
A New Method for Discovering Plant Biostimulants



Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Fermented Corn Straw
2.2. Insects and Feeding Conditions
2.3. Preparation of Samples
2.4. Isolation and Structural Characterization of HA in Samples
2.5. Identification of Organic Compounds in Digestive Tract Contents
2.6. Determination of the Seed Germination Index of Intestinal Compounds
2.7. Screening the Concentration of Plant Growth-Promoting Intestinal Compounds
2.8. Validation of the Plant Growth-Promoting Activity of Intestinal Compounds
2.9. Statistical Analysis
2.10. Method Flow Chart
3. Results
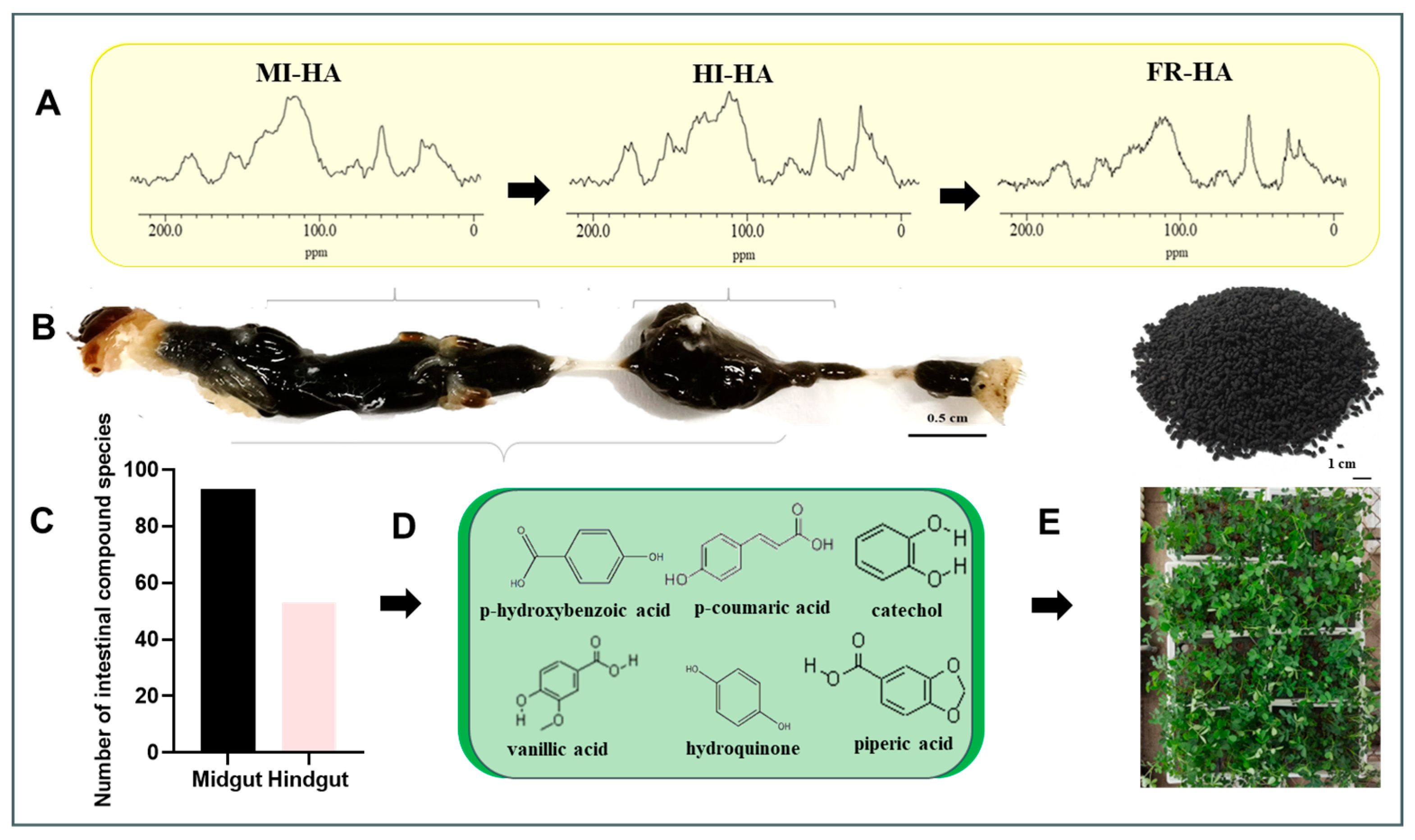
3.1. Analysis of HA Structure in the Gut of PBLs
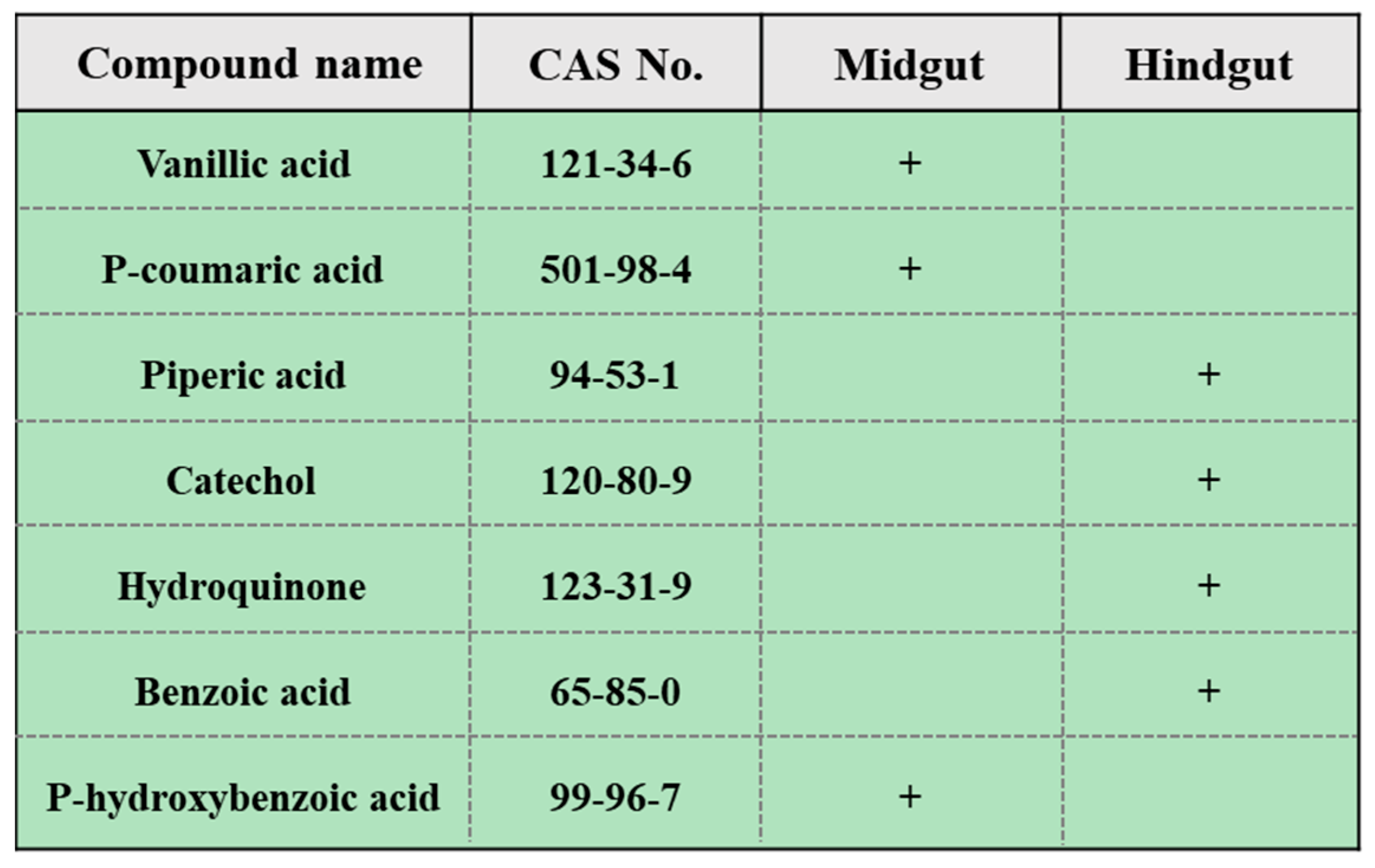
3.2. Organic Compounds in Gut Contents
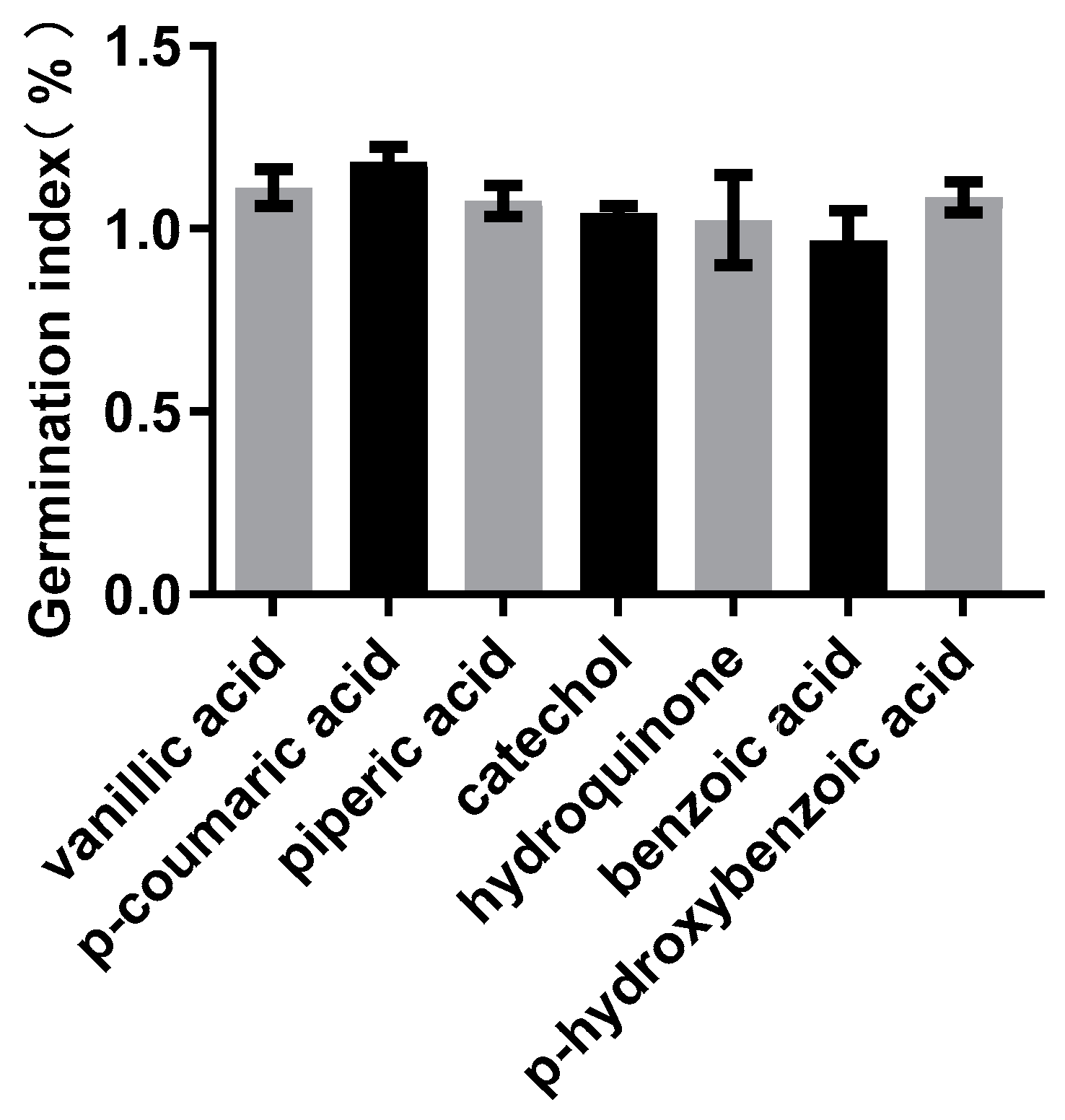
3.3. Determination of the Seed GI of the Selected Compounds
3.4. Screening of the Plant Growth-Promoting Concentration of these Compounds
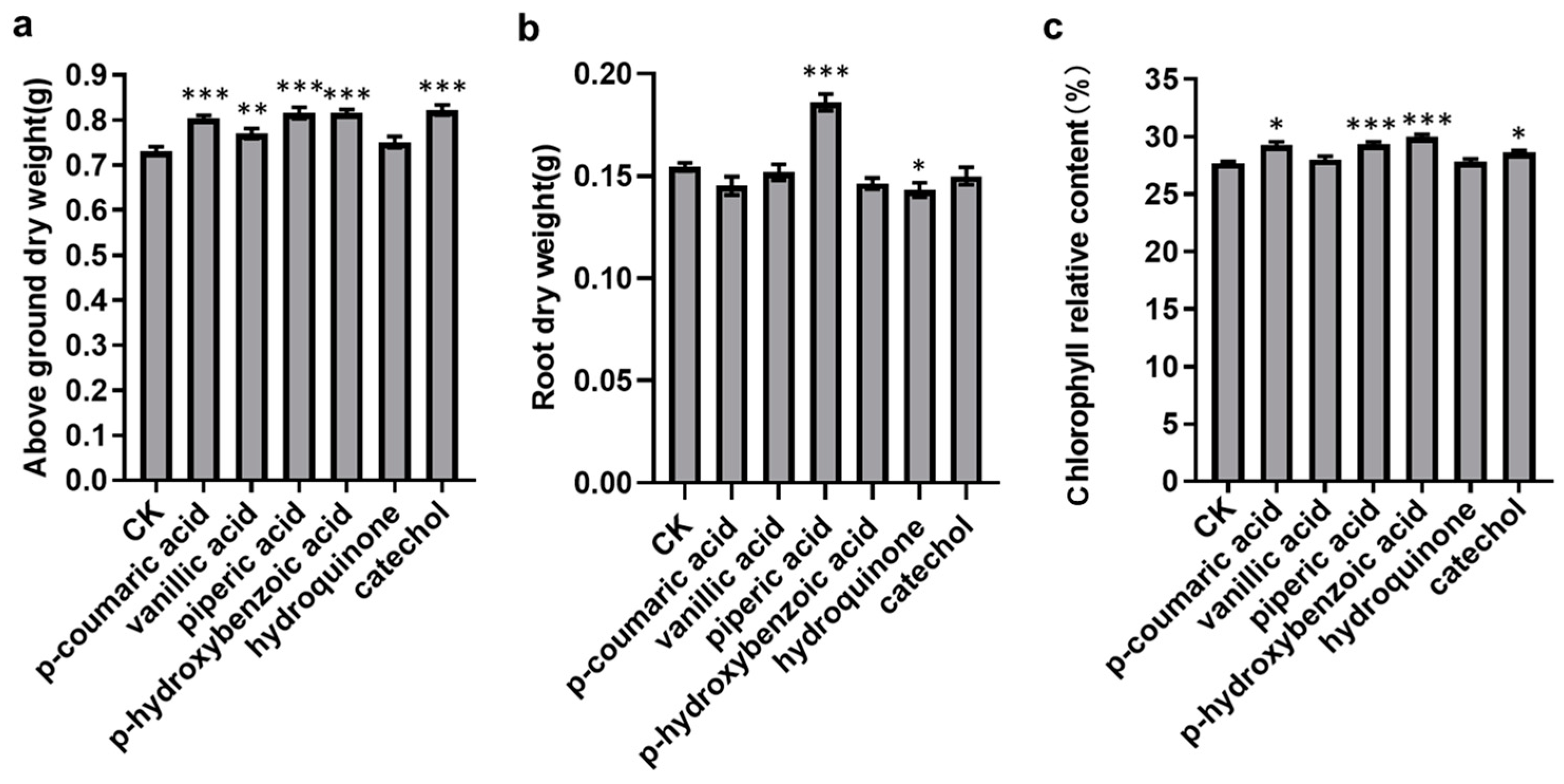
3.5. Validation of the Plant Growth-Promoting Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Madende, M.; Hayes, M. Fish By-Product Use as Biostimulants: An Overview of the Current State of the Art, Including Relevant Legislation and Regulations within the EU and USA. Molecules 2020, 25, 1122. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [PubMed]
- Pizzeghello, D.; Schiavon, M.; Francioso, O.; Dalla Vecchia, F.; Ertani, A.; Nardi, S. Bioactivity of Size-Fractionated and Unfractionated Humic Substances From Two Forest Soils and Comparative Effects on N and S Metabolism, Nutrition, and Root Anatomy of Allium sativum L. Front. Plant Sci. 2020, 11, 1203. [Google Scholar] [CrossRef]
- Keiji, J.; Lopes, O.F.; Paixão, M.D.J.d.; Angel, S.-M.M.; Corné, K.; Pasqualoto, C.L. From Lab to Field: Role of Humic Substances Under Open-Field and Greenhouse Conditions as Biostimulant and Biocontrol Agent. Front. Plant Sci. 2020, 11, 426. [Google Scholar]
- Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Jones, D.L.; Nebbioso, A.; Mazzei, P.; Piccolo, A. Humic and fulvic acids as biostimulants in horticulture. Sci. Hortic. 2015, 196, 15–27. [Google Scholar] [CrossRef]
- Wnuk, E.; Wasko, A.; Walkiewicz, A.; Bartminski, P.; Bejger, R.; Mielnik, L.; Bieganowski, A. The effects of humic substances on DNA isolation from soils. PeerJ 2020, 8, e9378. [Google Scholar] [CrossRef]
- Li, Y.; Fang, F.; Wei, J.; Wu, X.; Cui, R.; Li, G.; Zheng, F.; Tan, D. Humic Acid Fertilizer Improved Soil Properties and Soil Microbial Diversity of Continuous Cropping Peanut: A Three-Year Experiment. Sci. Rep. 2019, 9, 12014. [Google Scholar] [CrossRef]
- Ren, H.; Islam, M.S.; Wang, H.; Guo, H.; Wang, Z.; Qi, X.; Zhang, S.; Guo, J.; Wang, Q.; Li, B. Effect of Humic Acid on Soil Physical and Chemical Properties, Microbial Community Structure, and Metabolites of Decline Diseased Bayberry. Int. J. Mol. Sci. 2022, 23, 4707. [Google Scholar] [CrossRef]
- Johannes, L.; Markus, K. The contentious nature of soil organic matter. Nature 2015, 528, 60–68. [Google Scholar]
- Li, Y.; Fu, T.; Geng, L.; Shi, Y.; Chu, H.; Liu, F.; Liu, C.; Song, F.; Zhang, J.; Shu, C. Protaetia brevitarsis larvae can efficiently convert herbaceous and ligneous plant residues to humic acids. Waste Manag. 2019, 83, 79–82. [Google Scholar] [CrossRef]
- Dores-Silva, P.R.; Landgraf, M.D.; Rezende, M.O.O. Humification process in different kinds of organic residue by composting and vermicomposting: Have microbioreactors really accelerated the process? Environ. Sci. Pollut. Res. Int. 2018, 25, 17490–17498. [Google Scholar] [CrossRef] [PubMed]
- Frouz, J. Effects of soil macro- and mesofauna on litter decomposition and soil organic matter stabilization. Geoderma 2018, 332, 161–172. [Google Scholar] [CrossRef]
- Du, B.H.; Xuan, H.N.; Geng, L.L.; Li, W.H.; Zhang, J.; Xiang, W.S.; Liu, R.M.; Shu, C.L. Microflora for improving the Auricularia auricula spent mushroom substrate for Protaetia brevitarsis production. iScience 2022, 25, 105307. [Google Scholar] [CrossRef] [PubMed]
- Wei, P.; Li, Y.; Lai, D.; Geng, L.; Liu, C.; Zhang, J.; Shu, C.; Liu, R. Protaetia brevitarsis larvae can feed on and convert spent mushroom substrate from Auricularia auricula and Lentinula edodes cultivation. Waste Manag. 2020, 114, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Xuan, H.; Gao, P.; Du, B.; Geng, L.; Wang, K.; Huang, K.; Zhang, J.; Huang, T.; Shu, C. Characterization of Microorganisms from Protaetia brevitarsis Larva Frass. Microorganisms 2022, 10, 311. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Gao, P.W.; Geng, L.L.; Liu, C.Q.; Zhang, J.; Shu, C.L. Lignocellulose degradation in Protaetia brevitarsis larvae digestive tract: Refining on a tightly designed microbial fermentation production line. Microbiome 2022, 10, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Li, P.; Gao, Y.; Liu, C.; Shu, C. De novo genome assembly of the white-spotted flower chafer (Protaetia brevitarsis). GigaScience 2019, 8, giz019. [Google Scholar] [CrossRef]
- Tian, F.Y.; Liu, G.F.; Wu, J.X. Exploration on metabolic characteritics of Pengbo semi-wool sheep under the condition of grazing and barn feeding based on non-targeted metabonomics. Chin. J. Vet. Sci. 2020, 40, 1854–1863. [Google Scholar]
- Parvin, K.; Nahar, K.; Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Mohsin, S.M.; Fujita, M. Exogenous vanillic acid enhances salt tolerance of tomato: Insight into plant antioxidant defense and glyoxalase systems. Plant Physiol. Biochem. 2020, 150, 109–120. [Google Scholar] [CrossRef]
- Morriën, E.; Prescott, C.E. Pellets or particles? How can we predict the effect of soil macro-arthropods on litter decomposition? Funct. Ecol. 2018, 32, 2480–2482. [Google Scholar] [CrossRef]
- Joly, F.X.; Coq, S.; Coulis, M.; Nahmani, J.; Hättenschwiler, S. Litter conversion into detritivore faeces reshuffles the quality control over C and N dynamics during decomposition. Funct. Ecol. 2018, 32, 2605–2614. [Google Scholar] [CrossRef]
- Lai, D.Q.; Wang, Q.L.; Wu, Y.; Shu, C.L.; Zhang, Y.; Liu, C.Q. Effect of Protaetia brevitarsis (Lewis) larvae larvae dung on development of pepper seedling stage under low temperature. North. Hortic. 2019, 8, 63–66. [Google Scholar]
- Guo, X.X.; Liu, H.T.; Wu, S.B. Humic substances developed during organic waste composting: Formation mechanisms, structural properties, and agronomic functions. Sci. Total Environ. 2019, 662, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhao, Y.; Wang, R.; Lu, Q.; Wu, J.; Zhang, D.; Nie, Z.; Wei, Z. Effect of the addition of exogenous precursors on humic substance formation during composting. Waste Manag. 2018, 79, 462–471. [Google Scholar] [CrossRef]
- Chen, Y.A.; Zhou, Y.; Qin, Y.; Liu, D.; Zhao, X. Evaluation of the action of Tween 20 non-ionic surfactant during enzymatic hydrolysis of lignocellulose: Pretreatment, hydrolysis conditions and lignin structure. Bioresour. Technol. 2018, 269, 329–338. [Google Scholar] [CrossRef]
- Wang, M.; Schoettner, M.; Xu, S.Q.; Paetz, C.; Wilde, J.; Baldwin, I.T.; Groten, K. Catechol, a major component of smoke, influences primary root growth and root hair elongation through reactive oxygen species-mediated redox signaling. New Phytol. 2017, 213, 1755–1770. [Google Scholar] [CrossRef]
- Retig, N.; Chet, I. Catechol-induced resistance of tomato plants to Fusarium wilt. Physiol. Plant Pathol. 1974, 4, 469–472. [Google Scholar] [CrossRef]
- Nkomo, M.; Gokul, A.; Keyster, M.; Klein, A. Exogenous p-Coumaric Acid Improves Salvia hispanica L. Seedling Shoot Growth. Plants 2019, 8, 546. [Google Scholar] [CrossRef]
- Elwakil, M.A. Use of Antioxidant Hydroquinone in the Control of Seed-borne Fungi of Peanut with Special Reference to the Production of Good Quality Seed. Plant Pathol. J. 2003, 2, 75–79. [Google Scholar] [CrossRef]
- Muhammad, K.; Khan, A.L.; Liaqat, A.; Javid, H.; Muhammad, W.; Ahmed, A.H.; Imran, Q.M.; Yoon-Ha, K.; Sang-Mo, K.; Byung-Wook, Y. Hydroquinone; A Novel Bioactive Compound from Plant-Derived Smoke Can Cue Seed Germination of Lettuce. Front. Chem. 2017, 5, 30. [Google Scholar]
- Li, M.F.; Li, W.; Du, W.; Li, J.J. Dormancy Mechanism and Bioactivity of Hydroquinone Extracted from Seed of Podophyllum hexandrum Royle. Bull. Bot. Res. 2010, 30, 215–220. [Google Scholar]
- Shen, L.F.; Xu, Y.Y.; Huang, X.L.; Yang, M. Effects of p-Hydroxybenzoic Acid and Cinnamic Acid on the Growth and the Physiological Indexes of Seedlings of Eucalyp-tus grandis × E. urophylla No.9. J. Southwest For. Univ. (Nat. Sci.) 2020, 40, 19–26. [Google Scholar]
- Gao, B.; Jin, X.P.; Chen, Y. Effect of P-hydroxybenzoic Acid on Seedling Growth of Isatis indigotica. Gansu Agric. Sci. Technol. 2020, 57–61. [Google Scholar]
- Zarai, Z.; Boujelbene, E.; Salem, N.B.; Gargouri, Y.; Sayari, A. Antioxidant and antimicrobial activities of various solvent extracts, piperine and piperic acid from Piper nigrum. LWT-Food Sci. Technol. 2013, 50, 634–641. [Google Scholar] [CrossRef]
- Singh, G.; Wani, N.A.; Rahim, J.U.; Shankar, S.; Rai, R.; Katoch, M. Synergistic antimicrobial and antibiofilm activities of piperic acid and 4-ethylpiperic acid amides in combination with ciprofloxacin. J. Antibiot. 2022, 75, 236–242. [Google Scholar] [CrossRef]
- Araujo, O.P.D.; Brito, D.A.T.; Guerra, D.O.R.; Mastrangelo, G.A.G.; Mattos, D.O.J.; Benedito, A.D.S.B.; Roberto, L.M.; Silva, C.R.W.D.; Nascimento, F.A.T.D.; Almeida, V.M.L.A.D. Evaluation of the antinociceptive and anti-inflammatory activities of piperic acid: Involvement of the cholinergic and vanilloid systems. Eur. J. Pharmacol. 2018, 834, 54–64. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| % Alkyl C (0–44 ppm) | % Methoxy C (44–64 ppm) | % Alkoxy C (64–93 ppm) | % Aromatic C (93–142 ppm) | % Carboloy C (142–162 ppm) | % Carboxyl C (162–188 ppm) | |
|---|---|---|---|---|---|---|
| MI-HA | 15.08 | 10.09 | 5.21 | 57.62 | 5.97 | 6.04 |
| HI-HA | 17.77 | 8.36 | 5.11 | 50.21 | 11.03 | 7.52 |
| FR-HA | 19.42 | 10.40 | 2.96 | 53.03 | 7.44 | 6.77 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, P.; Wang, K.; Qi, C.; Chen, K.; Xiang, W.; Zhang, Y.; Zhang, J.; Shu, C. A New Method for Discovering Plant Biostimulants. Plants 2024, 13, 56. https://doi.org/10.3390/plants13010056
Gao P, Wang K, Qi C, Chen K, Xiang W, Zhang Y, Zhang J, Shu C. A New Method for Discovering Plant Biostimulants. Plants. 2024; 13(1):56. https://doi.org/10.3390/plants13010056
Chicago/Turabian StyleGao, Peiwen, Kui Wang, Chang Qi, Keming Chen, Wensheng Xiang, Yue Zhang, Jie Zhang, and Changlong Shu. 2024. "A New Method for Discovering Plant Biostimulants" Plants 13, no. 1: 56. https://doi.org/10.3390/plants13010056
APA StyleGao, P., Wang, K., Qi, C., Chen, K., Xiang, W., Zhang, Y., Zhang, J., & Shu, C. (2024). A New Method for Discovering Plant Biostimulants. Plants, 13(1), 56. https://doi.org/10.3390/plants13010056