

Codium fragile (Suringar) Hariot as Biostimulant Agent to Alleviate Salt Stress in Durum Wheat: Preliminary Results from Germination Trials





Abstract
:1. Introduction
2. Materials and Methods
2.1. Algal Collection and Extract Preparation
2.2. Experimental Design
- A liquid fungicide, usually used for cereals coating in Europe, at a dose of 200 g for 100 kg of seed, containing 2.34% Fludioxonil, 2.34% Difenoconazole, and 0.93% Tebuconazole;
- Codium fragile extract at 10 g dry matter (gdm) L−1;
- Distilled water.
2.3. Measured Traits
- Final germination percentage: (seeds germinated/total seeds) × 100, after 8 days;
- Germination time course.
- Number of roots, manually counted;
- Length of the main root (cm), measured with a ruler;
- Coleoptile length (cm), measured with a ruler;
- Root biomass (dry weight, g), for which all the roots of each replicate were separated from the germinated seeds and dried off for 2 days at 60 °C;
- Coleoptile biomass (dry weight, g), for which all the coleoptiles of each replicate were separated from the germinated seeds and dried off for 2 days at 60 °C.
2.4. Statistical Analysis
3. Results
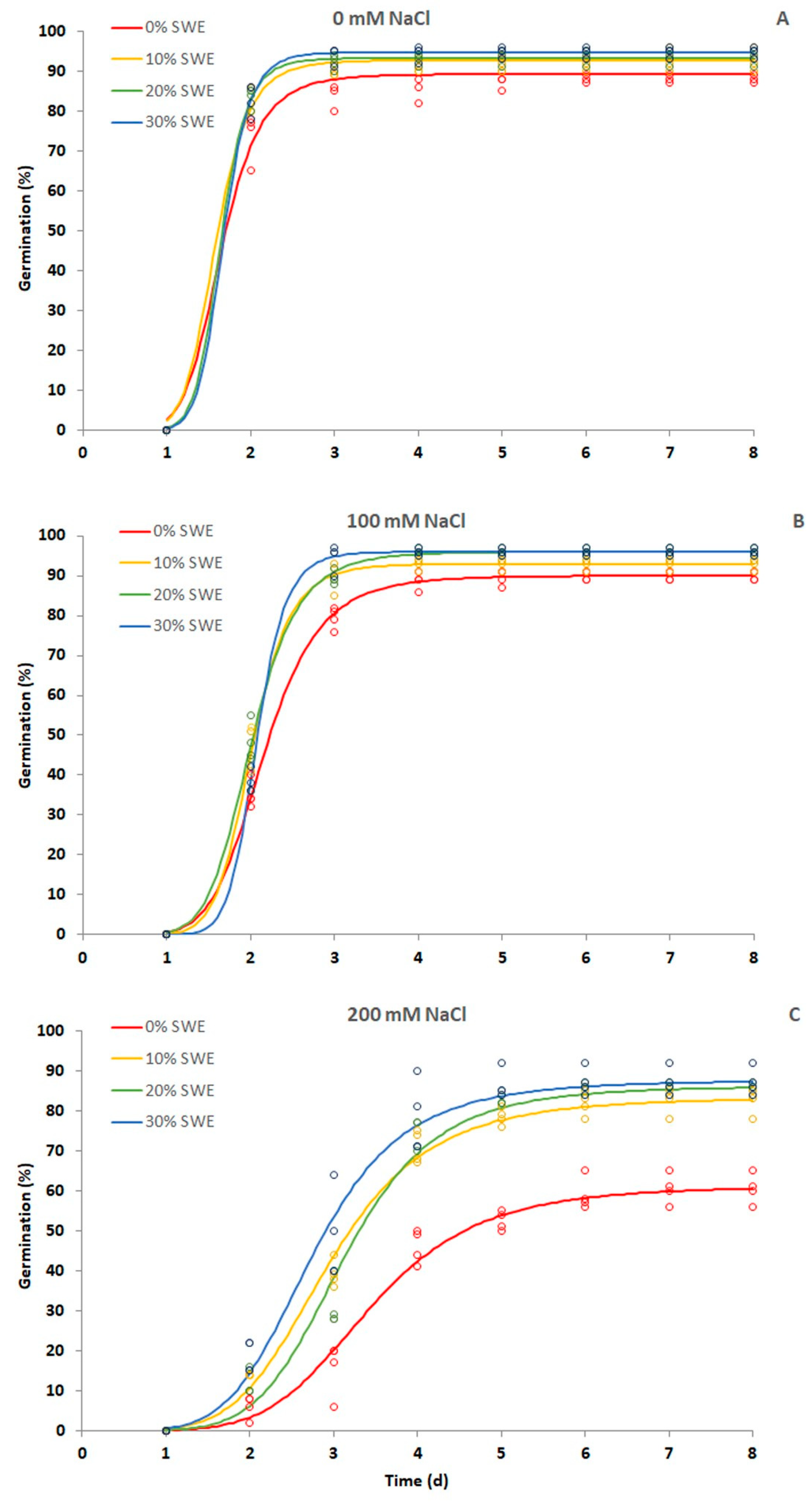
3.1. Germination Percentage and Median Germination Time (T50)
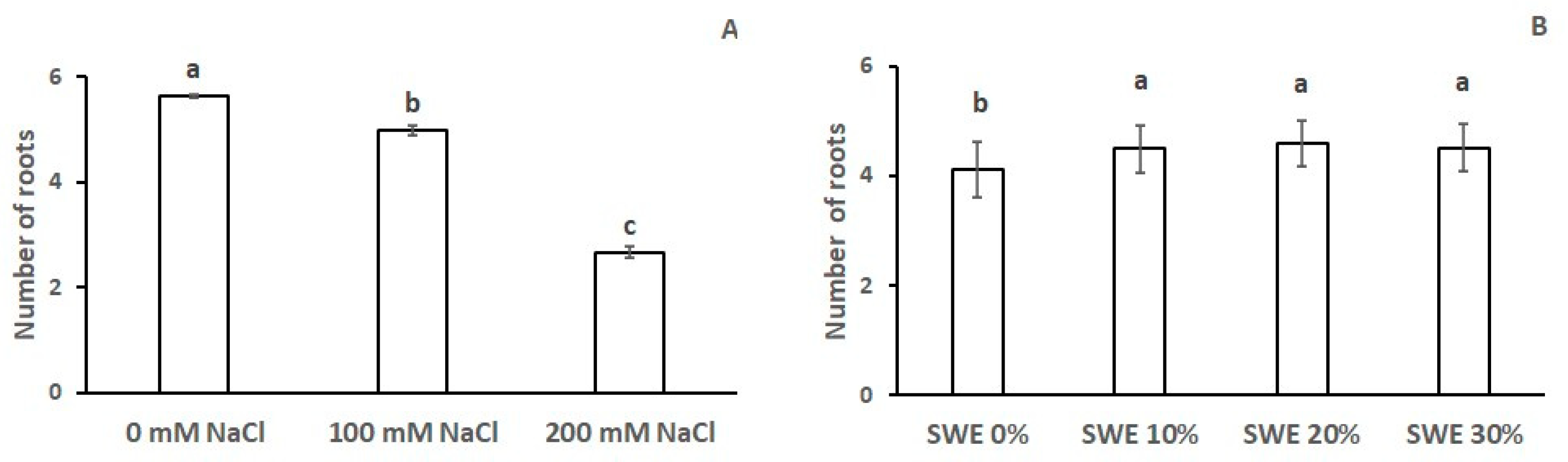
3.2. Number of Roots
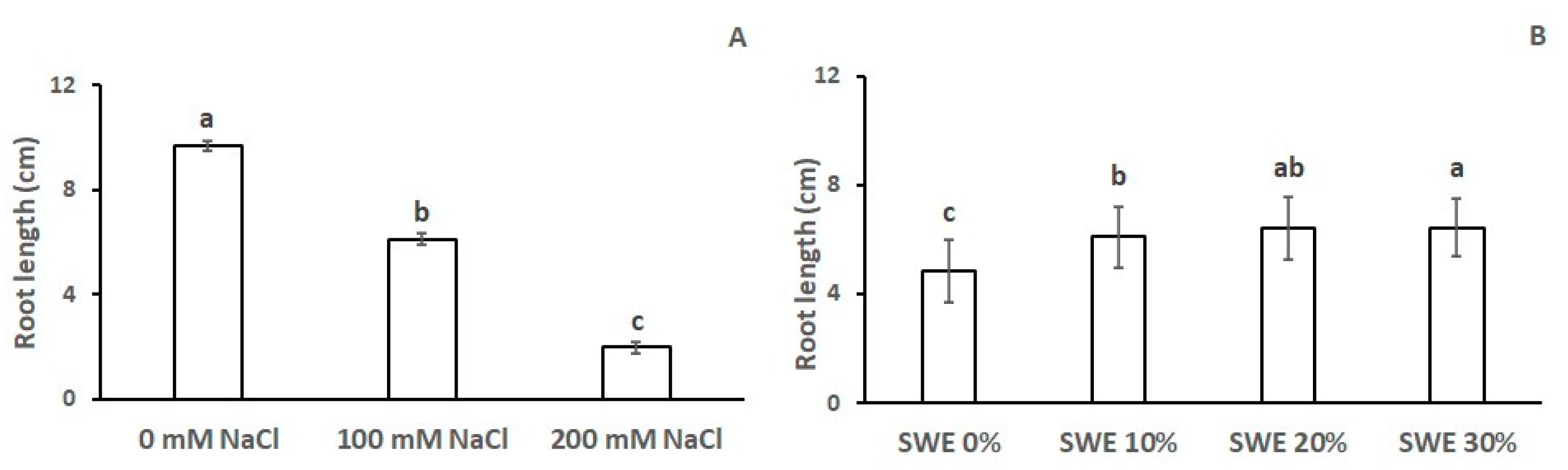
3.3. Length of the Main Root
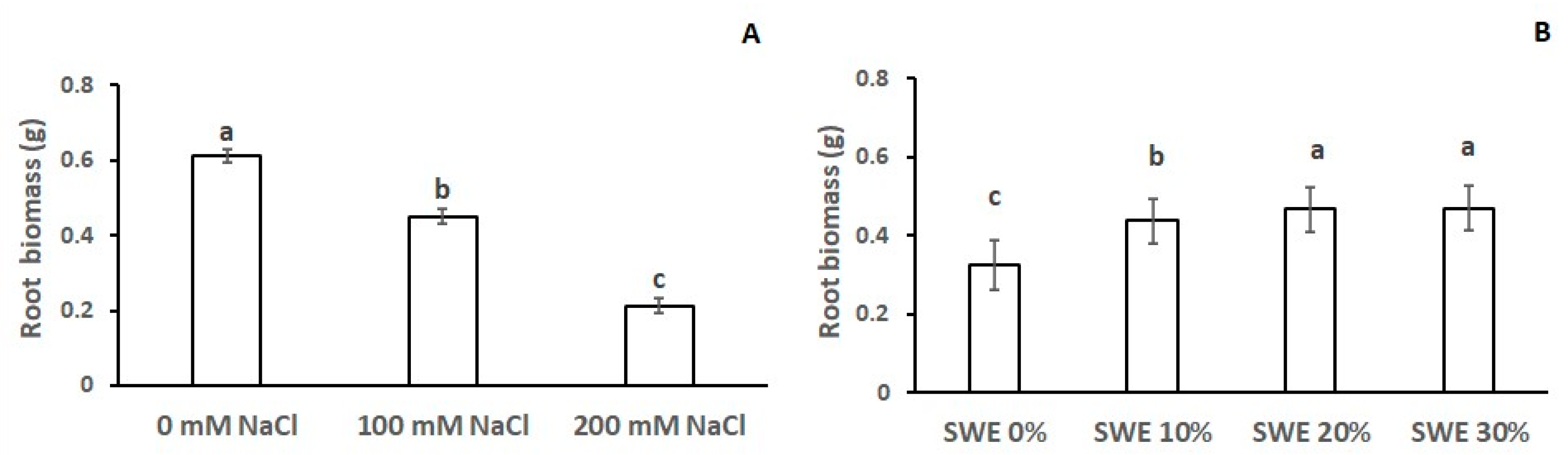
3.4. Root Biomass
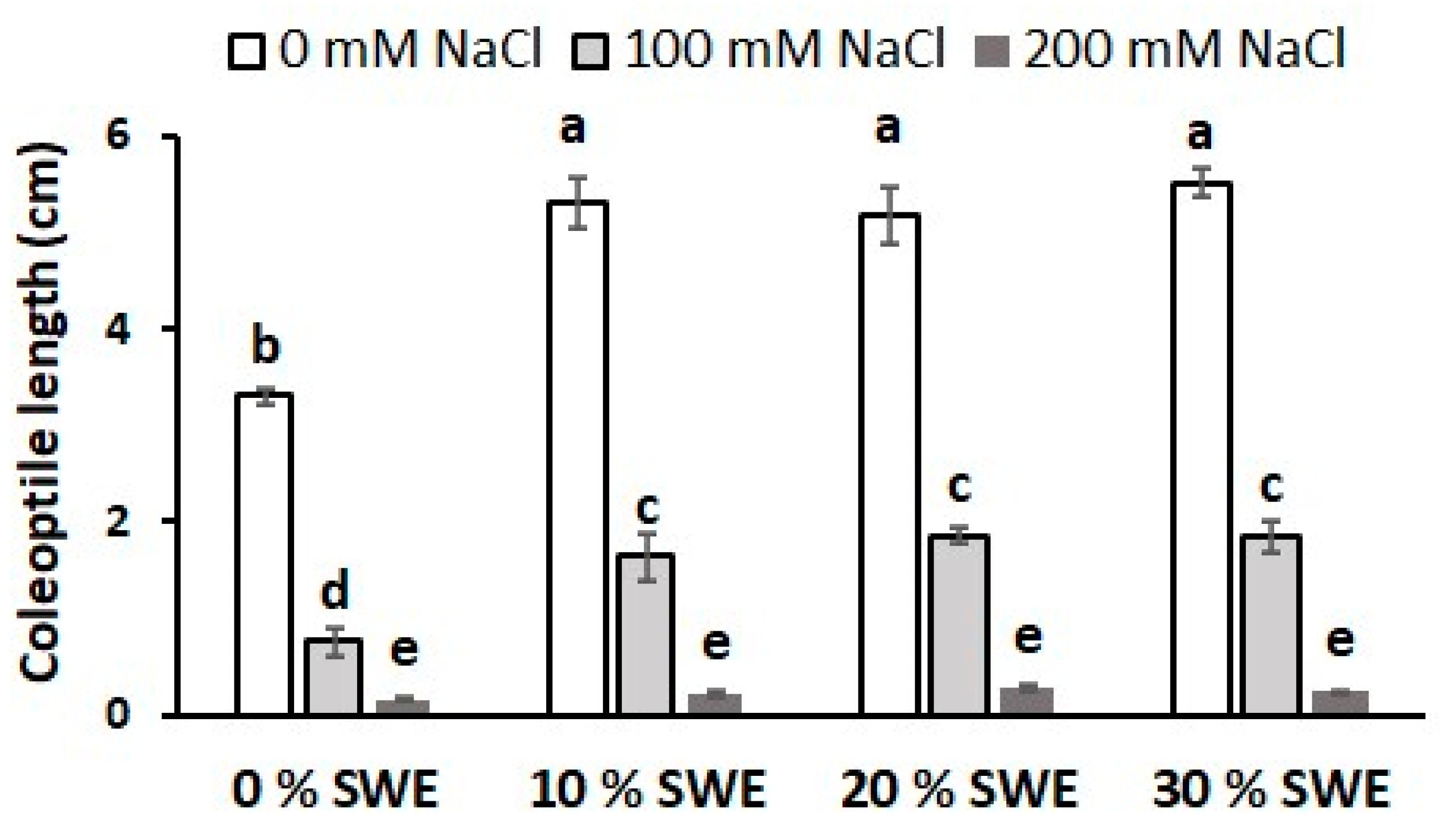
3.5. Coleoptile Length
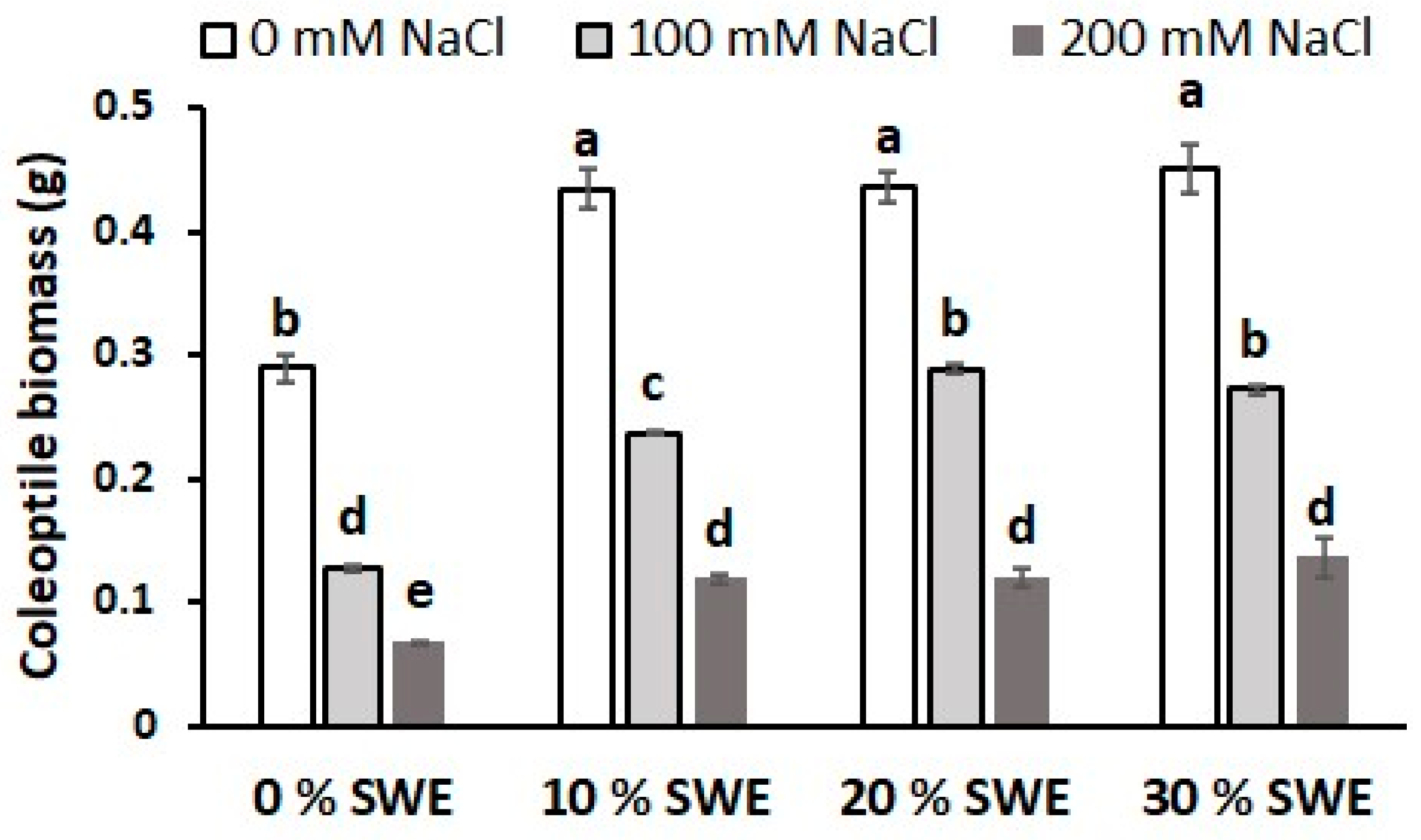
3.6. Coleoptile Biomass
4. Discussion
5. Conclusions and Future Prospects
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SWE | seaweed extract |
| 0% SWE | seed dressing solution containing 0 g of Codium fragile extract |
| 10% SWE | seed dressing solution containing 100 g of Codium fragile extract |
| 20% SWE | seed dressing solution containing 200 g of Codium fragile extract |
| 30% SWE | seed dressing solution containing 300 g of Codium fragile extract |
| T50 | median germination time |
References
- Minhas, P.S.; Ramos, T.B.; Ben-Gal, A.; Pereira, L.S. Coping with Salinity in Irrigated Agriculture: Crop Evapotranspiration and Water Management Issues. Agric. Water Manag. 2020, 227, 105832. [Google Scholar] [CrossRef]
- Pimentel, D.; Berger, B.; Filiberto, D.; Newton, M.; Wolfe, B.; Karabinakis, E.; Clark, S.; Poon, E.; Abbett, E.; Nandagopal, S. Water Resources: Agricultural and Environmental Issues. Bioscience 2004, 54, 909–918. [Google Scholar] [CrossRef]
- Jones, A.; Panagos, P.; Erhard, M.; Tóth, G.; Barcelo, S.; Bouraoui, F.; Bosco, C.; Dewitte, O.; Gardi, C.; Hervás, J.; et al. The State of Soil in Europe: A Contribution of the JRC to the European Environment Agency’s Environment State and Outlook Report—SOER 2010; Publications Office: Luxembourg, 2012; ISBN 9789279228063. [Google Scholar]
- Daliakopoulos, I.N.; Tsanis, I.K.; Koutroulis, A.; Kourgialas, N.N.; Varouchakis, A.E.; Karatzas, G.P.; Ritsema, C.J. The Threat of Soil Salinity: A European Scale Review. Sci. Total Environ. 2016, 573, 727–739. [Google Scholar] [CrossRef]
- Corwin, D.L. Climate Change Impacts on Soil Salinity in Agricultural Areas. Eur. J. Soil. Sci. 2021, 72, 842–862. [Google Scholar] [CrossRef]
- Machado, R.M.A.; Serralheiro, R.P. Soil Salinity: Effect on Vegetable Crop Growth. Management Practices to Prevent and Mitigate Soil Salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef]
- Stolte, J.; Tesfai, M.; Øygarden, L.; Kværnø, S.; Keizer, J.; Verheijen, F.; Panagos, P.; Ballabio, C.; Hessel, R. Soil Threats in Europe: Status, Methods, Drivers and Effects on Ecosystem Services. A Review Report, Deliverable 2.1 of the RECARE Project; European Commission DG Joint Research Centre: Luxembourg, 2015. [Google Scholar]
- Ferreira, C.S.S.; Seifollahi-Aghmiuni, S.; Destouni, G.; Ghajarnia, N.; Kalantari, Z. Soil Degradation in the European Mediterranean Region: Processes, Status and Consequences. Sci. Total Environ. 2022, 805, 20. [Google Scholar] [CrossRef]
- Bartels, D.; Sunkar, R. Drought and Salt Tolerance in Plants. CRC Crit. Rev. Plant Sci. 2005, 24, 23–58. [Google Scholar] [CrossRef]
- Bernstein, L. Osmotic Adjustment of Plants to Saline Media. I. Steady State. Am. J. Bot. 1961, 48, 909–918. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of Plant Responses to Salt Stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef] [PubMed]
- Cramer, W.; Guiot, J.; Fader, M.; Garrabou, J.; Gattuso, J.P.; Iglesias, A.; Lange, M.A.; Lionello, P.; Llasat, M.C.; Paz, S.; et al. Climate Change and Interconnected Risks to Sustainable Development in the Mediterranean. Nat. Clim. Chang. 2018, 8, 972–980. [Google Scholar] [CrossRef]
- USDA World Agricultural Production. Available online: https://www.fas.usda.gov/data/world-agricultural-production (accessed on 13 September 2023).
- FAO World Food Situation. Available online: https://www.fao.org/worldfoodsituation/csdb/en/ (accessed on 13 September 2023).
- Xynias, I.N.; Mylonas, I.; Korpetis, E.G.; Ninou, E.; Tsaballa, A.; Avdikos, I.D.; Mavromatis, A.G. Durum Wheat Breeding in the Mediterranean Region: Current Status and Future Prospects. Agronomy 2020, 10, 432. [Google Scholar] [CrossRef]
- Giambalvo, D.; Amato, G.; Badagliacca, G.; Ingraffia, R.; Di Miceli, G.; Frenda, A.S.; Plaia, A.; Venezia, G.; Ruisi, P. Switching from Conventional Tillage to No-Tillage: Soil N Availability, N Uptake, 15N Fertilizer Recovery, and Grain Yield of Durum Wheat. Field Crops Res. 2018, 218, 171–181. [Google Scholar] [CrossRef]
- Shafi, M.; Javed Khan, M.; Bakht, J.; Khan, M.A. Response of Wheat Genotypes to Salinity under Field Environment. Pak. J. Bot. 2013, 45, 787–794. [Google Scholar]
- Munns, R.; James, R.A. Screening Methods for Salinity Tolerance: A Case Study with Tetraploid Wheat. Plant Soil. 2003, 253, 201–218. [Google Scholar] [CrossRef]
- Foolad, M.R. Recent Advances in Genetics of Salt Tolerance in Tomato. Plant Cell Tissue Organ Cult. 2004, 76, 101–119. [Google Scholar] [CrossRef]
- Alaoui, M.M.; El Jourmi, L.; Ouarzane, A.; Lazar, S.; El Antri, S.; Zahouily, M.; Hmyene, A. Effet Du Stress Salin Sur La Germination et La Croissance de Six Variétés Marocaines de Blé (Effect of Salt Stress on Germination and Growth of Six Moroccan Wheat Varieties). J. Mater. Environ. Sci. 2013, 4, 997–1004. [Google Scholar]
- Hmissi, M.; Chaieb, M.; Krouma, A. Differences in the Physiological Indicators of Seed Germination and Seedling Establishment of Durum Wheat (Triticum durum Desf.) Cultivars Subjected to Salinity Stress. Agronomy 2023, 13, 1718. [Google Scholar] [CrossRef]
- Borrelli, G.; Ficco, D.; Giuzio, L.; Pompa, M.; Cattivelli, L.; Flagella, Z. Durum Wheat Salt Tolerance in Relation to Physiological, Yield and Quality Characters. Cereal Res. Commun. 2011, 39, 525–534. [Google Scholar] [CrossRef]
- Mbarki, S.; Skalicky, M.; Vachova, P.; Hajihashemi, S.; Jouini, L.; Zivcak, M.; Tlustos, P.; Brestic, M.; Hejnak, V.; Khelil, A.Z. Comparing Salt Tolerance at Seedling and Germination Stages in Local Populations of Medicago ciliaris L. to Medicago intertexta L. and Medicago scutellata L. Plants 2020, 9, 526. [Google Scholar] [CrossRef]
- Jacobsen, S.E.; Jensen, C.R.; Liu, F. Improving Crop Production in the Arid Mediterranean Climate. Field Crops Res. 2012, 128, 34–47. [Google Scholar] [CrossRef]
- Tomaz, A.; Palma, P.; Alvarenga, P.; Gonçalves, M.C. Soil Salinity Risk in a Climate Change Scenario and Its Effect on Crop Yield. In Climate Change and Soil Interactions; Elsevier: Amsterdam, The Netherlands, 2020; pp. 351–396. [Google Scholar] [CrossRef]
- Ronga, D.; Biazzi, E.; Parati, K.; Carminati, D.; Carminati, E.; Tava, A. Microalgal Biostimulants and Biofertilisers in Crop Productions. Agronomy 2019, 9, 192. [Google Scholar] [CrossRef]
- Arora, N.K. Impact of Climate Change on Agriculture Production and Its Sustainable Solutions. Environ. Sustain. 2019, 2, 95–96. [Google Scholar] [CrossRef]
- Mathlouthi, F.; Ruggeri, R.; Rossini, F. Alternative Solution to Synthetic Fertilizers for the Starter Fertilization of Bread Wheat under Mediterranean Climatic Conditions. Agronomy 2022, 12, 511. [Google Scholar] [CrossRef]
- Mathlouthi, F.; Ruggeri, R.; Rossini, A.; Rossini, F. A New Fertilization Approach for Bread Wheat in the Mediterranean Environment: Effects on Yield and Grain Protein Content. Agronomy 2022, 12, 2152. [Google Scholar] [CrossRef]
- Di Miceli, G.; Vultaggio, L.; Sabatino, L.; De Pasquale, C.; La Bella, S.; Consentino, B.B. Synergistic Effect of a Plant-Derived Protein Hydrolysate and Arbuscular Mycorrhizal Fungi on Eggplant Grown in Open Fields: A Two-Year Study. Horticulturae 2023, 9, 592. [Google Scholar] [CrossRef]
- Bulgari, R.; Cocetta, G.; Trivellini, A.; Vernieri, P.; Ferrante, A. Biostimulants and Crop Responses: A Review. Biol. Agric. Hortic. 2015, 31, 1–17. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The Role of Biostimulants and Bioeffectors as Alleviators of Abiotic Stress in Crop Plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [PubMed]
- Altındal, D. Effects of Seaweed Extract (SE) Applications on Seed Germination Characteristics of Wheat in Salinity Conditions. Int. J. Agric. For. Life Sci. 2019, 3, 115–120. [Google Scholar]
- Latique, S.; Aymen, E.M.; Halima, C.; Chérif, H.; Mimoun, E.K. Alleviation of Salt Stress in Durum Wheat (Triticum durum L.) Seedlings Through the Application of Liquid Seaweed Extracts of Fucus spiralis. Commun. Soil. Sci. Plant Anal. 2017, 48, 2582–2593. [Google Scholar] [CrossRef]
- Hernández-Herrera, R.M.; Santacruz-Ruvalcaba, F.; Ruiz-López, M.A.; Norrie, J.; Hernández-Carmona, G. Effect of Liquid Seaweed Extracts on Growth of Tomato Seedlings (Solanum lycopersicum L.). J. Appl. Phycol. 2014, 26, 619–628. [Google Scholar] [CrossRef]
- Makhaye, G.; Aremu, A.O.; Gerrano, A.S.; Tesfay, S.; Du Plooy, C.P.; Amoo, S.O. Biopriming with Seaweed Extract and Microbial-Based Commercial Biostimulants Influences Seed Germination of Five Abelmoschus esculentus Genotypes. Plants 2021, 10, 1327. [Google Scholar] [CrossRef]
- Eugenia, M.; Carvalho, A.; Roberto, P.; Dionisia, A.; Novembre, L.C.; Carvalho, M.E.A.; Castro, P.R.C.; Novembre, A.D.C.; Chamma, H.M.C.P. Seaweed Extract Improves the Vigor and Provides the Rapid Emergence of Dry Bean Seeds. J. Agric. Environ. Sci. 2013, 13, 1104–1107. [Google Scholar] [CrossRef]
- Chanthini, K.M.P.; Senthil-Nathan, S.; Pavithra, G.S.; Malarvizhi, P.; Murugan, P.; Deva-Andrews, A.; Janaki, M.; Sivanesh, H.; Ramasubramanian, R.; Stanley-Raja, V.; et al. Aqueous Seaweed Extract Alleviates Salinity-Induced Toxicities in Rice Plants (Oryza sativa L.) by Modulating Their Physiology and Biochemistry. Agriculture 2022, 12, 2049. [Google Scholar] [CrossRef]
- Gul, S.; Abideen, Z.; Adnan, M.Y.; Hanif, M.; Nawaz, M.F.; Shumaila; Keblawy, A.E. Seaweed-Derived Bio-Stimulant Improves Growth and Salt Tolerance of Radish Varieties under Saline Conditions. Biocatal. Agric. Biotechnol. 2023, 52, 102822. [Google Scholar] [CrossRef]
- Supraja, K.V.; Behera, B.; Balasubramanian, P. Efficacy of Microalgal Extracts as Biostimulants through Seed Treatment and Foliar Spray for Tomato Cultivation. Ind. Crops Prod. 2020, 151, 112453. [Google Scholar] [CrossRef]
- Radwan, A.M.; Ahmed, E.A.; Donia, A.M.; Mustafa, A.E.; Balah, M.A. Priming of Citrullus lanatus Var. colocynthoides Seeds in Seaweed Extract Improved Seed Germination, Plant Growth and Performance under Salinity Conditions. Sci. Rep. 2023, 13, 11884. [Google Scholar] [CrossRef]
- Hussein, M.H.; Eltanahy, E.; Al Bakry, A.F.; Elsafty, N.; Elshamy, M.M. Seaweed Extracts as Prospective Plant Growth Bio-Stimulant and Salinity Stress Alleviator for Vigna sinensis and Zea mays. J. Appl. Phycol. 2021, 33, 1273–1291. [Google Scholar] [CrossRef]
- Nezamdoost, D.; Ghahremani, Z.; Akbari, M.B.; Barzegar, T.; Ranjbar, M.E. Can Seed Priming with Seaweed Extract Neutralize the Effects of Salinity on New Red Fire Leafy Lettuce Characteristics? Gesunde Pflanz. 2023, 75, 955–969. [Google Scholar] [CrossRef]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed Extracts as Biostimulants in Horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Makhaye, G.; Mofokeng, M.M.; Tesfay, S.; Aremu, A.O.; Van Staden, J.; Amoo, S.O. Chapter 5—Influence of Plant Biostimulant Application on Seed Germination. In Biostimulants for Crops from Seed Germination to Plant Development A Practical Approach; Academic Press: Cambridge, MA, USA, 2021; pp. 109–135. [Google Scholar] [CrossRef]
- Taylor, A.G.; Allen, P.S.; Bennett, M.A.; Bradford, K.J.; Burris, J.S.; Misra, M.K. Seed Enhancements. Seed Sci. Res. 1998, 8, 245–256. [Google Scholar] [CrossRef]
- Amirkhani, M.; Mayton, H.S.; Netravali, A.N.; Taylor, A.G. A Seed Coating Delivery System for Bio-Based Biostimulants to Enhance Plant Growth. Sustainability 2019, 11, 5304. [Google Scholar] [CrossRef]
- Sghaier, Y.R.; Zakhama-Sraieb, R.; Mouelhi, S.; Vazquez, M.; Valle, C.; Ramos-Espla, A.A.; Astier, J.M.; Verlaque, M.; Charfi-Cheikhrouha, F. Review of Alien Marine Macrophytes in Tunisia. Mediterr. Mar. Sci. 2016, 17, 109–123. [Google Scholar] [CrossRef]
- Cherif, W.; Ktari, L.; El Bour, M.; Boudabous, A.; Grignon-Dubois, M. Codium fragile Subsp. fragile (Suringar) Hariot in Tunisia: Morphological Data and Status of Knowledge. Algae 2016, 31, 129–136. [Google Scholar] [CrossRef]
- El-Din, S.M.M. Utilization of Seaweed Extracts as Bio-Fertilizers to Stimulate the Growth of Wheat Seedlings. Egypt. Soc. Exp. Biol. 2015, 11, 31–39. [Google Scholar]
- Nessim, A. Mitigation of Lead Stress in Triticum aestivum by Seed Priming in Aqueous Extracts of The Macroalgea Halimeda opuntia and Codium fragile. Egypt. J. Bot. 2018, 58, 263–274. [Google Scholar] [CrossRef]
- Demir, N.; Dural, B.; Yildirim, K. Effect of Seaweed Suspensions on Seed Germination of Tomato, Pepper and Aubergine. J. Biol. Sci. 2006, 6, 1130–1133. [Google Scholar] [CrossRef]
- Righini, H.; Francioso, O.; Di Foggia, M.; Prodi, A.; Quintana, A.M.; Roberti, R. Tomato Seed Biopriming with Water Extracts from Anabaena minutissima, Ecklonia maxima and Jania adhaerens as a New Agro-Ecological Option against Rhizoctonia solani. Sci. Hortic. 2021, 281, 109921. [Google Scholar] [CrossRef]
- Anisimov, M.M.; Klykov, A.G. Metabolites of Terrestrial Plants and Marine Organisms as Potential Regulators of Growth of Agricultural Plants in the Russian Far East. J. Agric. Sci. 2014, 6, 88. [Google Scholar] [CrossRef]
- Royo, A.; Abiò, D. Salt Tolerance in Durum Wheat Cultivars. Span J. Agric. Res. 2003, 1, 27–35. [Google Scholar] [CrossRef]
- Cuong, D.M.; Kwon, S.J.; Van Nguyen, B.; Chun, S.W.; Kim, J.K.; Park, S.U. Effect of Salinity Stress on Phenylpropanoid Genes Expression and Related Gene Expression in Wheat Sprout. Agronomy 2020, 10, 390. [Google Scholar] [CrossRef]
- Balducci, E.; Tini, F.; Beccari, G.; Ricci, G.; Ceron-Bustamante, M.; Orfei, M.; Guiducci, M.; Covarelli, L. A Two-Year Field Experiment for the Integrated Management of Bread and Durum Wheat Fungal Diseases and of Deoxynivalenol Accumulation in the Grain in Central Italy. Agronomy 2022, 12, 840. [Google Scholar] [CrossRef]
- Ministero dell’Agricoltura e delle Foreste. Metodi Ufficiali di Analisi per le Sementi. Gazz. Uff. Della Repubb. Ital. 1993.
- Soltani, E.; Ghaderi-Far, F.; Baskin, C.C.; Baskin, J.M. Problems with Using Mean Germination Time to Calculate Rate of Seed Germination. Aust. J. Bot. 2015, 63, 631–635. [Google Scholar] [CrossRef]
- Scarici, E.; Ruggeri, R.; Provenzano, M.E.; Rossini, F. Germination and Performance of Seven Native Wildflowers in the Mediterranean Landscape Plantings. Imp. Jpn. Army 2018, 13, 163–171. [Google Scholar] [CrossRef]
- Farooq, M.; Basra, S.M.A.; Ahmad, N.; Hafeez, K. Thermal Hardening: A New Seed Vigor Enhancement Tool in Rice. J. Integr. Plant Biol. 2005, 47, 187–193. [Google Scholar] [CrossRef]
- Ritz, C.; Pipper, C.B.; Streibig, J.C. Analysis of Germination Data from Agricultural Experiments. Eur. J. Agron. 2013, 45, 1–6. [Google Scholar] [CrossRef]
- Rossini, F.; Ruggeri, R.; Celli, T.; Rogai, F.M.; Kuzmanović, L.; Richardson, M.D. Cool-Season Grasses for Overseeding Sport Turfs: Germination and Performance under Limiting Environmental Conditions. HortScience 2019, 54, 555–563. [Google Scholar] [CrossRef]
- R Core Team A Language and Environment for Statistical Computing 2021. Available online: https://www.R-project.org/ (accessed on 17 September 2023).
- Onofri, A.; Mesgaran, M.B.; Ritz, C. A Unified Framework for the Analysis of Germination, Emergence, and Other Time-to-Event Data in Weed Science. Weed Sci. 2022, 70, 259–271. [Google Scholar] [CrossRef]
- Ayed, S.; Rassaa, N.; Chamekh, Z.; Beji, S.; Karoui, F.; Bouzaien, T.; Mrabit, M.; Ben, Y.M. Effect of Salt Stress (Sodium Chloride) on Germination and Seedling Growth of Durum Wheat (Triticum durum Desf.) Genotypes. Int. J. Biodivers. Conserv. 2014, 6, 320–325. [Google Scholar] [CrossRef]
- Hussain, S.; Khaliq, A.; Matloob, A.; Wahid, M.A.; Afzal, I. Germination and Growth Response of Three Wheat Cultivars to NaCl Salinity. Soil. Environ. 2013, 32, 36–43. [Google Scholar]
- Latique, S.; Aymen Elouaer, M.; Chernane, H.; Hannachi, C.; Elkaoua, M. Effect of Seaweed Liquid Extract of Sargassum vulgare on Growth of Durum Wheat Seedlings (Triticum durum L.) under Salt Stress. Int. J. Innov. Appl. Stud. 2014, 7, 1430–1435. [Google Scholar]
- Feghhenabi, F.; Hadi, H.; Khodaverdiloo, H.; van Genuchten, M.T. Seed Priming Alleviated Salinity Stress during Germination and Emergence of Wheat (Triticum aestivum L.). Agric. Water Manag. 2020, 231, 106022. [Google Scholar] [CrossRef]
- Bonomelli, C.; Celis, V.; Lombardi, G.; Mártiz, J. Salt Stress Effects on Avocado (Persea americana Mill.) Plants with and without Seaweed Extract (Ascophyllum nodosum) Application. Agronomy 2018, 8, 64. [Google Scholar] [CrossRef]
- De Santis, M.A.; Soccio, M.; Laus, M.N.; Flagella, Z. Influence of Drought and Salt Stress on Durum Wheat Grain Quality and Composition: A Review. Plants 2021, 10, 2599. [Google Scholar] [CrossRef]
- Pastuszak, J.; Dziurka, M.; Hornyák, M.; Szczerba, A.; Kopeć, P.; Płażek, A. Physiological and Biochemical Parameters of Salinity Resistance of Three Durum Wheat Genotypes. Int. J. Mol. Sci. 2022, 23, 8397. [Google Scholar] [CrossRef]
- Pessarakli, M. Handbook of Plant and Crop Stress; Dekker, M., Ed.; CRC Press: Boca Raton, FL, USA, 1999; ISBN 0824719484. [Google Scholar]
- Hamed, S.M.; Abd El-Rhman, A.A.; Abdel-Raouf, N.; Ibraheem, I.B.M. Role of Marine Macroalgae in Plant Protection & Improvement for Sustainable Agriculture Technology. Beni. Suef. Univ. J. Basic. Appl. Sci. 2018, 7, 104–110. [Google Scholar] [CrossRef]
- Sadak, M.; El-Bassiouny, H.; Mahfouz, S.; El-Enany, M.; Elewa, T. Use of Thiamine, Pyridoxine and Biostimulant for Better Yield of Wheat Plants under Water Stress: Growth, Osmoregulations, Antioxidantive Defense and Protein Pattern. Egypt. J. Chem. 2022, 66, 407–424. [Google Scholar] [CrossRef]
- Stirk, W.A.; Van Staden, J. Comparison of Cytokinin- and Auxin-like Activity in Some Commercially Used Seaweed Extracts; Kluwer Academic Publishers: New York, NY, USA, 1997; Volume 8. [Google Scholar]
- Iqbal, M.; Ashraf, M. Presowing Seed Treatment with Cytokinins and Its Effect on Growth, Photosynthetic Rate, Ionic Levels and Yield of Two Wheat Cultivars Differing in Salt Tolerance. J. Integr. Plant Biol. 2005, 47, 1315–1325. [Google Scholar] [CrossRef]
- Iqbal, M.; Ashraf, M. Seed Treatment with Auxins Modulates Growth and Ion Partitioning in Salt-Stressed Wheat Plants. J. Integr. Plant Biol. 2007, 49, 1003–1015. [Google Scholar] [CrossRef]
- Iqbal, M.; Ashraf, M.; Jamil, A. Seed Enhancement with Cytokinins: Changes in Growth and Grain Yield in Salt Stressed Wheat Plants. Plant Growth Regul. 2006, 50, 29–39. [Google Scholar] [CrossRef]
- Ribba, T.; Garrido-Vargas, F.; O’Brien, J.A. Auxin-Mediated Responses under Salt Stress: From Developmental Regulation to Biotechnological Applications. J. Exp. Bot. 2020, 71, 3843–3853. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Foolad, M.R. Pre-Sowing Seed Treatment—A Shotgun Approach to Improve Germination, Plant Growth, and Crop Yield Under Saline and Non-Saline Conditions. Adv. Agron. 2005, 88, 223–271. [Google Scholar] [CrossRef]
- Lutts, S.; Majerus, V.; Kinet, J.M. NaCl Effects on Proline Metabolism in Rice (Oryza sativa) Seedlings. Physiol. Plant 1999, 105, 450–458. [Google Scholar] [CrossRef]
- Javid, M.G.; Sorooshzadeh, A.; Moradi, F.; Ali, S.; Sanavy, M.M.; Allahdadi, I. The Role of Phytohormones in Alleviating Salt Stress in Crop Plants. Aust. J. Crop Sci. 2011, 5, 726–734. [Google Scholar]
- Kuiper, D.; Schuit, J.; Kuiper, P.J.C. Actual Cytokinin Concentrations in Plant Tissue as an Indicator for Salt Resistance in Cereals. Plant Soil 1990, 123, 243–250. [Google Scholar] [CrossRef]
- Ortiz, J.; Uquiche, E.; Robert, P.; Romero, N.; Quitral, V.; Llantén, C. Functional and Nutritional Value of the Chilean Seaweeds Codium fragile, Gracilaria chilensis and Macrocystis pyrifera. Eur. J. Lipid Sci. Technol. 2009, 111, 320–327. [Google Scholar] [CrossRef]
- Salama, K.H.A.; Mansour, M.M.F.; Al-Malawi, H.A. Glycinebetaine Priming Improves Salt Tolerance of Wheat. Biologia 2015, 70, 1334–1339. [Google Scholar] [CrossRef]
- Mansour, M.M.F.; Ali, E.F. Evaluation of Proline Functions in Saline Conditions. Phytochemistry 2017, 140, 52–68. [Google Scholar]
- Ambreen, S.; Athar, H.u.R.; Khan, A.; Zafar, Z.U.; Ayyaz, A.; Kalaji, H.M. Seed Priming with Proline Improved Photosystem II Efficiency and Growth of Wheat (Triticum aestivum L.). BMC Plant Biol. 2021, 21, 502. [Google Scholar] [CrossRef] [PubMed]
- Khafagy, M.A.; Arafa, A.A.; El-Banna, M.F. Glycinebetaine and Ascorbic Acid Can Alleviate the Harmful Effects of Nacl Salinity in Sweet Pepper. Aust. J. Crop Sci. 2009, 3, 257–267. [Google Scholar]
- Liu, H.-L.; Sha, H.-j.; Wang, J.-g.; Liu, Y.; Zou, D.-t.; Zhao, H.-w. Effect of Seed Soaking with Exogenous Proline on Seed Germination of Rice Under Salt Stress. J. Northeast Agric. Univ. 2014, 21, 1–6. [Google Scholar] [CrossRef]
- Mahboob, W.; Athar Khan, M.; Shirazi, M.U. Induction of Salt Tolerance in Wheat Triticum aestivum L. Seedlings through Exogenous Application of Proline. Pak. J. Bot. 2016, 48, 861–867. [Google Scholar]
- Yang, Y.; Guo, Y. Elucidating the Molecular Mechanisms Mediating Plant Salt-Stress Responses. New Phytol. 2018, 217, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Quitadamo, F.; De Simone, V.; Beleggia, R.; Trono, D. Chitosan-Induced Activation of the Antioxidant Defense System Counteracts the Adverse Effects of Salinity in Durum Wheat. Plants 2021, 10, 1365. [Google Scholar] [CrossRef]
- Mehmood, S.; Khan, A.A.; Shi, F.; Tahir, M.; Sultan, T.; Munis, M.F.H.; Kaushik, P.; Alyemeni, M.N.; Chaudhary, H.J. Alleviation of Salt Stress in Wheat Seedlings via Multifunctional Bacillus aryabhattai pm34: An in-Vitro Study. Sustainability 2021, 13, 8030. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Aslam, M.T.; Alhammad, B.A.; Hassan, M.U.; Maqbool, R.; Chattha, M.U.; Khan, I.; Gitari, H.I.; Uslu, O.S.; Roy, R.; et al. Salinity Stress in Wheat: Effects, Mechanisms and Management Strategies. Phyton-Int. J. Exp. Bot. 2022, 91, 667–694. [Google Scholar] [CrossRef]
- Papenfus, H.B.; Kulkarni, M.G.; Stirk, W.A.; Finnie, J.F.; Van Staden, J. Effect of a Commercial Seaweed Extract (Kelpak®) and Polyamines on Nutrient-Deprived (N, P and K) Okra Seedlings. Sci. Hortic. 2013, 151, 142–146. [Google Scholar] [CrossRef]
- Ferreira, M.; Salgado, J.M.; Fernandes, H.; Peres, H.; Belo, I. Potential of Red, Green and Brown Seaweeds as Substrates for Solid State Fermentation to Increase Their Nutritional Value and to Produce Enzymes. Foods 2022, 11, 3864. [Google Scholar] [CrossRef]
- Itroutwar, P.D.; Kasivelu, G.; Raguraman, V.; Malaichamy, K.; Sevathapandian, S.K. Effects of Biogenic Zinc Oxide Nanoparticles on Seed Germination and Seedling Vigor of Maize (Zea mays). Biocatal. Agric. Biotechnol. 2020, 29, 101778. [Google Scholar] [CrossRef]
- Kamran, M.; Wang, D.; Xie, K.; Lu, Y.; Shi, C.; EL Sabagh, A.; Gu, W.; Xu, P. Pre-Sowing Seed Treatment with Kinetin and Calcium Mitigates Salt Induced Inhibition of Seed Germination and Seedling Growth of Choysum (Brassica rapa Var. parachinensis). Ecotoxicol. Environ. Saf. 2021, 227, 112921. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Gao, Y.; Wang, X.; Li, S.; Zheng, H.; Chen, Z.; Wu, F.; Du, X.; Sui, N. Exogenous Calcium Application Enhances Salt Tolerance of Sweet Sorghum Seedlings. J. Agron. Crop Sci. 2022, 208, 441–453. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Condition | SWE | Final Germination (%) | T50 (d) |
|---|---|---|---|
| Salinity 0 mM NaCl | 0% | 89.34 ± 1.28 b | 1.65 ± 0.02 a |
| 10% | 92.70 ± 1.45 a | 1.62 ± 0.05 a | |
| 20% | 93.13 ± 0.96 a | 1.71 ± 0.05 a | |
| 30% | 94.82 ± 0.79 a | 1.73 ± 0.03 a | |
| Salinity 100 mM NaCl | 0% | 90.10 ± 0.55 c | 2.15 ± 0.01 a |
| 10% | 92.79 ± 0.77 b | 2.03 ± 0.04 b | |
| 20% | 96.05 ± 0.60 a | 2.01 ± 0.03 b | |
| 30% | 96.05 ± 0.60 a | 2.04 ± 0.01 b | |
| Salinity 200 mM NaCl | 0% | 60.67 ± 1.08 c | 3.43 ± 0.11 a |
| 10% | 82.21 ± 1.22 b | 2.95 ± 0.07 bc | |
| 20% | 85.65 ± 0.51 ab | 3.13 ± 0.07 ab | |
| 30% | 87.70 ± 1.54 a | 2.77 ± 0.12 c |
| Number of Roots | Length of the Main Root | Root Biomass | Coleoptile Length | Coleoptile Biomass | |
|---|---|---|---|---|---|
| SWE | *** | *** | *** | *** | *** |
| Salinity | *** | *** | *** | *** | *** |
| SWE × Salinity | n.s. | n.s. | n.s. | *** | *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossini, A.; Ruggeri, R.; Mzid, N.; Rossini, F.; Di Miceli, G. Codium fragile (Suringar) Hariot as Biostimulant Agent to Alleviate Salt Stress in Durum Wheat: Preliminary Results from Germination Trials. Plants 2024, 13, 283. https://doi.org/10.3390/plants13020283
Rossini A, Ruggeri R, Mzid N, Rossini F, Di Miceli G. Codium fragile (Suringar) Hariot as Biostimulant Agent to Alleviate Salt Stress in Durum Wheat: Preliminary Results from Germination Trials. Plants. 2024; 13(2):283. https://doi.org/10.3390/plants13020283
Chicago/Turabian StyleRossini, Angelo, Roberto Ruggeri, Nada Mzid, Francesco Rossini, and Giuseppe Di Miceli. 2024. "Codium fragile (Suringar) Hariot as Biostimulant Agent to Alleviate Salt Stress in Durum Wheat: Preliminary Results from Germination Trials" Plants 13, no. 2: 283. https://doi.org/10.3390/plants13020283
APA StyleRossini, A., Ruggeri, R., Mzid, N., Rossini, F., & Di Miceli, G. (2024). Codium fragile (Suringar) Hariot as Biostimulant Agent to Alleviate Salt Stress in Durum Wheat: Preliminary Results from Germination Trials. Plants, 13(2), 283. https://doi.org/10.3390/plants13020283