacillus paridis sp. nov., an endophyte of Paris polyphylla Smith var. yunnanensis. Arch. Microbiol. 2022, 204, 129. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Hidalgo, P.; Flores-Félix, J.D.; Sánchez-Juanes, F.; Rivas, R.; Mateos, P.F.; Santa Regina, I.; Peix, Á.; Martínez-Molina, E.; Igual, J.M.; Velázquez, E. Identification of Canola Roots Endophytic Bacteria and Analysis of Their Potential as Biofertilizers for Canola Crops with Special Emphasis on Sporulating Bacteria. Agronomy 2021, 11, 1796. [Google Scholar] [CrossRef]
- Farokh, R.Z.; Sachdev, D.; Kazemi-Pour, N.; Engineer, A.; Pardesi, K.R.; Zinjarde, S.; Dhakephalkar, P.K.; Chopade, B.A. Characterization of Plant-Growth-Promoting Traits of Acinetobacter Species Isolated from Rhizosphere of Pennisetum glaucum. J. Microbiol. Biotechnol. 2011, 21, 556–566. [Google Scholar]
- Suckstorff, I.; Berg, G. Evidence for dose-dependent effects on plant growth by Stenotrophomonas strains from different origins. J. Appl. Microbiol. 2003, 95, 656–663. [Google Scholar] [CrossRef]
- Seckbach, J.; Grube, M. (Eds.) Symbioses and Stress, 1st ed.; Springer Science+Business Media: Dordrecht, The Netherlands, 2010. [Google Scholar]
- Kang, J.-P.; Huo, Y.; Kim, Y.-J.; Ahn, J.-C.; Hurh, J.; Yang, D.-U.; Yang, D.-C. Rhizobium panacihumi sp. nov., an isolate from ginseng-cultivated soil, as a potential plant growth promoting bacterium. Arch. Microbiol. 2018, 201, 99–105. [Google Scholar] [CrossRef]
- Chen, L.; He, L.Y.; Wang, Q.; Sheng, X.F. Synergistic effects of plant growth-promoting Neorhizobium huautlense T1-17 and immobilizers on the growth and heavy metal accumulation of edible tissues of hot pepper. J. Hazard. Mater. 2016, 312, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Chen, L.; He, L.-Y.; Sheng, X.-F. Increased biomass and reduced heavy metal accumulation of edible tissues of vegetable crops in the presence of plant growth-promoting Neorhizobium huautlense T1-17 and biochar. Agric. Ecosyst. Environ. 2016, 228, 9–18. [Google Scholar] [CrossRef]
- Österman, J.; Mousavi, S.A.; Koskinen, P.; Paulin, L.; Lindström, K. Genomic features separating ten strains of Neorhizobium galegae with different symbiotic phenotypes. BMC Genom. 2015, 16, 348. [Google Scholar] [CrossRef] [PubMed]
- Bonito, G.; Hameed, K.; Toome-Heller, M.; Healy, R.; Reid, C.; Liao, H.L.; Aime, M.C.; Schadt, C.; Vilgalys, R. Atractiella rhizophila, sp. nov., an endorrhizal fungus isolated from the Populus root microbiome. Mycologia 2017, 109, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Aoki, T.; O’Donnell, K.; Scandiani, M.M. Sudden death syndrome of soybean in South America is caused by four species of Fusarium: Fusarium brasiliense sp. nov., F. cuneirostrum sp. nov., F. tucumaniae, and F. virguliforme. Mycoscience 2005, 46, 162–183. [Google Scholar] [CrossRef]
- Deng, X.; Zhang, N.; Shen, Z.; Zhu, C.; Liu, H.; Xu, Z.; Li, R.; Shen, Q.; Salles, J.F. Soil microbiome manipulation triggers direct and possible indirect suppression against Ralstonia solanacearum and Fusarium oxysporum. npj Biofilms Microbiomes 2021, 7, 33. [Google Scholar] [CrossRef]
- Bitas, V.; McCartney, N.; Li, N.; Demers, J.; Kim, J.-E.; Kim, H.-S.; Brown, K.M.; Kang, S. Fusarium Oxysporum Volatiles Enhance Plant Growth Via Affecting Auxin Transport and Signaling. Front. Microbiol. 2015, 6, 1248. [Google Scholar] [CrossRef]
- Mulero-Aparicio, A.; Agustí-Brisach, C.; Varo, Á.; López-Escudero, F.J.; Trapero, A. A non-pathogenic strain of Fusarium oxysporum as a potential biocontrol agent against Verticillium wilt of olive. Biol. Control 2019, 139, 104045. [Google Scholar] [CrossRef]
- Sajeena, A.; Nair, D.S.; Sreepavan, K. Non-pathogenic Fusarium oxysporum as a biocontrol agent. Indian Phytopathol. 2020, 73, 177–183. [Google Scholar] [CrossRef]
- Tao, C.; Li, R.; Xiong, W.; Shen, Z.; Liu, S.; Wang, B.; Ruan, Y.; Geisen, S.; Shen, Q.; Kowalchuk, G.A. Bio-organic fertilizers stimulate indigenous soil Pseudomonas populations to enhance plant disease suppression. Microbiome 2020, 8, 137. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed]
- Niu, B.; Paulson, J.N.; Zheng, X.; Kolter, R. Simplified and representative bacterial community of maize roots. Proc. Natl. Acad. Sci. USA 2017, 114, E2450–E2459. [Google Scholar] [CrossRef]
- Han, Q.; Ma, Q.; Chen, Y.; Tian, B.; Xu, L.; Bai, Y.; Chen, W.; Li, X. Variation in rhizosphere microbial communities and its association with the symbiotic efficiency of rhizobia in soybean. ISME J. 2020, 14, 1915–1928. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 15 September 2021).
- Zhang, J.; Liu, Y.-X.; Zhang, N.; Hu, B.; Jin, T.; Xu, H.; Qin, Y.; Yan, P.; Zhang, X.; Guo, X.; et al. NRT1.1B is associated with root microbiota composition and nitrogen use in field-grown rice. Nat. Biotechnol. 2019, 37, 676–684. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Updating the 97% identity threshold for 16S ribosomal RNA OTUs. Bioinformatics 2018, 34, 2371–2375. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Liu, H.; Huang, X.; Tan, W.; Di, H.; Xu, J.; Li, Y. High manure load reduces bacterial diversity and network complexity in a paddy soil under crop rotations. Soil Ecol. Lett. 2020, 2, 104–119. [Google Scholar] [CrossRef]
- Lozupone, C.; Knight, R. UniFrac: A New Phylogenetic Method for Comparing Microbial Communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P. Vegan: Community Ecology Package Version 2.5-7. 2020. Available online: https://www.researchgate.net/publication/346579465 (accessed on 16 September 2021).
- Segata, N.; Izard, J.; Wald, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Chen, Y.; Neilson, J.W.; Kushwaha, P.; Maier, R.M.; Barberán, A. Life-history strategies of soil microbial communities in an arid ecosystem. ISME J. 2020, 15, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Jiao, S.; Chen, W.; Wang, J.; Du, N.; Li, Q.; Wei, G. Soil microbiomes with distinct assemblies through vertical soil profiles drive the cycling of multiple nutrients in reforested ecosystems. Microbiome 2018, 6, 146. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Guo, S.; Xiong, W.; Hang, X.; Gao, Z.; Jiao, Z.; Liu, H.; Mo, Y.; Zhang, N.; Kowalchuk, G.A.; Li, R.; et al. Protists as main indicators and determinants of plant performance. Microbiome 2021, 9, 64. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Eco. 2008, 26, 32–46. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OTU IDs | Genus | Spearman’s Rank Correlation | ||
|---|---|---|---|---|
| P | R | |||
| Potential key rhizosphere bacterial OTUs | OTU3058 | Methylovorus | 2.34 × 10−8 | 0.60 |
| OTU3280 | Oharaeibacter | 1.13 × 10−4 | 0.44 | |
| OTU2779 | Pseudarthrobacter | 3.47 × 10−5 | 0.47 | |
| OTU3480 | Neobacillus | 1.94 × 10−5 | 0.48 | |
| OTU369 | Peribacillus | 1.35 × 10−3 | 0.37 | |
| OTU113 | Acinetobacter | 2.29 × 10−3 | 0.35 | |
| OTU313 | Olivibacter | 7.34 × 10−3 | 0.31 | |
| OTU3424 | Stenotrophomonas | 2.30 × 10−2 | 0.27 | |
| OTU125 | Neorhizobium | 7.31 × 10−4 | 0.39 | |
| OTU935 | Dethiosulfatarculus | 2.91 × 10−2 | 0.26 | |
| OTU3336 | Methylophilus | 3.60 × 10−5 | 0.47 | |
| Potential key rhizosphere fungal OTUs | OTU1537 | Atractiellales_unidentified_1 | 1.01 × 10−6 | 0.54 |
| OTU113 | Pyrigemmula | 4.65 × 10−3 | 0.33 | |
| OTU1525 | Fusarium | 4.54 × 10−3 | 0.33 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, H.-C.; Sun, Y.; Tao, C.; Deng, X.; Xu, X.; Shen, Z.; Zhang, L.; Zheng, Z.; Huang, Y.; Hao, Y.; et al. Rhizosphere Microbiota Promotes the Growth of Soybeans in a Saline–Alkali Environment under Plastic Film Mulching. Plants 2023, 12, 1889. https://doi.org/10.3390/plants12091889
Mao H-C, Sun Y, Tao C, Deng X, Xu X, Shen Z, Zhang L, Zheng Z, Huang Y, Hao Y, et al. Rhizosphere Microbiota Promotes the Growth of Soybeans in a Saline–Alkali Environment under Plastic Film Mulching. Plants. 2023; 12(9):1889. https://doi.org/10.3390/plants12091889
Chicago/Turabian StyleMao, Han-Cheng, Yifei Sun, Chengyuan Tao, Xuhui Deng, Xu Xu, Zhenquan Shen, Laijie Zhang, Zehui Zheng, Yanhua Huang, Yongren Hao, and et al. 2023. "Rhizosphere Microbiota Promotes the Growth of Soybeans in a Saline–Alkali Environment under Plastic Film Mulching" Plants 12, no. 9: 1889. https://doi.org/10.3390/plants12091889
APA StyleMao, H.-C., Sun, Y., Tao, C., Deng, X., Xu, X., Shen, Z., Zhang, L., Zheng, Z., Huang, Y., Hao, Y., Zhou, G., Liu, S., Li, R., Guo, K., Tian, Z., & Shen, Q. (2023). Rhizosphere Microbiota Promotes the Growth of Soybeans in a Saline–Alkali Environment under Plastic Film Mulching. Plants, 12(9), 1889. https://doi.org/10.3390/plants12091889