
Effect of a Fortified Biostimulant Extract on Tomato Plant Productivity, Physiology, and Growing Media Properties

Abstract
:1. Introduction
2. Results
2.1. Plant Productivity
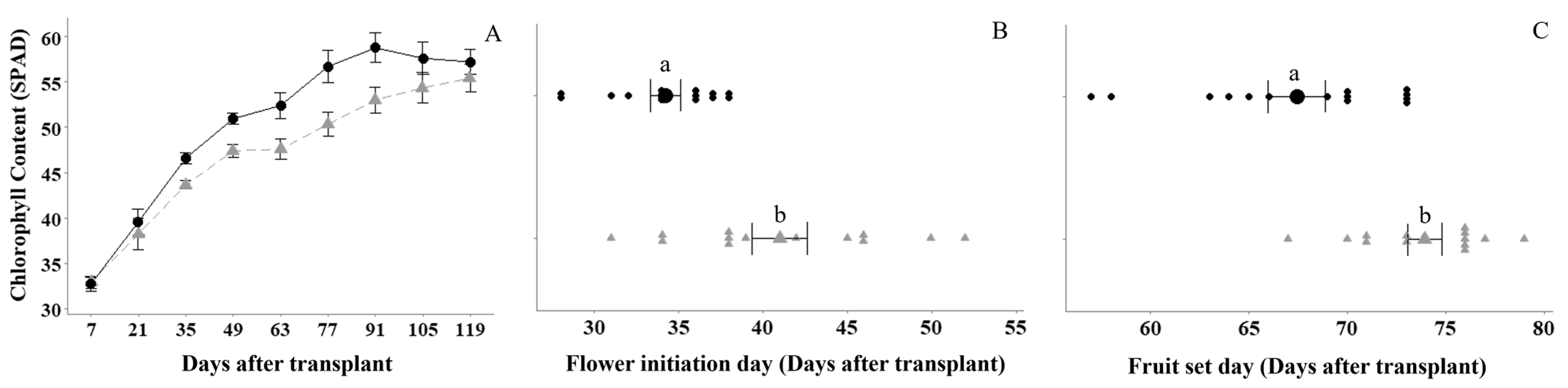
2.2. Plant Growth
2.3. Plant Nutrient Content
2.4. Available Nutrients in Growing Media
2.5. Microbiology in Growing Media
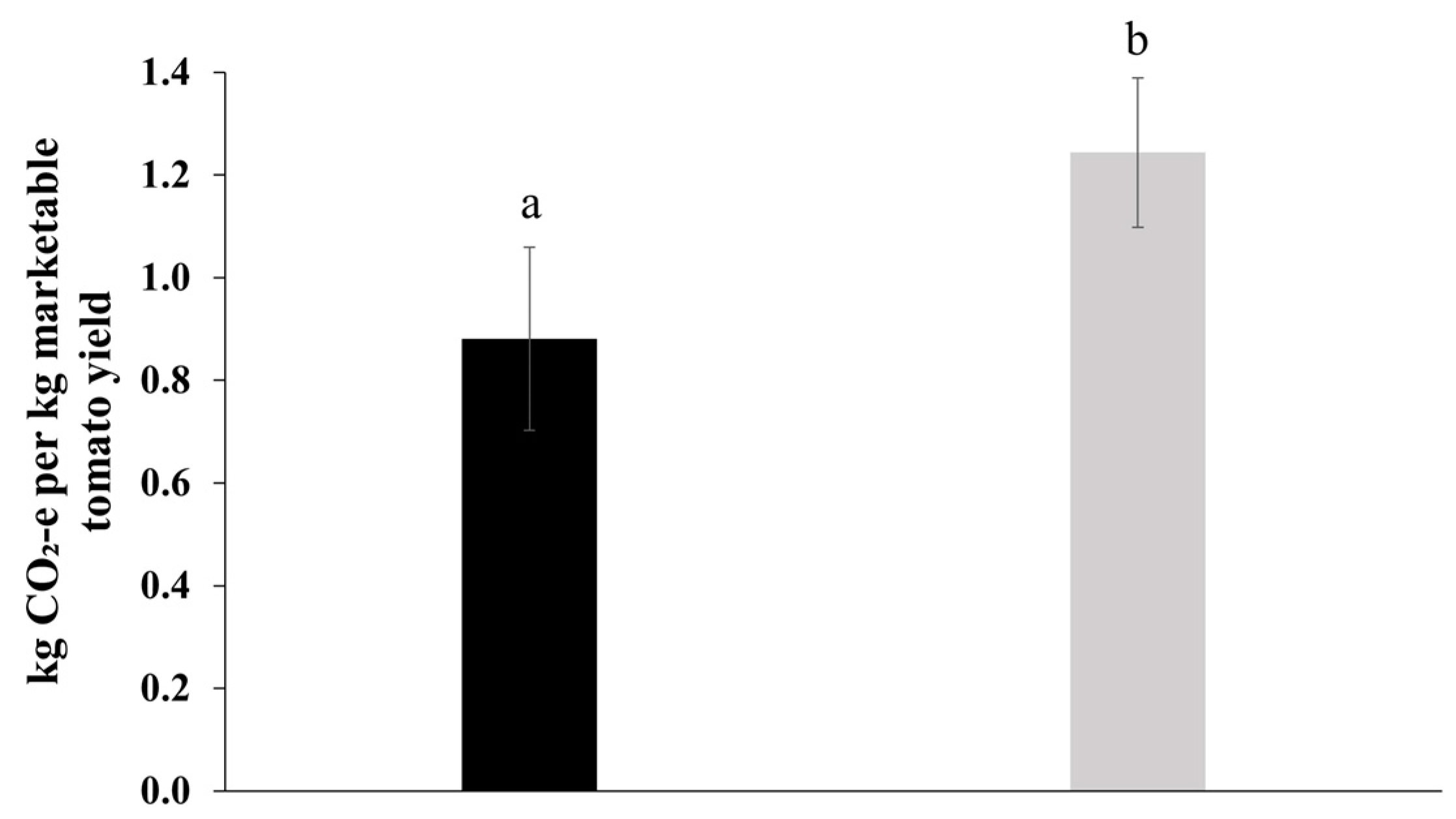
2.6. Greenhouse Gas Emissions
3. Discussion
3.1. The Effect of FBE on Tomato Plants
3.2. The Effect of FBE on Growing Media
3.3. The Effect of FBE on the Plant and Soil Ecosystem
4. Materials and Methods
4.1. Seedling Establishment
4.2. Experimental Plant Trials
4.3. Plant Growth and Productivity Assessments
4.4. Growing Medial Nutrient and Microbiology Assessments
4.5. Greenhouse Gas Emissions Assessment
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ray, D.K.; West, P.C.; Clark, M.; Gerber, J.S.; Prishchepov, A.V.; Chatterjee, S. Climate change has likely already affected global food production. PLoS ONE 2019, 14, e0217148. [Google Scholar] [CrossRef] [PubMed]
- Hughes, N. Analysis of Climate Change Impacts and Adaptation on Australian Farms; ABARES Insights; Australian Bureau of Agricultural and Resource Economics and Sciences (ABARES): Canberra, Australia, 2021. [Google Scholar] [CrossRef]
- Pedercini, M.; Züllich, G.; Dianati, K. Fertilizer Addiction: Implications for Sustainable Agriculture; Global Sustainable Development Report 2015 Brief. Available online: https://sustainabledevelopment.un.org/content/documents/592253-Pedercini-Fertilizer%20addiction.pdf (accessed on 20 October 2023).
- Mulvaney, R.; Khan, S.; Ellsworth, T. Synthetic nitrogen fertilizers deplete soil nitrogen: A global dilemma for sustainable cereal production. J. Environ. Qual. 2009, 38, 2295–2314. [Google Scholar] [CrossRef] [PubMed]
- Khangura, R.; Ferris, D.; Wagg, C.; Bowyer, J. Regenerative Agriculture—A Literature Review on the Practices and Mechanisms Used to Improve Soil Health. Sustainability 2023, 15, 2338. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Shukla, P.S.; Mantin, E.G.; Adil, M.; Bajpai, S.; Critchley, A.T.; Prithiviraj, B. Ascophyllum nodosum-based biostimulants: Sustainable applications in agriculture for the stimulation of plant growth, stress tolerance, and disease management. Front. Plant Sci. 2019, 10, 655. [Google Scholar] [CrossRef] [PubMed]
- Ali, O.; Ramsubhag, A.; Jayaraman, J. Biostimulant properties of seaweed extracts in plants: Implications towards sustainable crop production. Plants 2021, 10, 531. [Google Scholar] [CrossRef] [PubMed]
- Arioli, T.; Mattner, S.; Islam, M.; Tran, T.L.C.; Weisser, M.; Winberg, P.; Cahill, D. Applications of seaweed extracts in agriculture: An Australian perspective. J. Appl. Phycol. 2023, 1–14. [Google Scholar] [CrossRef]
- Deolu-Ajayi, A.O.; van der Meer, I.M.; Van der Werf, A.; Karlova, R. The power of seaweeds as plant biostimulants to boost crop production under abiotic stress. Plant Cell Environ. 2022, 45, 2537–2553. [Google Scholar] [CrossRef]
- Jindo, K.; Goron, T.L.; Pizarro-Tobías, P.; Sánchez-Monedero, M.Á.; Audette, Y.; Deolu-Ajayi, A.O.; van der Werf, A.; Goitom Teklu, M.; Shenker, M.; Pombo Sudré, C. Application of biostimulant products and biological control agents in sustainable viticulture: A review. Front. Plant Sci. 2022, 13, 3657. [Google Scholar] [CrossRef]
- Santaniello, A.; Scartazza, A.; Gresta, F.; Loreti, E.; Biasone, A.; Di Tommaso, D.; Piaggesi, A.; Perata, P. Ascophyllum nodosum seaweed extract alleviates drought stress in Arabidopsis by affecting photosynthetic performance and related gene expression. Front. Plant Sci. 2017, 8, 1362. [Google Scholar] [CrossRef]
- Cook, J.; Zhang, J.; Norrie, J.; Blal, B.; Cheng, Z. Seaweed extract (Stella Maris®) activates innate immune responses in Arabidopsis thaliana and protects host against bacterial pathogens. Mar. Drugs 2018, 16, 221. [Google Scholar] [CrossRef] [PubMed]
- Omidbakhshfard, M.A.; Sujeeth, N.; Gupta, S.; Omranian, N.; Guinan, K.J.; Brotman, Y.; Nikoloski, Z.; Fernie, A.R.; Mueller-Roeber, B.; Gechev, T.S. A biostimulant obtained from the seaweed Ascophyllum nodosum protects Arabidopsis thaliana from severe oxidative stress. Int. J. Mol. Sci. 2020, 21, 474. [Google Scholar] [CrossRef] [PubMed]
- Staykov, N.S.; Angelov, M.; Petrov, V.; Minkov, P.; Kanojia, A.; Guinan, K.J.; Alseekh, S.; Fernie, A.R.; Sujeeth, N.; Gechev, T.S. An Ascophyllum nodosum-derived biostimulant protects model and crop plants from oxidative stress. Metabolites 2020, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.T.; Arioli, T.; Cahill, D.M. Seaweed Extract-Stimulated Priming in Arabidopsis thaliana and Solanum lycopersicum. Plants 2021, 10, 2476. [Google Scholar] [CrossRef] [PubMed]
- Rasul, F.; Gupta, S.; Olas, J.J.; Gechev, T.; Sujeeth, N.; Mueller-Roeber, B. Priming with a seaweed extract strongly improves drought tolerance in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 1469. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.L.C.; Callahan, D.L.; Islam, M.T.; Wang, Y.; Arioli, T.; Cahill, D. Comparative metabolomic profiling of Arabidopsis thaliana roots and leaves reveals complex response mechanisms induced by a seaweed extract. Front. Plant Sci. 2023, 14, 699. [Google Scholar] [CrossRef] [PubMed]
- Fleming, T.R.; Fleming, C.C.; Levy, C.C.; Repiso, C.; Hennequart, F.; Nolasco, J.B.; Liu, F. Biostimulants enhance growth and drought tolerance in Arabidopsis thaliana and exhibit chemical priming action. Ann. Appl. Biol. 2019, 174, 153–165. [Google Scholar] [CrossRef]
- Islam, M.T.; Gan, H.M.; Ziemann, M.; Hussain, H.I.; Arioli, T.; Cahill, D. Phaeophyceaean (brown algal) extracts activate plant defense systems in Arabidopsis thaliana challenged with Phytophthora cinnamomi. Front. Plant Sci. 2020, 11, 852. [Google Scholar] [CrossRef]
- Madende, M.; Hayes, M. Fish by-product use as biostimulants: An overview of the current state of the art, including relevant legislation and regulations within the EU and USA. Molecules 2020, 25, 1122. [Google Scholar] [CrossRef]
- Meddich, A. Biostimulants for resilient agriculture—Improving plant tolerance to abiotic stress: A concise review. Gesunde Pflanz. 2023, 75, 709–727. [Google Scholar] [CrossRef]
- Canellas, L.P.; Olivares, F.L. Physiological responses to humic substances as plant growth promoter. Chem. Biol. Technol. Agric. 2014, 1, 3. [Google Scholar] [CrossRef]
- Liatile, P.C.; Potgieter, G.; Moloi, M.J. A Natural Bio-Stimulant Consisting of a Mixture of Fish Protein Hydrolysates and Kelp Extract Enhances the Physiological, Biochemical and Growth Responses of Spinach under Different Water Levels. Plants 2022, 11, 3374. [Google Scholar] [CrossRef] [PubMed]
- DPIRD. Emissions Baseline Report for the Agriculture Sector in Western Australia. Available online: https://www.agric.wa.gov.au/sites/gateway/files/2020%20Emissions%20Baseline%20Report%20for%20the%20Agriculture%20Sector%20in%20Western%20Australia.pdf (accessed on 30 October 2023).
- Ling, Q.; Huang, W.; Jarvis, P. Use of a SPAD-502 meter to measure leaf chlorophyll concentration in Arabidopsis thaliana. Photosynth. Res. 2011, 107, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Sim, C.; Zaharah, A.; Tan, M.; Goh, K. Rapid determination of leaf chlorophyll concentration, photosynthetic activity and NK concentration of Elaies guineensis via correlated SPAD-502 chlorophyll index. Asian J. Agric. Res. 2015, 9, 132–138. [Google Scholar]
- Mutale-Joan, C.; Redouane, B.; Najib, E.; Yassine, K.; Lyamlouli, K.; Laila, S.; Zeroual, Y.; Hicham, E.A. Screening of microalgae liquid extracts for their bio stimulant properties on plant growth, nutrient uptake and metabolite profile of Solanum lycopersicum L. Sci. Rep. 2020, 10, 2820. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Wang, X.; Chen, B.; Zhang, M.; Ma, J. Seaweed extract improved yields, leaf photosynthesis, ripening time, and net returns of tomato (Solanum lycopersicum Mill.). ACS Omega 2020, 5, 4242–4249. [Google Scholar] [CrossRef] [PubMed]
- Abetz, P.; Young, C. The effect of seaweed extract sprays derived from Ascophyllum nodosum on lettuce and cauliflower crops. Bot. Mar. 1983, 26, 487–492. [Google Scholar] [CrossRef]
- Crouch, I.; Van Staden, J. Effect of seaweed concentrate on the establishment and yield of greenhouse tomato plants. J. Appl. Phycol. 1992, 4, 291–296. [Google Scholar] [CrossRef]
- Yadav, A.N.; Kour, D.; Kaur, T.; Devi, R.; Yadav, N. Agriculturally important fungi for crop productivity: Current research and future challenges. In Agriculturally Important Fungi for Sustainable Agriculture: Volume 1: Perspective for Diversity and Crop Productivity; Springer: Cham, Switzerland, 2020; pp. 275–286. [Google Scholar] [CrossRef]
- Ding, W.; Cong, W.-F.; Lambers, H. Plant phosphorus-acquisition and-use strategies affect soil carbon cycling. Trends Ecol. Evol. 2021, 36, 899–906. [Google Scholar] [CrossRef]
- Beslemes, D.; Tigka, E.; Roussis, I.; Kakabouki, I.; Mavroeidis, A.; Vlachostergios, D. Effect of Arbuscular Mycorrhizal Fungi on Nitrogen and Phosphorus Uptake Efficiency and Crop Productivity of Two-Rowed Barley under Different Crop Production Systems. Plants 2023, 12, 1908. [Google Scholar] [CrossRef]
- Ma, M.; Taylor, P.W.; Chen, D.; Vaghefi, N.; He, J.-Z. Major soilborne pathogens of field processing tomatoes and management strategies. Microorganisms 2023, 11, 263. [Google Scholar] [CrossRef] [PubMed]
- Kuzyakov, Y.; Xu, X. Competition between roots and microorganisms for nitrogen: Mechanisms and ecological relevance. New Phytol. 2013, 198, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Henneron, L.; Kardol, P.; Wardle, D.A.; Cros, C.; Fontaine, S. Rhizosphere control of soil nitrogen cycling: A key component of plant economic strategies. New Phytol. 2020, 228, 1269–1282. [Google Scholar] [CrossRef] [PubMed]
- Hussain, H.I.; Kasinadhuni, N.; Arioli, T. The effect of seaweed extract on tomato plant growth, productivity and soil. J. Appl. Phycol. 2021, 33, 1305–1314. [Google Scholar] [CrossRef]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed extracts as biostimulants of plant growth and development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Jin, Q.; Zhang, Y.; Wang, Q.; Li, M.; Sun, H.; Liu, N.; Zhang, L.; Zhang, Y.; Liu, Z. Effects of potassium fulvic acid and potassium humate on microbial biodiversity in bulk soil and rhizosphere soil of Panax ginseng. Microbiol. Res. 2022, 254, 126914. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.; Zaidi, A.; Ahemad, M.; Oves, M.; Wani, P.A. Plant growth promotion by phosphate solubilizing fungi–current perspective. Arch. Agron. Soil Sci. 2010, 56, 73–98. [Google Scholar] [CrossRef]
- Colombo, C.; Palumbo, G.; Angelico, R.; Cho, H.G.; Francioso, O.; Ertani, A.; Nardi, S. Spontaneous aggregation of humic acid observed with AFM at different pH. Chemosphere 2015, 138, 821–828. [Google Scholar] [CrossRef]
- Sánchez-Sánchez, A.; Sánchez-Andreu, J.; Juárez, M.; Jordá, J.; Bermúdez, D. Improvement of iron uptake in table grape by addition of humic substances. J. Plant Nutr. 2006, 29, 259–272. [Google Scholar] [CrossRef]
- Çimrin, K.M.; Türkmen, Ö.; Turan, M.; Tuncer, B. Phosphorus and humic acid application alleviate salinity stress of pepper seedling. Afr. J. Biotechnol. 2010, 9, 5845–5851. [Google Scholar]
- El-Nemr, M.; El-Desuki, M.; El-Bassiony, A.; Fawzy, Z. Response of growth and yield of cucumber plants (Cucumis sativus L.) to different foliar applications of humic acid and bio-stimulators. Aust. J. Basic Appl. Sci. 2012, 6, 630–637. [Google Scholar]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Parađiković, N.; Vinković, T.; Vinković Vrček, I.; Žuntar, I.; Bojić, M.; Medić-Šarić, M. Effect of natural biostimulants on yield and nutritional quality: An example of sweet yellow pepper (Capsicum annuum L.) plants. J. Sci. Food Agric. 2011, 91, 2146–2152. [Google Scholar] [CrossRef] [PubMed]
- García-Martínez, A.; Díaz, A.; Tejada, M.; Bautista, J.; Rodríguez, B.; Santa María, C.; Revilla, E.; Parrado, J. Enzymatic production of an organic soil biostimulant from wheat-condensed distiller solubles: Effects on soil biochemistry and biodiversity. Process Biochem. 2010, 45, 1127–1133. [Google Scholar] [CrossRef]
- Schimel, J.P.; Bennett, J. Nitrogen mineralization: Challenges of a changing paradigm. Ecology 2004, 85, 591–602. [Google Scholar] [CrossRef]
- Spohn, M.; Kuzyakov, Y. Phosphorus mineralization can be driven by microbial need for carbon. Soil Biol. Biochem. 2013, 61, 69–75. [Google Scholar] [CrossRef]
- Horwath, W.R. The Role of the Soil Microbial Biomass in Cycling Nutrients; Tate, K.R., Ed.; World Scientific Publishing Europe Ltd.: London, UK, 2017; pp. 41–66. [Google Scholar]
- Hodge, A.; Fitter, A.H. Substantial nitrogen acquisition by arbuscular mycorrhizal fungi from organic material has implications for N cycling. Proc. Natl. Acad. Sci. USA 2010, 107, 13754–13759. [Google Scholar] [CrossRef]
- Paries, M.; Gutjahr, C. The good, the bad, and the phosphate: Regulation of beneficial and detrimental plant–microbe interactions by the plant phosphate status. New Phytol. 2023, 239, 29–46. [Google Scholar] [CrossRef]
- Kazan, K.; Lyons, R. The link between flowering time and stress tolerance. J. Exp. Bot. 2016, 67, 47–60. [Google Scholar] [CrossRef]
- Premalatha, K.; Botlagunta, N.; Santhosh, D.; Hiremath, C.; Verma, R.K.; Shanker, K.; Sundaresan, V.; Kalra, A. Enhancement of soil health, germination and crop productivity in Andrographis paniculata (Burm. f.) Nees, an important medicinal crop by using a composite bio inoculant. J. Plant Nutr. 2021, 44, 2331–2346. [Google Scholar]
- Lu, T.; Ke, M.; Lavoie, M.; Jin, Y.; Fan, X.; Zhang, Z.; Fu, Z.; Sun, L.; Gillings, M.; Peñuelas, J. Rhizosphere microorganisms can influence the timing of plant flowering. Microbiome 2018, 6, 231. [Google Scholar] [CrossRef] [PubMed]
- Novak, J.M.; Busscher, W.J.; Watts, D.W.; Amonette, J.E.; Ippolito, J.A.; Lima, I.M.; Gaskin, J.; Das, K.; Steiner, C.; Ahmedna, M. Biochars impact on soil-moisture storage in an ultisol and two aridisols. Soil Sci. 2012, 177, 310–320. [Google Scholar] [CrossRef]
- Maboko, M.; Du Plooy, C.; Chiloane, S. Effect of plant population, fruit and stem pruning on yield and quality of hydroponically grown tomato. Afr. J. Agric. Res. 2011, 6, 5144–5148. [Google Scholar]
- Prativa, K.; Bhattarai, B. Effect of integrated nutrient management on the growth, yield and soil nutrient status in tomato. Nepal J. Sci. Technol. 2011, 12, 23–28. [Google Scholar]
- Hussain, I.K.I.; Ahmed, M.; Khan, S.M.; Naveed, A.K.K.; Ali, S.; Hussain, I.; Sajid, M. Screening of different exotic lines of tomato (Lycopersicon esculentum L.) under the agro climatic condition of Haripur. Pure Appl. Biol. 2017, 6, 1251–1259. [Google Scholar]
- Yang, T.; Kim, H.-J. Nutrient management regime affects water quality, crop growth, and nitrogen use efficiency of aquaponic systems. Sci. Hortic. 2019, 256, 108619. [Google Scholar] [CrossRef]
- Minolta, K. Chlorophyll Meter SPAD-502Plus. A Lightweight Handheld Meter for Measuring the Chlorophyll Content of Leaves without Causing Damage to Plants. Available online: https://www.konicaminolta.com/instruments/download/catalog/color/pdf/spad502plus_catalog_eng.pdf (accessed on 24 November 2023).
- Huang, P.; de-Bashan, L.; Crocker, T.; Kloepper, J.W.; Bashan, Y. Evidence that fresh weight measurement is imprecise for reporting the effect of plant growth-promoting (rhizo) bacteria on growth promotion of crop plants. Biol. Fertil. Soils 2017, 53, 199–208. [Google Scholar] [CrossRef]
- Rayment, G.E.; Lyons, D.J. Soil chemical Methods: Australasia; Walker, J., Ed.; CSIRO Publishing: Colingwood, Australia, 2011; Volume 3. [Google Scholar]
- Rayment, G.; Peverill, K.; Shelley, B. Australian Soil and Plant Analysis Council soil and plant quality assurance programs and the impact of variability of test results on clients. Aust. J. Exp. Agric. 1998, 38, 777–784. [Google Scholar] [CrossRef]
- Madejón, P.; Murillo, J.M.; Marañón, T.; Cabrera, F.; Soriano, M. Trace element and nutrient accumulation in sunflower plants two years after the Aznalcóllar mine spill. Sci. Total Environ. 2003, 307, 239–257. [Google Scholar] [CrossRef]
- Schwarz, D.; Thompson, A.J.; Kläring, H.-P. Guidelines to use tomato in experiments with a controlled environment. Front. Plant Sci. 2014, 5, 625. [Google Scholar] [CrossRef]
- Mikhail, E.; Mikhail, T. Soil Microbiology. The Mikhail System: Understanding and Achieving Optimum Soil Balance, 2nd ed.; Edward H Mikhail: Endeavors Hill, Australia, 2017; pp. 135–138. [Google Scholar]
- Hillier, J.; Walter, C.; Malin, D.; Garcia-Suarez, T.; Mila-i-Canals, L.; Smith, P. A farm-focused calculator for emissions from crop and livestock production. Environ. Model. Softw. 2011, 26, 1070–1078. [Google Scholar] [CrossRef]
- Lam, W.Y.; Sim, S.; Kulak, M.; van Zelm, R.; Schipper, A.M.; Huijbregts, M.A. Drivers of variability in greenhouse gas footprints of crop production. J. Clean. Prod. 2021, 315, 128121. [Google Scholar] [CrossRef]
- Cool Soil Initiative. Available online: https://www.csu.edu.au/cool-soil-initiative (accessed on 30 October 2023).
- Porter, I.; Riches, D.; Scheer, C. Benchmarking and mitigation of nitrous oxide emissions from manures and fertilisers used in temperate vegetable crops in Australia. Soil Res. 2017, 55, 534–546. [Google Scholar] [CrossRef]
- Riches, D.; Mattner, S.; Davies, R.; Porter, I. Mitigation of nitrous oxide emissions with nitrification inhibitors in temperate vegetable cropping in southern Australia. Soil Res. 2016, 54, 533–543. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Nutrient (mg Plant−1) | FBE Treatment | Untreated Control | Ratio | 95% CI | p-Value |
|---|---|---|---|---|---|
| Total N | 384 a | 259 b | 1.48 | (1.27, 1.74) | <0.001 |
| Total P | 84.6 a | 67.5 b | 1.25 | (1.07, 1.47) | 0.010 |
| Total K | 660 a | 504 b | 1.31 | (1.10, 1.49) | 0.006 |
| Total S | 168 a | 126 b | 1.34 | (1.04, 1.72) | 0.029 |
| Total Ca | 283 a | 219 b | 1.29 | (1.07, 1.57) | 0.014 |
| Total Mg | 91.7 a | 68.8 b | 1.33 | (1.11, 1.60) | 0.006 |
| Total Fe | 2.48 a | 1.61 b | 1.54 | (1.17, 2.05) | 0.006 |
| Total Mn | 1.41 a | 0.97 b | 1.45 | (1.17, 1.80) | 0.003 |
| Total Zn | 1.29 a | 1.07 a | 1.20 | (0.87, 1.66) | 0.237 |
| Total Cu | 0.39 a | 0.29 b | 1.34 | (1.06, 1.71) | 0.021 |
| Total Co | 0.0019 a | 0.0013 b | 1.50 | (1.19, 1.89) | 0.003 |
| Total Mo | 0.0026 a | 0.0021 a | 1.26 | (0.92, 1.72) | 0.141 |
| Total B | 0.49 a | 0.4 b | 1.22 | (1.01, 1.47) | 0.042 |
| Nutrients in Growing Media (ppm g−1) | FBE Treatment | Untreated Control | Diff. | 95% CI | p-Value |
|---|---|---|---|---|---|
| Available N | 90.1 a | 48.8 b | 41.3 | (8.6, 74.0) | 0.017 |
| Available P | 213 a | 180 b | 33 | (10, 56) | 0.009 |
| Available K | 1475 a | 1421 a | 54 | (−85, 192) | 0.420 |
| Available S | 860 a | 694 a | 167 | (35, 368) | 0.097 |
| Available Ca | 6424 a | 6259 a | 166 | (−352, 684) | 0.500 |
| Available Mg | 1245 a | 1288 a | −43 | (−150, 64) | 0.400 |
| Available Fe | 33.5 a | 25.5 b | 8.0 | (0.0, 16.0) | 0.050 |
| Available Mn | 22.4 a | 22.1 a | 0.2 | (−4.4, 4.8) | 0.920 |
| Available Zn | 13.4 a | 13.9 a | −0.5 | (−1.7, 0.6) | 0.330 |
| Available Cu | 17 a | 18.7 b | −1.7 | (−3.2, −0.1) | 0.039 |
| Available Co | 0.38 a | 0.45 a | 0.07 | (−0.19, 0.05) | 0.230 |
| Available B | 0.20 a | 0.15 a | 0.05 | (−0.00, 0.10) | 0.075 |
| Available Mo | 1.53 a | 1.46 a | 0.07 | (−0.17, 0.31) | 0.550 |
| Media Microbiology (cfu g−1) | FBE Treatment | Untreated Control | Ratio | 95% CI | p-Value |
|---|---|---|---|---|---|
| Total active population | 2951 a | 1905 b | 1.55 | (1.23, 1.95) | 0.001 |
| Total active fungi | 1950 a | 1132 b | 1.72 | (1.31, 2.27) | <0.001 |
| Total active yeasts | 0.54 a | 0.39 a | 1.38 | (0.64, 3.01) | 0.380 |
| Total active bacteria | 762 a | 675 a | 1.13 | (0.72, 1.76) | 0.560 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weisser, M.; Mattner, S.W.; Southam-Rogers, L.; Hepworth, G.; Arioli, T. Effect of a Fortified Biostimulant Extract on Tomato Plant Productivity, Physiology, and Growing Media Properties. Plants 2024, 13, 4. https://doi.org/10.3390/plants13010004
Weisser M, Mattner SW, Southam-Rogers L, Hepworth G, Arioli T. Effect of a Fortified Biostimulant Extract on Tomato Plant Productivity, Physiology, and Growing Media Properties. Plants. 2024; 13(1):4. https://doi.org/10.3390/plants13010004
Chicago/Turabian StyleWeisser, Marianne, Scott William Mattner, Liam Southam-Rogers, Graham Hepworth, and Tony Arioli. 2024. "Effect of a Fortified Biostimulant Extract on Tomato Plant Productivity, Physiology, and Growing Media Properties" Plants 13, no. 1: 4. https://doi.org/10.3390/plants13010004
APA StyleWeisser, M., Mattner, S. W., Southam-Rogers, L., Hepworth, G., & Arioli, T. (2024). Effect of a Fortified Biostimulant Extract on Tomato Plant Productivity, Physiology, and Growing Media Properties. Plants, 13(1), 4. https://doi.org/10.3390/plants13010004