


Quality Characteristics of Twelve Advanced Lines of Avena magna ssp. domestica Grown in Three Contrasting Locations in Morocco

 , ,
, ,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Materials
2.2. Chemical Analysis
2.3. Statistical Analysis
3. Results and Discussion
3.1. Chemical Composition
3.2. Mineral Composition
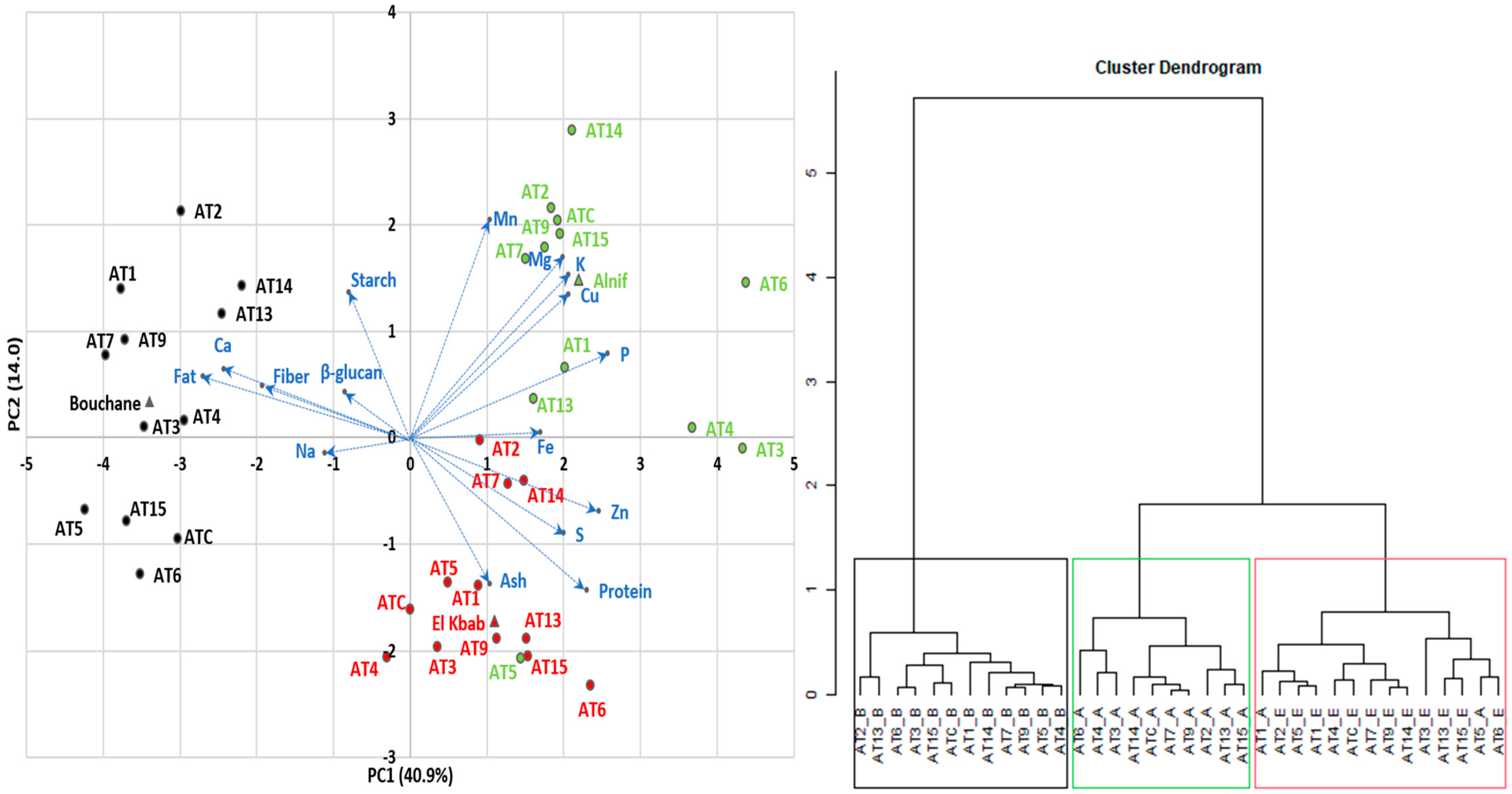
3.3. Multivariate Analysis
3.3.1. Correlations
3.3.2. Principal Component Analysis
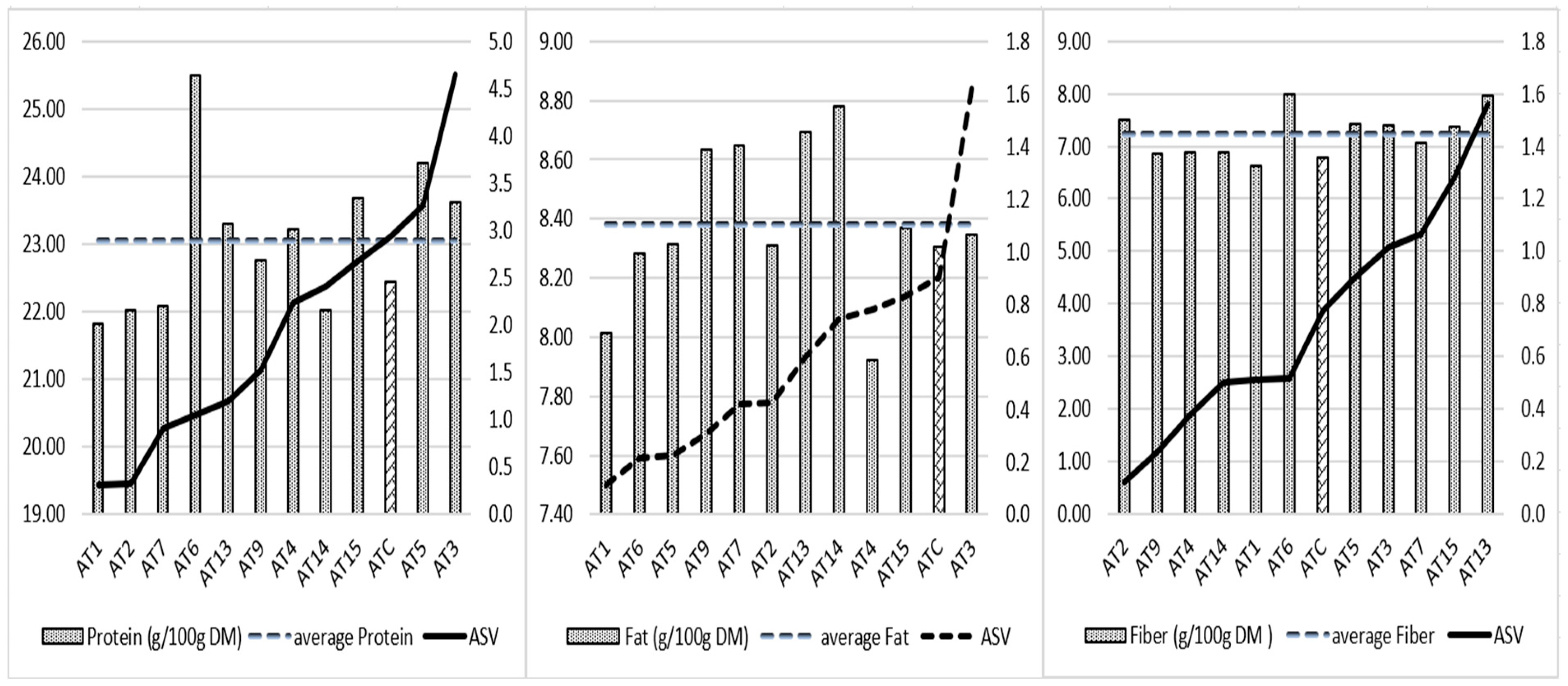
3.3.3. Analysis of Protein, Fat, and Fiber Stability of 12 Lines of A. magna domestica ssp.
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations, Department of Economic and Social Affairs, Population Division. World Population Prospects 2019: Highlights; (ST/ESA/SER.A/423); United Nations: New York, NY, USA, 2019. [Google Scholar]
- Falcon, W.P.; Naylor, R.L.; Shankar, N.D. Rethinking global food demand for 2050. Popul. Dev. Rev. 2022, 48, 921–957. [Google Scholar] [CrossRef]
- Satterthwaite, D.; McGranahan, G.; Tacoli, C. Urbanization and its implications for food and farming. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2809–2820. [Google Scholar] [CrossRef]
- UNPD. World Urbanization Prospects: The 2018 Revision; United Nations, Department of Economic and Social Affairs, Population Division: New York, NY, USA, 2018. [Google Scholar]
- Jacott, C.N.; Boden, S.A. Feeling the heat: Developmental and molecular responses of wheat and barley to high ambient temperatures. J. Exp. Bot. 2020, 71, 5740–5751. [Google Scholar] [CrossRef] [PubMed]
- Drewnowski, A.; Finley, J.; Hess, J.M.; Ingram, J.; Miller, G.; Peters, C. Toward healthy diets from sustainable food systems. Curr. Dev. Nutr. 2020, 4, nzaa083. [Google Scholar] [CrossRef] [PubMed]
- Alaimo, K.; Chilton, M.; Jones, S.J. Food insecurity, hunger, and malnutrition. In Present Knowledge in Nutrition; Academic Press: Cambridge, MA, USA, 2020; pp. 311–326. [Google Scholar]
- Loskutov, I.G.; Khlestkina, E.K. Wheat, Barley, and Oat Breeding for Health Benefit Components in Grain. Plants 2021, 10, 86. [Google Scholar] [CrossRef] [PubMed]
- Pal, M.; Molnár, J. Growing Importance of Cereals in Nutrition and Healthy Life. Int. J. Food Sci. Agric. 2021, 5, 275–277. Available online: http://www.hillpublisher.com/journals/jsfa (accessed on 25 February 2023).
- Song, M.; Wu, K.; Meyerhardt, J.A.; Ogino, S.; Wang, M.; Fuchs, C.S.; Giovannucci, E.L.; Chan, A.T. Fiber Intake and Survival After Colorectal Cancer Diagnosis. JAMA Oncol. 2018, 4, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Paudel, D.; Dhungana, B.; Caffe, M.; Krishnan, P. A review of health-beneficial properties of oats. Foods 2021, 10, 2591. [Google Scholar] [CrossRef]
- Yao, N.; Jannink, J.L.; White, P.J. Molecular Weight Distribution of (1→3)(1→4)-β-Glucan Affects Pasting Properties of Flour from Oat Lines with High and Typical Amounts of β-Glucan. Cereal Chem. 2007, 84, 471–479. [Google Scholar] [CrossRef]
- Boczkowska, M.; Podyma, W.; Łapiński, B. Oat. In Genetic and Genomic Resources for Grain Cereals Improvement; Academic Press: Cambridge, MA, USA, 2016; pp. 159–225. [Google Scholar]
- Ahola, H.G.; Sontag-Strohm, T.S.; Schulman, A.H.; Tanhuanpää, P.; Viitala, S.; Huang, X. Immunochemical analysis of oat avenins in an oat cultivar and landrace collection. J. Cereal Sci. 2020, 95, 103053. [Google Scholar] [CrossRef]
- Németh, R.; Turóczi, F.; Csernus, D.; Solymos, F.; Jaksics, E.; Tömösközi, S. Characterization of chemical composition and techno-functional properties of oat cultivars. Cereal Chem. 2021, 98, 1183–1192. [Google Scholar] [CrossRef]
- Givens, D.; Davies, T.; Laverick, R. Effect of variety, nitrogen fertiliser and various agronomic factors on the nutritive value of husked and naked oats grain. Anim. Feed. Sci. Technol. 2004, 113, 169–181. [Google Scholar] [CrossRef]
- Bari, A. Oats in Morocco. 2016. Available online: https://www.researchgate.net/publication/31057526.12 (accessed on 22 February 2022).
- Tyack, N.; Dempewolf, H. The economics of crop wild relatives under climate change. In Crop Wild Relatives and Climate Change; John Wiley & Sons: Hoboken, NJ, USA, 2015; pp. 281–291. [Google Scholar]
- Bohra, A.; Kilian, B.; Sivasankar, S.; Caccamo, M.; Mba, C.; McCouch, S.R.; Varshney, R.K. Reap the crop wild relatives for breeding future crops. Trends Biotechnol. 2022, 40, 412–431. [Google Scholar] [CrossRef] [PubMed]
- Gutaker, R.M.; Chater, C.C.; Brinton, J.; Castillo-Lorenzo, E.; Breman, E.; Pironon, S. Scaling up neodomestication for climate-ready crops. Curr. Opin. Plant Biol. 2022, 66, 102169. [Google Scholar] [CrossRef]
- Von Wettberg, E.; Davis, T.M.; Smýkal, P. Wild plants as source of new crops. Front. Plant Sci. 2020, 11, 591554. [Google Scholar] [CrossRef] [PubMed]
- Kahane, R.; Hodgkin, T.; Jaenicke, H.; Hoogendoorn, C.; Hermann, M.; Keatinge, J.D.H.; Hughes, J.D.; Padulosi, S.; Looney, N. Agrobiodiversity for food security, health and income. Agron. Sustain. Dev. 2013, 33, 671–693. [Google Scholar] [CrossRef]
- Mohler, V.; Paczos-Grzęda, E.; Sowa, S. Loving the Alien: The Contribution of the Wild in Securing the Breeding of Cultivated Hexaploid Wheat and Oats. Agriculture 2023, 13, 2060. [Google Scholar] [CrossRef]
- Lyrene, P.M.; Shands, H.L. Heading dates in six Avena sativa L. × A. sterilis L. crosses. Crop Sci. 1975, 15, 359–360. [Google Scholar] [CrossRef]
- Ladizinsky, G. Domestication via hybridization of the wild tetraploid oats Avena magna and A. murphyi. Theor. Appl. Genet. 1995, 91, 639–646. [Google Scholar] [CrossRef]
- Jackson, E.W. High Protein Oat Species. W.I.P.O. Patent Application WO 2017/070104 A1, 27 April 2017. [Google Scholar]
- Palmgren, M.G.; Clemens, S.; Williams, L.E.; Krämer, U.; Borg, S.; Schjørring, J.K.; Sanders, D. Zinc biofortification of cereals: Problems and solutions. Trends Plant Sci. 2008, 13, 464–473. [Google Scholar] [CrossRef]
- Jellen, E.N.; Jackson, E.W.; Elhadji, T.; Young, L.K.; El Mouttaqi, A.; Al Halfa, I.; El Fartassi, I.; Katile, L.S.; Linchangco, R.; Klassen, K.; et al. Adaptation and agronomic performance of domesticated Moroccan Oat (Avena magna ssp. domestica) lines under subsistence farming conditions at multiple locations in Morocco. Agronomy 2021, 11, 1037. [Google Scholar] [CrossRef]
- Thiam, E.H.; Jellen, E.N.; Jackson, E.W.; Nelson, M.; Rogers, W.; El Mouttaqi, A.; Benlhabib, O. Productivity and stability evaluation of 12 selected Avena magna ssp. domestica lines based on multi-location experiments during three cropping seasons in Morocco. Agriculture 2023, 13, 1486. [Google Scholar]
- Manzali, R.; Douaik, A.; Bouksaim, M.; Ladizinsky, G.; Saidi, N. Assessment of important technological parameters of new Moroccan domesticated tetraploid oat lines of Avena magna. Proc. Appl. Bot. Genet. Breed. 2018, 179, 32–42. [Google Scholar] [CrossRef]
- Horwitz, W.; Latimer, G.W. AOAC Official Method 990.03, Protein (Crude) in Animal Feed, Combustion Method. In Official Methods of Analysis of AOAC International; AOAC: Gaithersurg, MD, USA, 2006; pp. 30–31. [Google Scholar]
- AOAC. Official Method 922.06: Fat in Flour: Acid hydrolysis Method. In Official Methods of Analysis of AOAC International, 19th ed.; AOAC International: Gaithersburg, MD, USA, 2012. [Google Scholar]
- American Association of Cereal Chemists. Method 40-75.01: Determination of Minerals by Inductively Coupled Plasma Spectroscopy. In Approved Methods of the AACC, 10th ed.; American Association of Cereal Chemists: St. Paul, MN, USA, 2001. [Google Scholar]
- AOAC. Official Method 995.16 β-D-Glucan in Barley and Oats Streamlined Enzymatic Method: First Action. In Official Methods of Analysis of AOAC International; AOAC: Gaithersburg, MD, USA, 1995. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org (accessed on 15 September 2022).
- Husson, F.; Le, S.; Pagès, J. Exploratory Multivariate Analysis by Example Using R; Chapman and Hall: New York, NY, USA, 2010. [Google Scholar]
- Purchase, J.L. Parametric Analysis to Describe Genotype × Environment Interaction and Yield Stability in Winter Wheat. Ph.D. Thesis, University of Free State, Bloemfontein, South Africa, 1997. [Google Scholar]
- Mut, Z.; Akay, H.; Köse, E.D.Ö. Grain yield, quality traits and grain yield stability of local oat cultivars. J. Soil. Sci. Plant Nutr. 2018, 18, 269–281. [Google Scholar]
- Nelson, S.O.; Kraszewski, A.W.; Trabelsi, S.; Lawrence, K.C. Using cereal grain permittivity for sensing moisture content. IEEE Trans. Instrum. Meas. 2000, 49, 470–475. [Google Scholar]
- Koehler, P.; Wieser, H. Chemistry of Cereal Grains. In Handbook on Sourdough Biotechnology; Springer Science + Business Media: New York, NY, USA, 2013; pp. 11–45. [Google Scholar] [CrossRef]
- Saidi, N.; Saidi, S.; Hilali, A.; Benchekroun, M.; Al Faiz, C.; Bouksaim, M.; Shaimi, N.; Souihka, A.; Salih Idrissi, A.; Gaboune, F.; et al. Improvement of oat hexaploid lines’s groat nutritive value via hybridisation with tetraploid oat A. magna. Am. J. Res. Commun. 2013, 9, 126–135. [Google Scholar]
- Ladizinsky, G. Studies in Oat Evolution: A Man’s Life with Avena; Springer: Berlin/Heidelberg, Germany; New York, NY, USA; Dordrecht, The Netherlands; London, UK, 2012; ISBN 978-3-642-30547-4. [Google Scholar] [CrossRef]
- Arendt, E.K.; Zannini, E. Oats. In Cereal Grains for the Food and Beverage Industries; Woodhead Publishing: Cambridge, UK, 2013; Volume 1, pp. 243–283e. [Google Scholar] [CrossRef]
- Zhou, M.; Robards, K.; Glennie-Holmes, M.; Helliwell, S. Structure and pasting properties of oat starch. Cereal Chem. 1998, 75, 273–281. [Google Scholar] [CrossRef]
- Blackwood, A.D.; Salter, J.; Dettmar, P.W.; Chaplin, M.F. Dietary fibre, physicochemical properties and their relationship to health. J. R. Soc. Promot. Health 2000, 120, 242–247. [Google Scholar] [CrossRef]
- Babio, N.; Balanza, R.; Basulto, J.; Bullo, M.; Salas-Salvado, J. Dietary fibre: Influence on body weight, glycemic control and plasma cholesterol profile. Nutr. Hosp. 2010, 25, 327–340. [Google Scholar]
- Biel, W.; Kazimierska, K.; Bashutska, U. Nutritional value of wheat, triticale, barley and oat grains. Acta Sci. Pol. Zootech. 2020, 19, 19–28. [Google Scholar] [CrossRef]
- Lazaridou, A.; Biliaderis, C.G.; Izydorczyk, M.S. Cereal beta-glucans: Structures, physical properties, and physiological functions. In Functional Food Carbohydrates; CRC Press: Boca Raton, FL, USA, 2007; pp. 1–72. [Google Scholar]
- Zambrano, J.A.; Thyagarajan, A.; Sardari, R.R.; Olsson, O. Characterization of high Arabinoxylan oat lines identified from a mutagenized oat population. Food Chem. 2023, 404, 134687. [Google Scholar] [CrossRef]
- Redaelli, R.; Scalfati, G.; Ciccoritti, R.; Cacciatori, P.; De Stefanis, E.; Sgrulletta, D. Effects of Genetic and Agronomic Factors on Grain Composition in Oats. Cereal Res. Commun. 2015, 43, 144–154. [Google Scholar] [CrossRef]
- De Oliveira Maximino, J.V.; Barros, L.M.; Pereira, R.M.; de Santi, I.I.; Aranha, B.C.; Busanello, C.; Viana, V.E.; Freitag, R.A.; Batista, B.L.; de Oliveira, A.C.; et al. Mineral and Fatty Acid Content Variation in White Oat Genotypes Grown in Brazil. Biol. Trace Elem. Res. 2020, 199, 1194–1206. [Google Scholar] [CrossRef]
- Garutti, M.; Nevola, G.; Mazzeo, R.; Cucciniello, L.; Totaro, F.; Bertuzzi, C.A.; Caccialanza, R.; Pedrazzoli, P.; Puglisi, F. The Impact of Cereal Grain Composition on the Health and Disease Outcomes. Front. Nutr. 2022, 9, 888974. [Google Scholar] [CrossRef] [PubMed]
- Wessells, K.R.; Singh, G.M.; Brown, K.H. Estimating the Global Prevalence of Inadequate Zinc Intake from National Food Balance Sheets: Effects of Methodological Assumptions. PLoS ONE 2012, 7, e50565. [Google Scholar] [CrossRef]
- Kassebaum, N.J.; Jasrasaria, R.; Naghavi, M.; Wulf, S.K.; Johns, N.; Lozano, R.; Regan, M.; Weatherall, D.; Chou, D.P.; Eisele, T.P.; et al. A systematic analysis of global anemia burden from 1990 to 2010. Blood 2014, 123, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Doehlert, D.C.; Simsek, S.; Thavarajah, D.; Thavarajah, P.; Ohm, J. Detailed Composition Analyses of Diverse Oat Genotype Kernels Grown in Different Environments in North Dakota. Cereal Chem. 2013, 90, 572–578. [Google Scholar] [CrossRef]
- Benlhabib, O.; Yazar, A.; Qadir, M.; Lourenço, E.; Jacobsen, S.E. How can we improve Mediterranean cropping systems? J. Agro. Crop Sci. 2014, 200, 325–332. [Google Scholar] [CrossRef]
- Carvalho, F.I.F.; Lorencetti, C.; Benin, G. Estimates of the Correlation and Implications in Plant Breeding; UFPel: Pelotas, Brazil, 2004; 142p. [Google Scholar]
- Lorencetti, C.; Carvalho, F.I.F.; Oliveira, A.C.; Valério, I.P.; Hartwig, I.; Benin, G.; Schmidt, D.A.M. Aplicability of phenotypic and canonic correlations and path coefficients in the selection of oat genotypes. Sci. Agric. 2006, 63, 11–19. [Google Scholar] [CrossRef]
- Hawerroth, M.C.; de Carvalho, F.I.F.; de Oliveira, A.C.; da Silva, J.A.G.; Gutkoski, L.C.; Sartori, J.F.; Woyann, L.G.; Barbieri, R.L.; Hawerroth, F.J. Adaptability and stability of white oat cultivars in relation to chemical composition of the caryopsis. Pesqui. Agropecuária Bras. 2013, 48, 42–50. [Google Scholar] [CrossRef]
- Holland, J.B. Oat improvement. In Crop Improvement for the 21st Century; Kang, M.S., Ed.; Research Signpost: Trivandrum, India, 1997; Chapter 4; pp. 57–98. [Google Scholar]
- Peterson, D.M.; Wesenberg, D.M.; Burrup, D.E.; Erickson, C.A. Relationships among agronomic traits and grain composition in oat genotypes grown in different environments. Crop Sci. 2005, 45, 1249–1255. [Google Scholar] [CrossRef]
- Chernyshova, A.A.; White, P.J.; Scott, M.P.; Jannink, J. Selection for nutritional function and agronomic performance in oat. Crop Sci. 2007, 47, 2330–2339. [Google Scholar] [CrossRef]
- Marinissen, J.; Arelovich, H.M.; Martínez, M.F.; Zilio, L.; Ombrosi, D. Oats grain supplementation of calves grazing oats pasture. 1. Diet composition, live weight gain and blood parameters. Rev. Arg. Prod. Anim. 2004, 24, 80–81. [Google Scholar]
- Martinez, M.; Arelovich, H.; Wehrhahne, L. Grain yield, nutrient content and lipid profile of oat genotypes grown in a semiarid environment. Field Crops Res. 2010, 116, 92–100. [Google Scholar] [CrossRef]
- Hizbai, B.T.; Gardner, K.M.; Wight, C.P.; Dhanda, R.K.; Molnar, S.J.; Johnson, D.; Frégeau-Reid, J.; Yan, W.; Rossnagel, B.G.; Holland, J.B.; et al. Quantitative trait loci affecting oil content, oil composition, and other agronomically important traits in oat. Plant Genome 2012, 5, 164–175. [Google Scholar] [CrossRef]
- Silva, C.F.L.; Milach, S.C.K.; Silva, S.D.A.; Montero, C.R. Near infrared reflectance spectroscopy (NIRS) to assess protein and lipid contents in Avena sativa L. Crop Breed. Appl. Biotechnol. 2008, 8, 127–133. [Google Scholar]
- Rasane, P.; Jha, A.; Sabikhi, L.; Kumar, A.; Unnikrishnan, V.S. Nutritional advantages of oats and opportunities for its processing as value added foods-a review. J. Food Sci. Technol. 2015, 52, 662–675. [Google Scholar] [PubMed]
- Sinkovič, L.; Rakszegi, M.; Pipan, B.; Meglič, V. Compositional Traits of Grains and Groats of Barley, Oat and Spelt Grown at Organic and Conventional Fields. Foods 2023, 12, 1054. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.C. Role of calcium in micronutrient stresses of plants. Commun. Soil Sci. Plant Anal. 1979, 10, 459–472. [Google Scholar] [CrossRef]
- Thor, K. Calcium—Nutrient and Messenger. Front. Plant Sci. 2019, 10, 440. [Google Scholar] [CrossRef]
- Saleem, M.H.; Usman, K.; Rizwan, M.; Al Jabri, H.; Alsafran, M. Functions and strategies for enhancing zinc availability in plants for sustainable agriculture. Front. Plant Sci. 2022, 13, 1033092. [Google Scholar] [CrossRef]
- Mousia, Z.; Edherly, S.; Pandiella, S.S.; Webb, C. Effect of wheat pearling on flour quality. Food Res. Int. 2004, 37, 449–459. [Google Scholar]
- De Brier, N.; Gomand, S.V.; Donner, E.; Paterson, D.; Delcour, J.A.; Lombi, E.; Smolders, E. Distribution of Minerals in Wheat Grains (Triticum aestivum L.) and in Roller Milling Fractions Affected by Pearling. J. Agric. Food Chem. 2015, 63, 1276–1285. [Google Scholar] [CrossRef] [PubMed]
- Jordan-Meille, L.; Holland, J.; McGrath, S.; Glendining, M.; Thomas, C.; Haefele, S. The grain mineral composition of barley, oat and wheat on soils with pH and soil phosphorus gradients. Eur. J. Agron. 2021, 126, 126281. [Google Scholar] [CrossRef]
- Raboy, V. Seeds for a better future: ‘low phytate’ grains help to overcome malnutrition and reduce pollution. Trends Plant Sci. 2001, 6, 458–462. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, U.; Cummins, E. Simulation of the factors affecting β-glucan levels during the cultivation of oats. J. Cereal Sci. 2009, 50, 175–183. [Google Scholar] [CrossRef]
- Zhou, M.X.; Glennie Holmes, M.; Robards, K.; Helliwell, S. Fatty Acid Composition of Lipids of Australian Oats. J. Cereal Sci. 1998, 28, 311–319. [Google Scholar] [CrossRef]
- Banaś, A.; Dębski, H.; Banaś, W.; Heneen, W.K.; Dahlqvist, A.; Bafor, M.; Gummeson, P.-O.; Marttila, S.; Ekman, Å.; Carlsson, A.S.; et al. Lipids in grain tissues of oat (Avena sativa): Differences in content, time of deposition, and fatty acid composition. J. Exp. Bot. 2007, 58, 2463–2470. [Google Scholar] [CrossRef]
- Ali, Q.; Ashraf, M.; Anwar, F. Seed Composition and Seed Oil Antioxidant Activity of Maize Under Water Stress. J. Am. Oil Chem. Soc. 2010, 87, 1179–1187. [Google Scholar] [CrossRef]
- Ullah, S.; Khan, M.N.; Lodhi, S.S.; Ahmed, I.; Tayyab, M.; Mehmood, T.; Din, I.U.; Khan, M.; Sohail, Q.; Akram, M. Targeted metabolomics reveals fatty acid abundance adjustments as playing a crucial role in drought-stress response and post-drought recovery in wheat. Front. Genet. 2022, 13, 972696. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.; Wilckens, R.; Jorge, J.A.R.A.; Aranda, M. Controlled water stress to improve functional and nutritional quality in quinoa seed. Boletín Latinoam. Caribe Plantas Med. Aromáticas 2013, 12, 457–468. [Google Scholar]



{kind=link}
{kind=link}
| Experimental Site | Zone | Coordinates | Alt | Soil Type | Temp | Rain | Hum | S.H | Days to Harvest | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Latitude | Longitude | Mn | Mx | ||||||||
| Alnif | Oasis | 31°06′37″ N | 5°03′50″ W | 1320 | Fluvisols | 9 | 43 | 43.2 * | 26.2 | 3529 | 177 |
| Bouchane | Phosphate Plateau | 32°14′35″ N | 8°19′45″ W | 830 | Cambisols | 9 | 38 | 250.2 | 54.1 | 3304 | 219 |
| El Kbab | Middle Atlas | 32°42′25″ N | 5°31′57″ W | 1540 | Fluvisols | 4 | 38 | 573.0 | 50.9 | 3477 | 206 |
| Line/Site | Protein | Moisture | Fat | Fiber | β-Glucan | Ash | Starch |
|---|---|---|---|---|---|---|---|
| ATC | 22.4 f ± 1.40 | 12.9 a ± 0.14 | 8.31 b ± 0.42 | 6.77 de ± 0.51 | 2.49 a ± 0.07 | 2.49 b ± 0.03 | 57.0 b ± 1.28 |
| AT1 | 21.8 g ± 1.82 | 12.7 ab ± 0.15 | 8.01 c ± 1.05 | 6.62 e ± 0.69 | 2.77 a ± 0.20 | 2.60 ab ± 0.05 | 58.2 a ± 1.53 |
| AT2 | 22.0 fg ± 1.90 | 12.8 ab ± 0.36 | 8.31 b ± 1.26 | 7.51 b ± 0.96 | 2.74 a ± 0.38 | 2.5 b ± 0.19 | 58.1 a ± 1.13 |
| AT3 | 23.6 c ± 2.80 | 12.8 ab ± 0.51 | 8.35 b ± 1.41 | 7.40 bc ± 1.44 | 2.69 a ± 0.43 | 2.72 a ± 0.25 | 55.7 c ± 0.95 |
| AT4 | 23.2 d ± 2.23 | 12.7 b ± 0.18 | 7.92 c ± 1.62 | 6.87 de ± 1.09 | 2.34 a ± 0.15 | 2.51 b ± 0.09 | 57.4 ab ± 1.02 |
| AT5 | 24.2 b ± 2.44 | 12.6 b ± 0.18 | 8.31 b ± 1.10 | 7.41 bc ± 0.71 | 2.42 a ± 0.11 | 2.59 ab ± 0.15 | 57.0 b ± 1.04 |
| AT6 | 25.4 a ± 2.05 | 12.6 b ± 0.22 | 8.28 b ± 1.26 | 8.00 a ± 1.06 | 2.60 a ± 0.05 | 2.58 ab ± 0.22 | 54.9 d ± 0.45 |
| AT7 | 22.0 fg ± 2.14 | 12.9 a ± 0.23 | 8.65 a ± 1.21 | 7.07 cd ± 0.62 | 2.42 a ± 0.19 | 2.44 b ± 0.09 | 58.1 a ± 0.70 |
| AT9 | 22.8 e ± 1.45 | 12.8 ab ± 0.28 | 8.63 a ± 1.05 | 6.85 de ± 0.81 | 2.45 a ± 0.26 | 2.52 b ± 0.11 | 57.7 ab ± 0.67 |
| AT13 | 23.3 d ± 2.63 | 12.7 ab ± 0.21 | 8.69 a ± 0.79 | 7.96 a ± 1.50 | 2.63 a ± 0.39 | 2.62 ab ± 0.10 | 57.1 b ± 0.91 |
| AT14 | 22.0 fg ± 0.53 | 12.8 ab ± 0.32 | 8.78 a ± 1.17 | 6.88 de ± 0.74 | 2.20 a ± 0.02 | 2.42 b ± 0.17 | 58.0 a ± 1.11 |
| AT15 | 23.7 c ± 1.71 | 12.7 ab ± 0.28 | 8.37 b ± 1.36 | 7.38 bc ± 1.20 | 2.64 a ± 0.21 | 2.56 ab ± 0.16 | 57.5 ab ± 0.73 |
| Alnif | 24.1 a ± 2.05 | 12.8 a ± 0.31 | 7.56 c ± 0.60 | 6.84 b ± 0.57 | 2.52 a ± 0.29 | 2.57 a ± 0.16 | 57.2 b ± 1.22 |
| Bouchane | 20.9 b ± 1.11 | 12.6 b ± 0.20 | 9.78 a ± 0.40 | 8.35 a ± 0.84 | 2.61 a ± 0.22 | 2.49 b ± 0.12 | 57.7 a ± 1.46 |
| El Kbab | 24.2 a ± 1.21 | 12.8 a ± 0.27 | 7.82 b ± 0.48 | 6.49 c ± 0.50 | 2.46 a ± 0.27 | 2.59 a ± 0.19 | 56.9 b ± 1.35 |
| Wheat | Rye | Corn | Barley | Rice | Millet | A. sativa | A. magna * | |
|---|---|---|---|---|---|---|---|---|
| Moisture | 12.6 | 13.6 | 11.3 | 12.1 | 13.0 | 12.0 | 13.1 | 12.8 (12.3–13.6) |
| Protein (N × 6.25) | 11.3 | 9.4 | 8.8 | 11.1 | 7.7 | 10.5 | 10.8 | 23.1 (19.4–27.4) |
| Lipids | 1.8 | 1.7 | 3.8 | 2.1 | 2.2 | 3.9 | 7.2 | 8.4 (6.4–10.3) |
| Available carbohydrates | 59.4 | 60.3 | 65.0 | 62.7 | 73.7 | 68.2 | 56.2 | 57.2 ** (54.0–61.1) |
| Fiber | 13.2 | 13.1 | 9.8 | 9.7 | 2.2 | 3.8 | 9.8 | 7.2 (5.6–10.2) |
| Minerals | 1.7 | 1.9 | 1.3 | 2.3 | 1.2 | 1.6 | 2.9 | 2.6 (2.1–3.1) |
| Macroelements (×10−1 g/100 g DM Basis) | Microelements (×10−3 g/100 g DM Basis) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Line/Site | Na | K | Ca | P | S | Mg | Fe | Zn | Cu | Mn |
| ATC | 0.03 b ± 0.00 | 3.65 a ± 0.04 | 0.65 c ± 0.01 | 4.65 ab ± 0.05 | 2.08 c ± 0.02 | 1.44 c ± 0.02 | 6.80 a ± 0.97 | 3.30 ab ± 0.66 | 0.55 ab ± 0.16 | 4.26 cd ± 0.58 |
| AT1 | 0.03 b ± 0.01 | 3.38 ab ± 0.02 | 0.71 abc ± 0.01 | 4.52 ab ± 0.02 | 2.12 c ± 0.02 | 1.46 bc ± 0.01 | 8.22 a ± 2.57 | 3.27 ab ± 0.41 | 0.57 ab ± 0.15 | 4.46 abcd ± 0.14 |
| AT2 | 0.03 ab ± 0.00 | 3.55 ab ± 0.05 | 0.77 a ± 0.01 | 4.66 ab ± 0.04 | 2.23 abc ± 0.01 | 1.60 ab ± 0.01 | 9.33 a ± 3.10 | 3.16 b ± 0.30 | 0.59 a ± 0.13 | 4.80 ab ± 0.07 |
| AT3 | 0.04 a ± 0.00 | 3.68 a ± 0.05 | 0.68 bc ± 0.01 | 4.67 ab ± 0.06 | 2.23 abc ± 0.02 | 1.53 abc ± 0.01 | 7.70 a ± 1.57 | 3.32 ab ± 0.61 | 0.55 ab ± 0.13 | 4.61 abcd ± 0.50 |
| AT4 | 0.04 ab ± 0.00 | 3.48 ab ± 0.07 | 0.69 abc ± 0.01 | 4.53 ab ± 0.07 | 2.32 ab ± 0.01 | 1.52 abc ± 0.02 | 8.04 a ± 2.59 | 3.32 ab ± 0.50 | 0.49 b ± 0.15 | 4.34 bcd ± 0.28 |
| AT5 | 0.04 a ± 0.00 | 3.03 b ± 0.04 | 0.72 abc ± 0.01 | 4.26 b ± 0.06 | 2.22 abc ± 0.02 | 1.48 bc ± 0.01 | 7.60 a ± 1.92 | 3.30 ab ± 0.48 | 0.49 b ± 0.05 | 4.23 d ± 0.29 |
| AT6 | 0.03 b ± 0.00 | 3.67 a ± 0.06 | 0.74 ab ± 0.01 | 4.76 a ± 0.09 | 2.35 a ± 0.02 | 1.57 abc ± 0.01 | 8.00 a ± 1.74 | 3.52 ab ± 0.65 | 0.58 a ± 0.20 | 4.50 abcd ± 0.64 |
| AT7 | 0.03 ab ± 0.00 | 3.45 ab ± 0.03 | 0.72 abc ± 0.01 | 4.61 ab ± 0.06 | 2.24 abc ± 0.02 | 1.52 abc ± 0.01 | 7.18 a ± 0.83 | 3.63 a ± 0.79 | 0.50 b ± 0.10 | 4.79 ab ± 0.61 |
| AT9 | 0.03 ab ± 0.00 | 3.45 ab ± 0.04 | 0.70 abc ± 0.01 | 4.54 ab ± 0.04 | 2.20 abc ± 0.02 | 1.53 abc ± 0.01 | 7.24 a ± 1.22 | 3.45 ab ± 0.68 | 0.53 ab ± 0.10 | 4.70 abc ± 0.43 |
| AT13 | 0.03 b ± 0.00 | 3.55 ab ± 0.06 | 0.74 ab ± 0.01 | 4.51 ab ± 0.05 | 2.40 a ± 0.02 | 1.60 ab ± 0.00 | 6.53 a ± 0.66 | 3.70 a ± 0.69 | 0.57 ab ± 0.11 | 4.49 abcd ± 0.23 |
| AT14 | 0.04 ab ± 0.00 | 3.62 a ± 0.05 | 0.65 c ± 0.01 | 4.70 a ± 0.04 | 2.25 abc ± 0.01 | 1.63 a ± 0.01 | 7.10 a ± 1.05 | 3.66 a ± 0.57 | 0.56 ab ± 0.08 | 4.90 a ± 0.45 |
| AT15 | 0.04 ab ± 0.00 | 3.68 a ± 0.05 | 0.70 abc ± 0.01 | 4.57 ab ± 0.05 | 2.26 abc ± 0.01 | 1.49 bc ± 0.02 | 6.42 a ± 1.04 | 3.68 a ± 0.84 | 0.55 ab ± 0.09 | 4.40 bcd ± 0.51 |
| Alnif | 0.03 b ± 0.00 | 4.01 a ± 0.03 | 0.64 b ± 0.01 | 5.09 a ± 0.03 | 2.27 a ± 0.01 | 1.64 a ± 0.01 | 8.00 a ± 2.34 | 3.70 b ± 0.25 | 0.67 a ± 0.10 | 4.80 a ± 0.54 |
| Bouchane | 0.04 a ± 0.00 | 3.24 b ± 0.02 | 0.84 a ± 0.01 | 4.09 c ± 0.28 | 2.12 b ± 0.01 | 1.46 b ± 0.01 | 6.47 b ± 0.80 | 2.80 c ± 0.16 | 0.47 b ± 0.04 | 4.39 b ± 0.41 |
| El Kbab | 0.03 b ± 0.00 | 3.29 b ± 0.03 | 0.65 b ± 0.01 | 4.57 b ± 0.04 | 2.34 a ± 0.02 | 1.49 b ± 0.01 | 8.06 a ± 1.51 | 3.86 a ± 0.54 | 0.50 b ± 0.10 | 4.40 b ± 0.23 |
| Wheat | Rye | Corn | Barley | Rice | Millet | A. sativa | A. magna * | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | Mean | Min | Max | |||||||
| Sodium | 0.02 | 0.02 | 0.35 | 0.12 | 0.05 | 0.05 | 0.02 | 0.03 | 0.02 | 0.04 |
| Potassium | 4.94 | 5.10 | 2.87 | 4.52 | 0.92 | 1.95 | 4.29 | 3.53 | 2.70 | 4.70 |
| Calcium | 0.30 | 0.24 | 0.15 | 0.33 | 0.24 | 0.08 | 0.54 | 0.70 | 0.50 | 1.00 |
| Phosphorous | 3.04 | 3.32 | 2.56 | 2.64 | 0.94 | 2.85 | 5.23 | 4.57 | 3.50 | 5.80 |
| Magnesium | 1.60 | 1.10 | 1.20 | 1.33 | 0.20 | 1.14 | 1.77 | 1.53 | 1.30 | 1.80 |
| Iron | 3.30 | 2.60 | 2.40 | 3.60 | 0.80 | 3.00 | 4.70 | 7.50 | 4.50 | 14.0 |
| Zinc | 2.90 | 2.70 | 2.20 | 2.80 | 1.30 | 1.70 | 4.00 | 3.40 | 2.50 | 4.80 |
| Copper | 0.40 | 0.40 | 0.30 | 0.50 | 0.20 | 0.80 | 0.60 | 0.50 | 0.30 | 0.90 |
| Manganese | 4.00 | 2.60 | 0.50 | 1.90 | 1.10 | 1.60 | 4.90 | 4.50 | 3.50 | 5.80 |
| Fat | Fiber | β-Glucan | Starch | Ash | Na | K | Ca | P | S | Mg | Fe | Zn | Cu | Mn | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Protein | −0.764 ** | −0.379 * | −0.324 | −0.517 ** | 0.451 ** | −0.141 | 0.349 * | −0.636 ** | 0.549 ** | 0.712 ** | 0.276 | 0.324 | 0.718 ** | 0.357 * | −0.041 |
| Fat | 1 | 0.686 ** | 0.277 | 0.227 | −0.326 | 0.311 | −0.532 ** | 0.801 ** | −0.733 ** | −0.539 ** | −0.403 * | −0.583 ** | −0.738 ** | −0.515 ** | −0.132 |
| Fiber | 1 | 0.131 | −0.048 | −0.153 | 0.159 | −0.240 | 0.764 ** | −0.513 ** | −0.331 * | −0.191 | −0.407 * | −0.593 ** | −0.277 | −0.113 | |
| Ash | 1 | −0.121 | 0.243 | 0.059 | 0.023 | 0.397 * | −0.100 | −0.313 | −0.068 | −0.119 | −0.376 * | −0.056 | −0.102 | ||
| Starch | 1 | −0.413 * | 0.135 | −0.105 | 0.180 | −0.146 | −0.237 | −0.059 | −0.234 | −0.110 | −0.054 | 0.194 | |||
| β-glucan | 1 | −0.074 | 0.097 | −0.311 | 0.215 | 0.253 | 0.075 | 0.174 | 0.302 | 0.143 | −0.170 | ||||
| Na | 1 | −0.152 | 0.297 | −0.233 | −0.231 | −0.204 | −0.386 * | −0.235 | −0.263 | −0.189 | |||||
| K | 1 | −0.378 * | 0.883 ** | 0.206 | 0.763 ** | 0.233 | 0.408 * | 0.720 ** | 0.382 * | ||||||
| Ca | 1 | −0.556 ** | −0.442 ** | −0.312 | −0.411 * | −0.690 ** | −0.471 ** | −0.198 | |||||||
| P | 1 | 0.431 ** | 0.756 ** | 0.342 * | 0.679 ** | 0.606 ** | 0.338 * | ||||||||
| S | 1 | 0.381 * | 0.277 | 0.745 ** | 0.253 | 0.135 | |||||||||
| Mg | 1 | 0.334 * | 0.418 * | 0.633 ** | 0.581 ** | ||||||||||
| Fe | 1 | 0.242 | 0.480 ** | 0.218 | |||||||||||
| Zn | 1 | 0.360 * | 0.156 | ||||||||||||
| Cu | 1 | 0.472 ** |
| Protein | Fat | Fiber | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Source | Df | Sum Sq | F Value | Var a | Sum Sq | F Value | Var a | Sum Sq | F Value | Var a |
| Site (E) | 2 | 244.07 | 502.67 *** | 51% | 105.74 | 1722.84 *** | 81% | 70.52 | 496.95 *** | 65% |
| Rep (Site) | 6 | 1.46 | 13.39 | - | 0.18 | 5.15 | - | 0.43 | 0.84 | - |
| Line (G) | 11 | 117.79 | 590.46 *** | 25% | 6.88 | 105.06 *** | 5% | 20.67 | 22.34 *** | 19% |
| G × E | 22 | 116.84 | 292.85 *** | 24% | 18.35 | 140.10 *** | 14% | 18.12 | 9.79 *** | 17% |
| IPCA1 | 12 | 90.05 | 413.77 *** | 77% | 11.19 | 156.68 *** | 61% | 11.87 | 11.76 *** | 66% |
| IPCA2 | 10 | 26.79 | 147.75 ** | 23% | 7.16 | 120.20 *** | 39% | 6.24 | 7.42 *** | 35% |
| Residuals | 66 | 1.20 | 0.39 | 5.55 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thiam, E.h.; Dunn, M.; Jackson, E.W.; Jellen, E.N.; Nelson, M.; Rogers, W.; Wallace, C.; Ahlborn, G.; Mounir, M.; Yakovac, T.; et al. Quality Characteristics of Twelve Advanced Lines of Avena magna ssp. domestica Grown in Three Contrasting Locations in Morocco. Plants 2024, 13, 294. https://doi.org/10.3390/plants13020294
Thiam Eh, Dunn M, Jackson EW, Jellen EN, Nelson M, Rogers W, Wallace C, Ahlborn G, Mounir M, Yakovac T, et al. Quality Characteristics of Twelve Advanced Lines of Avena magna ssp. domestica Grown in Three Contrasting Locations in Morocco. Plants. 2024; 13(2):294. https://doi.org/10.3390/plants13020294
Chicago/Turabian StyleThiam, El hadji, Michael Dunn, Eric W. Jackson, Eric N. Jellen, Mark Nelson, Will Rogers, Carol Wallace, Gene Ahlborn, Majid Mounir, Teresa Yakovac, and et al. 2024. "Quality Characteristics of Twelve Advanced Lines of Avena magna ssp. domestica Grown in Three Contrasting Locations in Morocco" Plants 13, no. 2: 294. https://doi.org/10.3390/plants13020294
APA StyleThiam, E. h., Dunn, M., Jackson, E. W., Jellen, E. N., Nelson, M., Rogers, W., Wallace, C., Ahlborn, G., Mounir, M., Yakovac, T., Morris, S., & Benlhabib, O. (2024). Quality Characteristics of Twelve Advanced Lines of Avena magna ssp. domestica Grown in Three Contrasting Locations in Morocco. Plants, 13(2), 294. https://doi.org/10.3390/plants13020294