
Two Growing-Season Warming Partly Promoted Growth but Decreased Reproduction and Ornamental Value of Impatiens oxyanthera


Abstract
:1. Introduction
2. Results
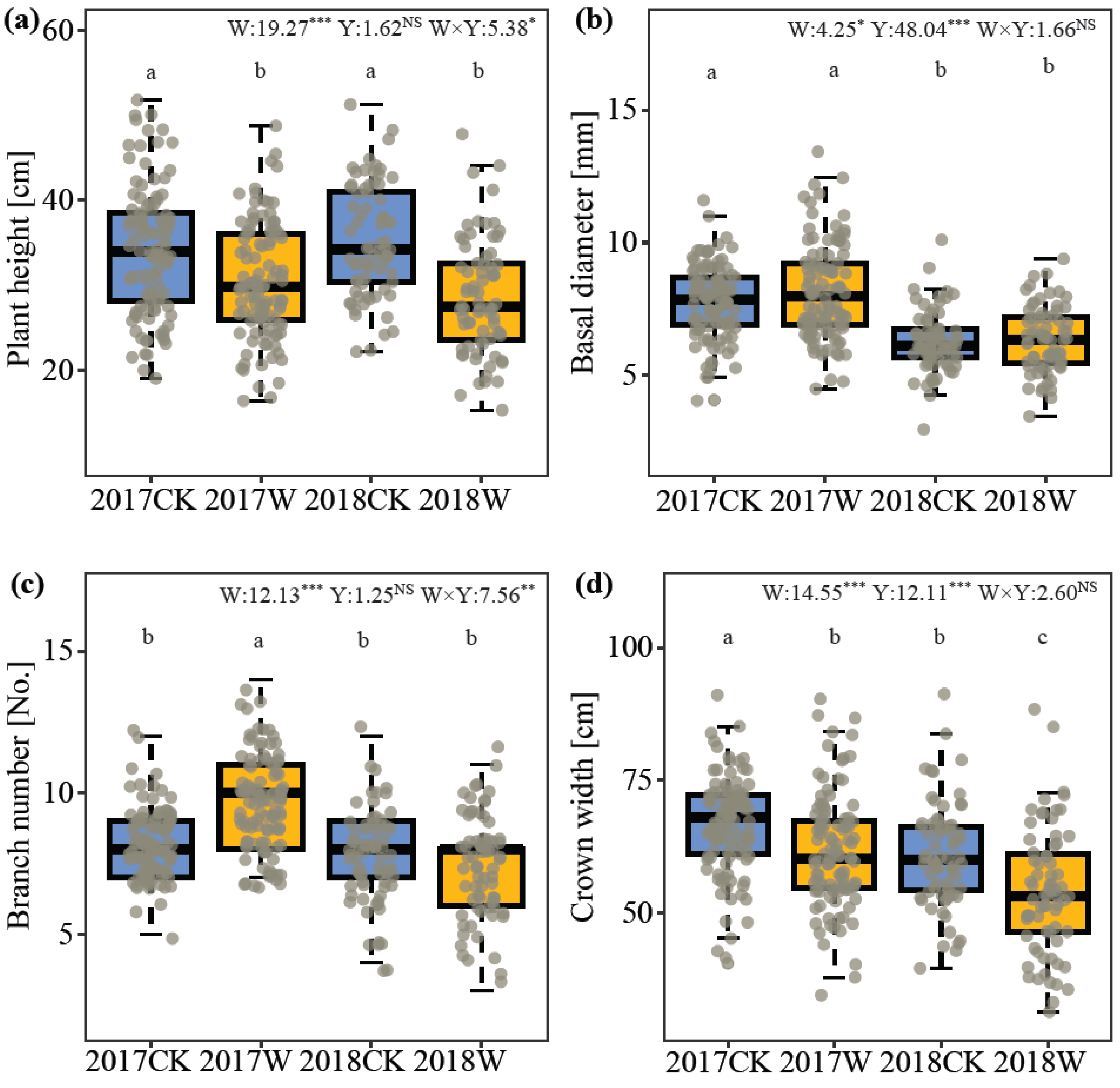
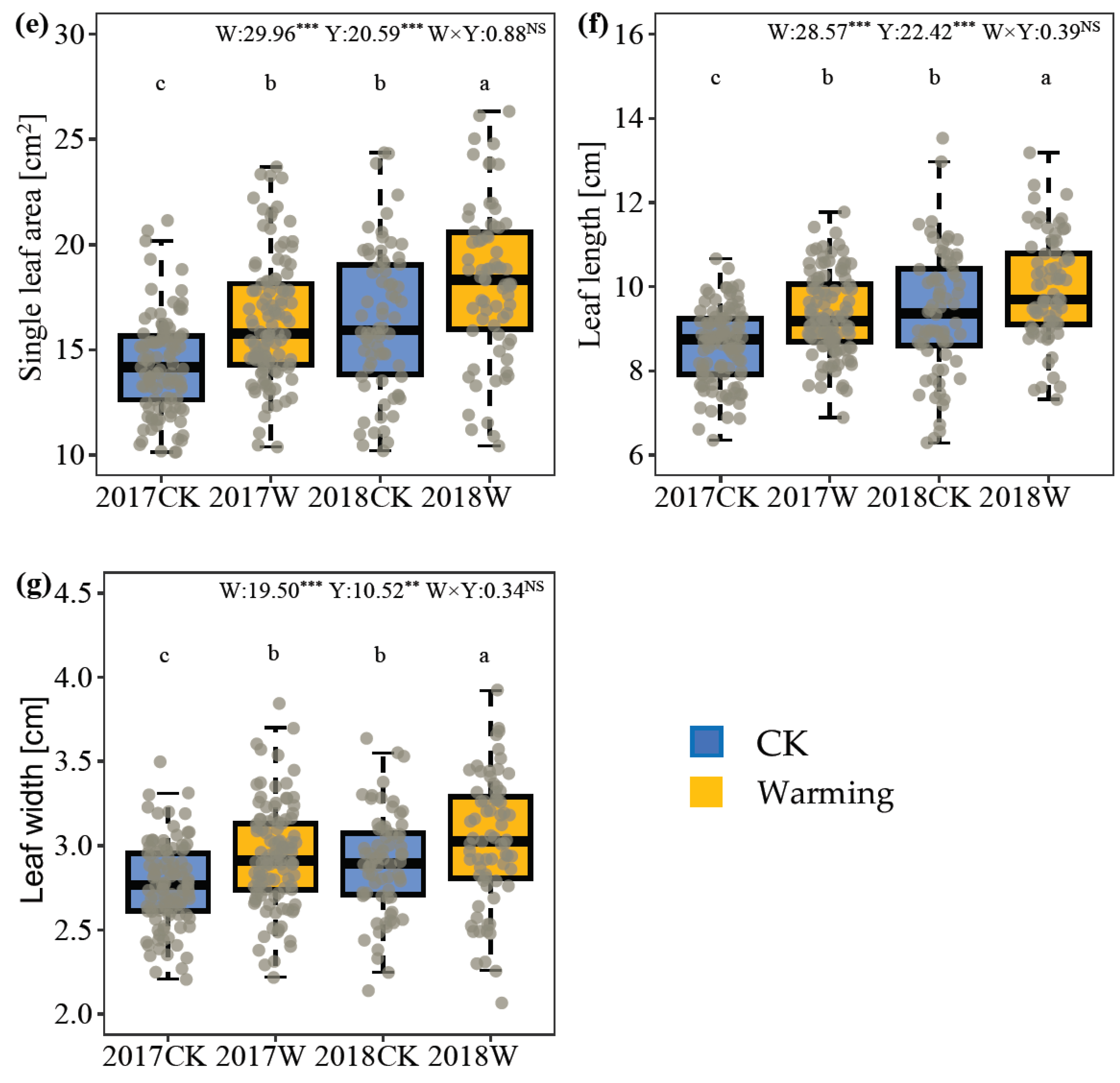
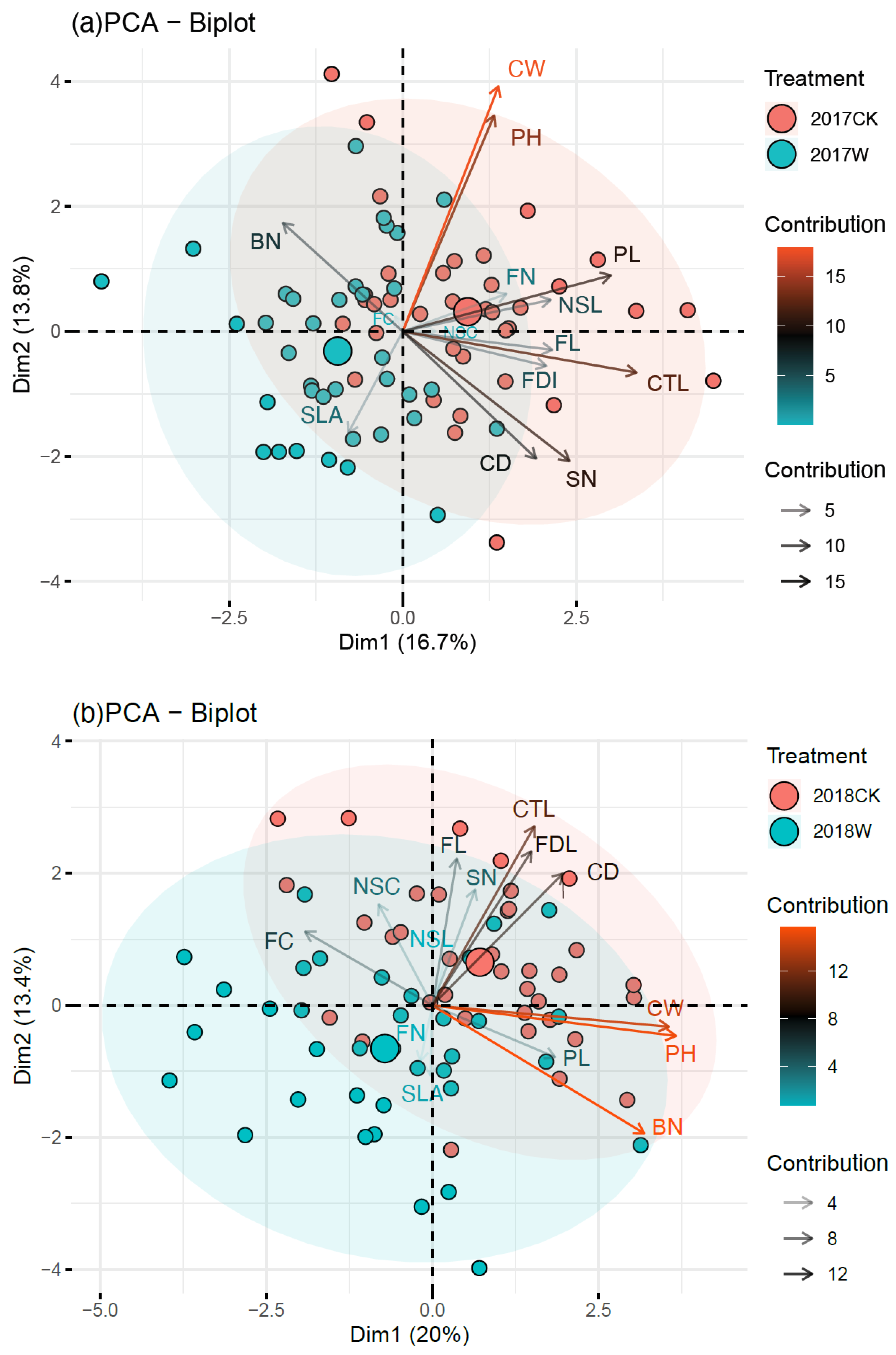
2.1. Plant Vegetative Growth
2.2. Flower Longevity and Flowering Duration
2.3. Floral Traits
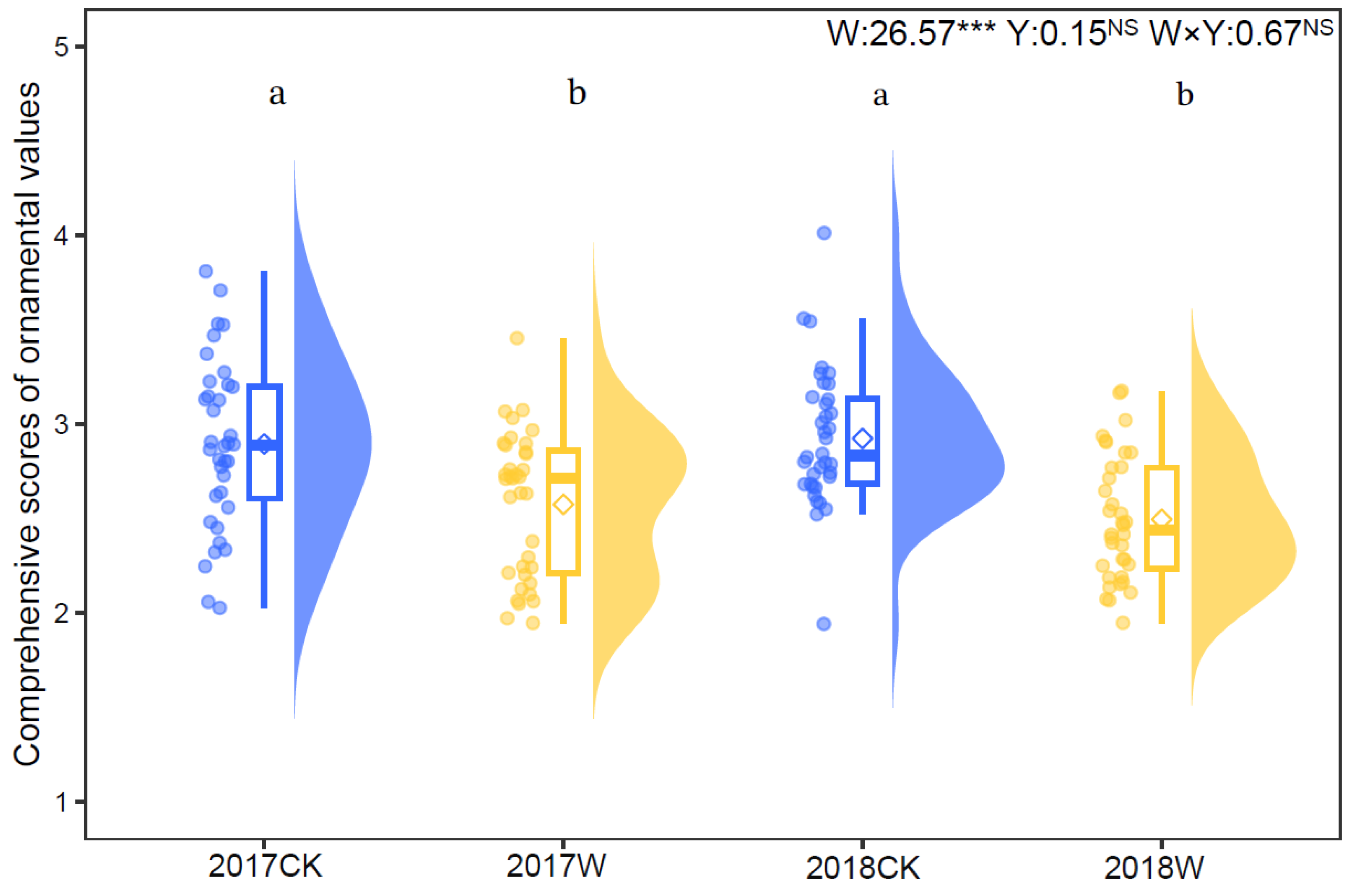
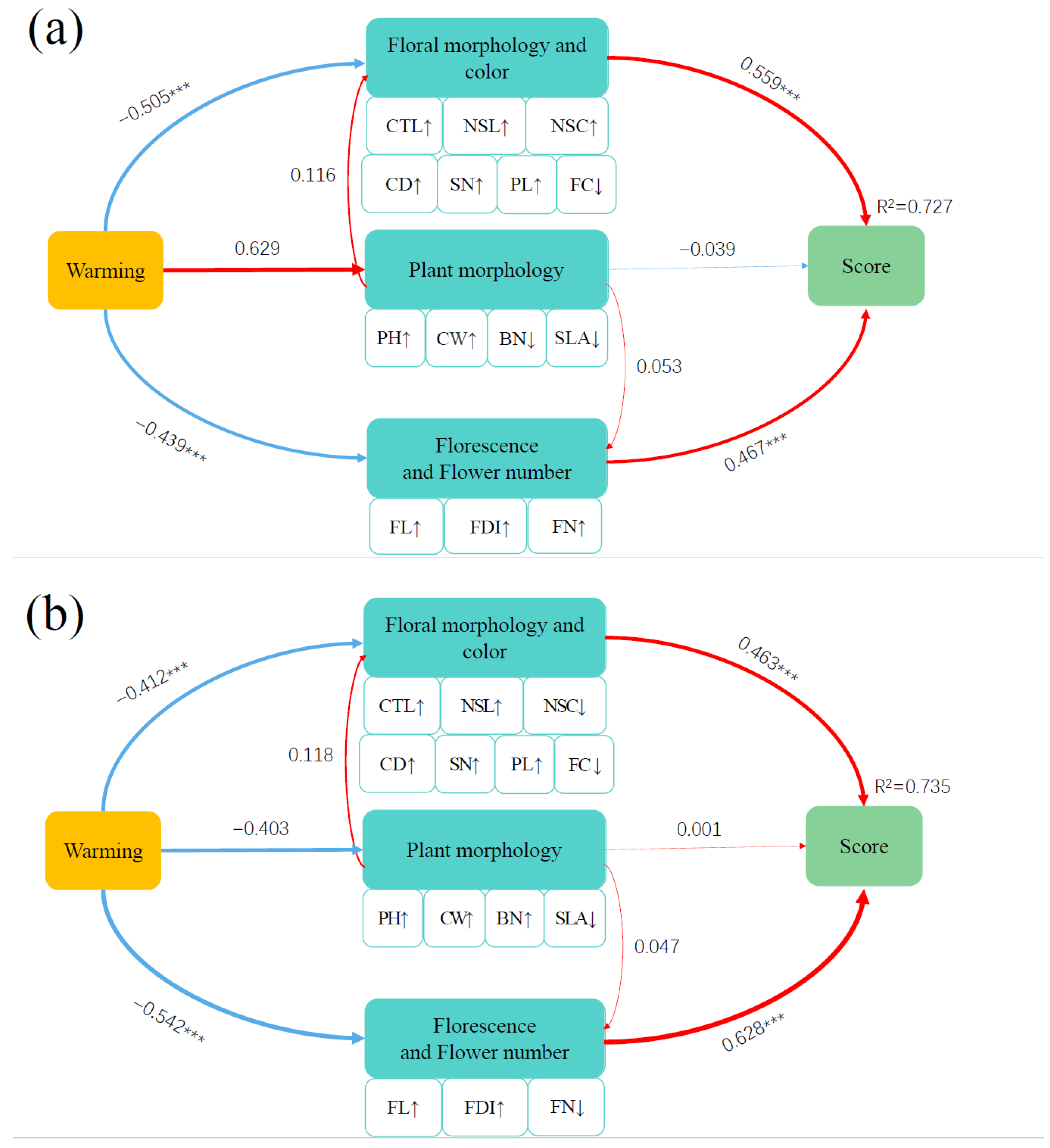
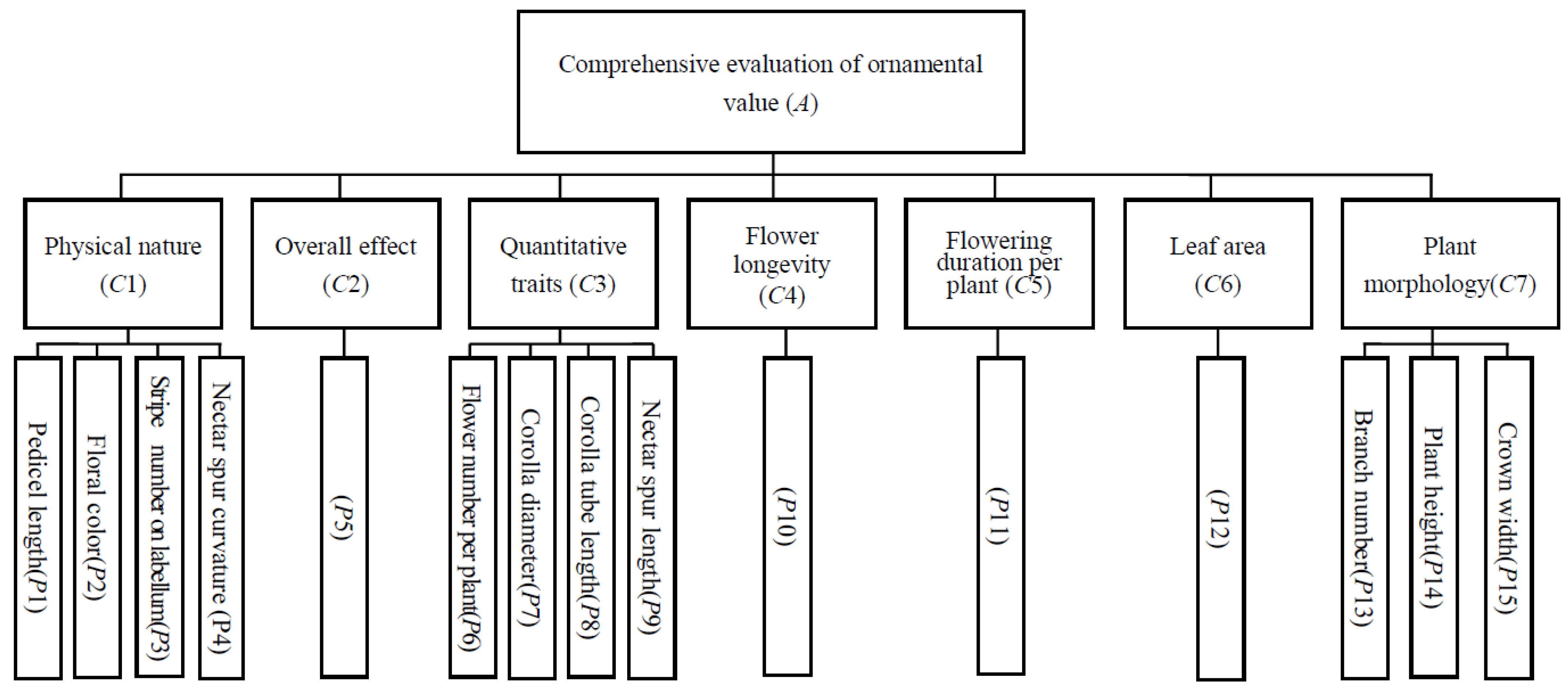
2.4. Comprehensive Ornamental Value
3. Discussion
3.1. Effect of Warming on Plant Vegetative Growth
3.2. Effect of Warming on the Ornamental Time and Flowering Period
3.3. Effect of Warming on Flower Ornamental Characteristics
3.4. Effect of Warming on Comprehensive Ornamental Value
4. Materials and Methods
4.1. Study Site and Plant Materials
4.2. Warming Treatment
4.3. Determination of Air Temperature, Humidity and Soil Temperature
4.4. Measurement of Plant Morphology
4.5. Determination of Ornamental Traits of the Flower
4.6. Determination of Anthocyanin Content
4.7. Evaluation of Ornamental Value
4.8. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pennisi, E. On the origin of flowering plants. Science 2009, 324, 28–31. [Google Scholar] [CrossRef]
- Matthews, H.D.; Wynes, S. Current global efforts are insufficient to limit warming to 1.5 °C. Science 2022, 376, 1404–1409. [Google Scholar] [CrossRef]
- Armstrong McKay, D.I.; Staal, A.; Abrams, J.F.; Winkelmann, R.; Sakschewski, B.; Loriani, S.; Fetzer, I.; Cornell, S.E.; Rockström, J.; Lenton, T.M. Exceeding 1.5 °C global warming could trigger multiple climate tipping points. Science 2022, 377, eabn7950. [Google Scholar] [CrossRef]
- Patil, R.H.; Laegdsmand, M.; Olesen, J.E.; Porter, J.R. Growth and yield response of winter wheat to soil warming and rainfall patterns. J. Agric. Sci. 2010, 148, 553–566. [Google Scholar] [CrossRef]
- Wigge, P.A. Ambient temperature signalling in plants. Curr. Opin. Plant Biol. 2013, 16, 661–666. [Google Scholar] [CrossRef]
- McClung, C.R.; Lou, P.; Hermand, V.; Kim, J.A. The importance of ambient temperature to growth and the induction of flowering. Front. Plant Sci. 2016, 7, 1266. [Google Scholar] [CrossRef] [PubMed]
- Quint, M.; Delker, C.; Franklin, K.A.; Wigge, P.A.; Halliday, K.J.; van Zanten, M. Molecular and genetic control of plant thermomorphogenesis. Nat. Plants 2016, 2, 15190. [Google Scholar] [CrossRef]
- Winkler, M.; Lamprecht, A.; Steinbauer, K.; Hülber, K.; Theurillat, J.; Breiner, F.; Choler, P.; Ertl, S.; Girón, A.G.; Rossi, G.; et al. The rich sides of mountain summits—A pan-European view on aspect preferences of alpine plants. J. Biogeogr. 2016, 43, 2261–2273. [Google Scholar] [CrossRef]
- Kulonen, A.; Imboden, R.A.; Rixen, C.; Maier, S.B.; Wipf, S. Enough space in a warmer world? Microhabitat diversity and small-scale distribution of alpine plants on mountain summits. Divers. Distrib. 2018, 24, 252–261. [Google Scholar] [CrossRef]
- Pereira, H.M.; Leadley, P.W.; Proença, V.; Alkemade, R.; Scharlemann, J.P.W.; Fernandez-Manjarrés, J.F.; Araújo, M.B.; Balvanera, P.; Biggs, R.; Cheung, W.W.L.; et al. Scenarios for Global Biodiversity in the 21st Century. Science 2010, 330, 1496–1501. [Google Scholar] [CrossRef]
- Moritz, C.; Agudo, R. The Future of Species Under Climate Change: Resilience or Decline? Science 2013, 341, 504–508. [Google Scholar] [CrossRef]
- Román-Palacios, C.; Wiens, J.J. Recent responses to climate change reveal the drivers of species extinction and survival. Proc. Natl. Acad. Sci. USA 2020, 117, 4211–4217. [Google Scholar] [CrossRef] [PubMed]
- Urban, M.C. Accelerating extinction risk from climate change. Science 2015, 348, 571–573. [Google Scholar] [CrossRef]
- Warren, R.; Price, J.; Graham, E.; Forstenhaeusler, N.; VanDerWal, J. The projected effect on insects, vertebrates, and plants of limiting global warming to 1.5 °C rather than 2 °C. Science 2018, 360, 791–795. [Google Scholar] [CrossRef]
- Trisos, C.H.; Merow, C.; Pigot, A.L. The projected timing of abrupt ecological disruption from climate change. Nature 2020, 580, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Panetta, A.M.; Stanton, M.L.; Harte, J. Climate warming drives local extinction: Evidence from observation and experimentation. Sci. Adv. 2018, 4, eaaq1819. [Google Scholar] [CrossRef]
- Kirillova, I.A.; Kirillov, D.V. Impact of weather conditions on seasonal development, population structure and reproductive success on Dactylorhiza traunsteineri (Orchidaceae) in the Komi Republic (Russia). Nat. Conserv. Res. 2020, 5 (Suppl. 1), 77–89. [Google Scholar] [CrossRef]
- Jiang, H.; Chen, J.; Liu, G.; Zhou, P.; Jin, Q.; Wang, Y.; Guo, H.; Qian, P.; Xu, Y. Screening of early flowering lotus (Nelumbo nucifera Gaertn.) cultivars and effects of different cultivars on flowering period. Plants 2023, 12, 1683. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, R.; Bhandari, K.; Nayyar, H. Temperature stress and redox homeostasis in agricultural crops. Front. Environ. Sci. 2015, 3, 11. [Google Scholar] [CrossRef]
- Singh, V.; Nguyen, C.T.; van Oosterom, E.J.; Chapman, S.C.; Jordan, D.R.; Hammer, G.L. Sorghum genotypes differ in high temperature responses for seed set. Field Crops Res. 2015, 171, 32–40. [Google Scholar] [CrossRef]
- Devireddy, A.R.; Tschaplinski, T.J.; Tuskan, G.A.; Muchero, W.; Chen, J.G. Role of reactive oxygen species and hormones in plant responses to temperature changes. Int. J. Mol. Sci. 2021, 22, 8843. [Google Scholar] [CrossRef]
- Monder, M.J.; Bąbelewski, P.; Szperlik, J.; Kościelak, A. The adjustment of China endemic Heptacodium miconioides Rehd. to temperate zone of Poland. BMC Plant Biol. 2023, 23, 184. [Google Scholar] [CrossRef] [PubMed]
- Meineri, E.; Skarpaas, O.; Spindelböck, J.; Bargmann, T.; Vandvik, V. Direct and size-dependent effects of climate on flowering performance in alpine and lowland herbaceous species. J. Veg. Sci. 2014, 25, 275–286. [Google Scholar] [CrossRef]
- Miller-Struttmann, N.E.; Geib, J.C.; Franklin, J.D.; Kevan, P.G.; Holdo, R.M.; Ebert-May, D.; Lynn, A.M.; Kettenbach, J.A.; Hedrick, E.; Galen, C. Functional mismatch in a bumble bee pollination mutualism under climate change. Science 2015, 349, 1541–1544. [Google Scholar] [CrossRef] [PubMed]
- Haeuser, E.; Dawson, W.; Kleunen, M.V. The effects of climate warming and disturbance on the colonization potential of ornamental alien plant species. J. Ecol. 2017, 105, 1698–1708. [Google Scholar] [CrossRef]
- Springate, D.A.; Kover, P.X. Plant responses to elevated temperatures: A field study on phenological sensitivity and fitness responses to simulated climate warming. Glob. Change Biol. 2014, 20, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Hoover, S.E.; Ladley, J.J.; Shchepetkina, A.A.; Tisch, M.; Gieseg, S.P.; Tylianakis, J.M. Warming, CO2, and nitrogen deposition interactively affect a plant-pollinator mutualism. Ecol. Lett. 2012, 15, 227–234. [Google Scholar] [CrossRef] [PubMed]
- de Manincor, N.; Fisogni, A.; Rafferty, N.E. Warming of experimental plant-pollinator communities advances phenologies, alters traits, reduces interactions and depresses reproduction. Ecol. Lett. 2023, 26, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Dela, G.; Or, E.; Ovadia, R.; Nissim-Levi, A.; Weiss, D.; Oren-Shamir, M. Changes in anthocyanin concentration and composition in ‘Jaguar’ rose flowers due to transient high-temperature conditions. Plant Sci. 2003, 164, 333–340. [Google Scholar] [CrossRef]
- Dai, W.K.; Ochola, A.C.; Li, Y.Q. Spatio-temporal variations in pollen limitation and floral traits of an alpine lousewort (Pedicularis rhinanthoides) in relation to pollinator availability. Plants 2022, 12, 78. [Google Scholar] [CrossRef]
- Arroyo, M.T.; Dudley, L.S.; Jespersen, G.; Pacheco, D.A.; Cavieres, L.A. Temperature-driven flower longevity in a high-alpine species of Oxalis influences reproductive assurance. New Phytol. 2013, 200, 1260–1268. [Google Scholar] [CrossRef]
- Seymour, R.S.; Gibernau, M.; Pirintsos, S.A. Thermogenesis of three species of Arum from Crete. Plant Cell Environ. 2009, 32, 1467–1476. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.L. Floral of China, 2nd, ed.; Beijing Science Press: Beijing, China, 2001; Volume 47. [Google Scholar]
- Dan, Y.; Baxter, A.; Zhang, S.; Pantazis, C.J.; EVeilleux, R. Development of efficient plant regeneration and transformation system for impatiens using agrobacterium tumefaciens and multiple bud cultures as explants. BMC Plant Biol. 2010, 10, 165. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.F.; Ding, B.Y. The impatiens of Zhejiang wild flower resources and development. Chin. Wild Plant Resour. 2000, 19, 27–29. [Google Scholar]
- Wang, Y. Collection and Preservation of Impatiens spp. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2008. [Google Scholar]
- Wang, Q. Biological Effects of Experimental Warming on Pollination in Impatiens oxyanthera (Balsaminaceae). Ph.D. Thesis, The University of Chinese Academy of Sciences, Beijing, China, 2013. [Google Scholar]
- Cui, M.; Yang, B.; Ren, G.; Yu, H.; Dai, Z.; Li, J.; Ran, Q.; Stevanato, P.; Wan, J.; Du, D. Effects of warming, phosphorous deposition, and both treatments on the growth and physiology of invasive Solidago canadensis and native Artemisia argyi. Plants 2023, 12, 1370. [Google Scholar] [CrossRef] [PubMed]
- Marlène, A.; François, O. Growth temperature affects inflorescence architecture in Arabidopsis thaliana. Botany 2013, 91, 642–651. [Google Scholar]
- Lysenko, E.A.; Kozuleva, M.A.; Klaus, A.A.; Pshybytko, N.L.; Kusnetsov, V.V. Lower air humidity reduced both the plant growth and activities of photosystems I and II under prolonged heat stress. Plant Physiol. Biochem. 2023, 194, 246–262. [Google Scholar] [CrossRef] [PubMed]
- Nagarathn, T.; Shadakshari, Y.; Jagadish, K.; Sanjay, M. Interactions of auxin and cytokinins in regulating axillary bud formation in sunflower (Helianthus annuus L.). Helia 2010, 33, 85–94. [Google Scholar] [CrossRef]
- Li, Y.B.; Hou, R.X.; Tao, F.L. Wheat morpho-physiological traits and radiation use efficiency under interactive effects of warming and tillage management. Plant Cell Environ. 2020, 44, 2386–2401. [Google Scholar] [CrossRef]
- Chen, B.-M.; Gao, Y.; Liao, H.-X.; Peng, S.-L. Differential responses of invasive and native plants to warming with simulated changes in diurnal temperature ranges. AoB Plants 2017, 9, plx028. [Google Scholar] [CrossRef]
- Aspi, J.; Jäkäläniemi, A.; Tuomi, J.; Siikamäki, P. Multilevel phenotypic selection on morphological characters in a metapopulation of Silene tatarica. Evolution 2003, 57, 509–517. [Google Scholar]
- Zeng, Z.; Huan, H.H.; Liu, G.; Xiao, J.; Huang, Y.Y.; Xu, X.; Dong, T.F. Effects of elevated temperature and CO2 concentration on growth and leaf quality of Morus alba seedlings. Chin. J. Appl. Ecol. 2016, 27, 2445–2451. [Google Scholar]
- Sage, T.L.; Bagha, S.; Lundsgaard-Nielsen, V.; Branch, H.A.; Sultmanis, S.; Sage, R.F. The effect of high temperature stress on male and female reproduction in plants. Field Crop. Res. 2015, 182, 30–42. [Google Scholar] [CrossRef]
- Ishii, H.S.; Sakai, S. Effects of display size and position on individual floral longevity in racemes of Narthecium asiaticum (Liliaceae). Funct. Ecol. 2001, 15, 396–405. [Google Scholar] [CrossRef]
- Itagaki, T.; Sakai, S. Relationship between floral longevity and sex allocation among flowers within inflorescences in Aquilegia buergeriana var. Oxysepala (Ranunculaceae). Am. J. Bot. 2006, 93, 1320–1327. [Google Scholar] [CrossRef] [PubMed]
- Bock, A.; Sparks, T.H.; Estrella, N.; Jee, N.; Casebow, A.; Schunk, C.; Leuchner, M.; Menzel, A. Changes in first flowering dates and flowering duration of 232 plant species on the island of Guernsey. Glob. Change Biol. 2014, 20, 3508–3519. [Google Scholar] [CrossRef] [PubMed]
- Sood, A.; Duchin, S.; Adamov, Z.; Carmeli-Weissberg, M.; Shaya, F.; Spitzer-Rimon, B. Abscisic acid mediates the reduction of petunia flower size at elevated temperatures due to reduced cell division. Planta 2022, 255, 18. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yang, Y.; Wang, Q. Photosynthetic physiological response of Impatiens oxyanthera to Simulated Warming. J. China West Norm. Univ. (Nat. Sci.) 2019, 40, 339–345. [Google Scholar]
- Suraweera, D.D.; Groom, T.; Nicolas, M.E. Nicolas. Exposure to heat stress during flowering period reduces flower yield and pyrethrins in Pyrethrum (Tanacetum cinerariifolium). J. Agron. Crop Sci. 2020, 206, 568–578. [Google Scholar] [CrossRef]
- Lambrecht, S.C. Floral water costs and size variation in the highly selfing Leptosiphon bicolor (Polemoniaceae). Int. J. Plant Sci. 2013, 174, 74–84. [Google Scholar] [CrossRef]
- Gómez, J.M.; Bosch, J.; Perfectti, F.; Fernandez, J.D.; Abdelaziz, M.; Camacho, J.P.M. Association between floral traits and rewards in Erysimum mediohispanicum (Brassicaceae). Ann. Bot. 2008, 101, 1413–1420. [Google Scholar] [CrossRef] [PubMed]
- Ellis, A.G.; Johnson, S.D.; Conner, J.K. Gender differences in the effects of floral spur length manipulation on fitness in a hermaphrodite orchid. Int. J. Plant Sci. 2010, 171, 1010–1019. [Google Scholar] [CrossRef]
- Sletvold, N.; Ågren, J. Nonadditive effects of floral display and spur length on reproductive success in a deceptive orchid. Ecology 2011, 92, 2167–2174. [Google Scholar] [CrossRef] [PubMed]
- Boberg, E.; Ågren, J. Despite their apparent integration, spur length but not perianth size affects reproductive success in the moth-pollinated orchid Platanthera bifolia. Funct. Ecol. 2009, 23, 1022–1028. [Google Scholar] [CrossRef]
- Jia, Y.; Zhao, J.-L.; Pan, Y.-Z.; Xu, Y.; Sun, L.-X.; Liu, Q.-L. Collection and evaluation of Primula species of western Sichuan in China. Genet. Resour. Crop Evol. 2014, 61, 1245–1262. [Google Scholar] [CrossRef]
- Shen, G.; Wang, W. Circlize package in R and Analytic Hierarchy Process (AHP): Contribution values of ABCDE and AGL6 genes in the context of floral organ development. PLoS ONE 2022, 17, e0261232. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.S.; Chen, L.J.; Yang, X.J.; Duan, L.D. A comprehensive evaluation of the wild ground cover plants resources in Yunshan, Hunan. Acta Prataculturae Sin. 2015, 24, 30–40. [Google Scholar]
- Cicevan, R.; Sestras, A.F.; Plazas, M.; Boscaiu, M.; Vilanova, S.; Gramazio, P.; Vicente, O.; Prohens, J.; Sestras, R.E. Biological traits and genetic relationships amongst cultivars of three species of tagetes (Asteraceae). Plants 2022, 11, 760. [Google Scholar] [CrossRef]
- Xing, G.; Qu, L.; Zhang, Y.; Xue, L.; Su, J.; Lei, J. Collection and evaluation of wild tulip (Tulipa spp.) resources in China. Genet. Resour. Crop Evol. 2017, 64, 641–652. [Google Scholar] [CrossRef]
- Yang, Z.; Meng, T.F.; Bi, X.Y.; Lei, J.J. Investigation and evaluation of wild Iris resources in Liaoning Province, China. Genet. Resour. Crop Evol. 2017, 64, 967–978. [Google Scholar]
- Ai, Y.; Chen, L.; Lan, S.R.; Xu, J.Q.; Wang, C.S.; Peng, D.H. Comprehensive evaluation of the ornamental value of Cymbidium ensifolium cultivars using analytical hierarchy process method. J. Fujian Agric. For. Univ. (Nat. Sci. Ed.) 2019, 48, 736–741. [Google Scholar]
- Chandel, A.; Thakur, M.; Rakwal, A.; Chauhan, S.; Bhargava, B. Exogenous applications of gibberellic acid modulate the growth, flowering and longevity of calla lily. Heliyon 2023, 9, e16319. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Wu, H.; Qin, J.; Lin, C.; Tang, W.; Chen, Y. Recent climate changes over the Tibetan Plateau and their impacts on energy and water cycle: A review. Glob. Planet. Change 2014, 112, 79–91. [Google Scholar] [CrossRef]
- Liu, L.; Wu, W.; Zheng, Y.L.; Huang, C.Y.; Liu, R.J. Variations on the chemical components of the volatile oil of Houttuynia cordata Thunb. populations from different valleys and altitudes of Mt. Emei. Acta Ecol. Sin. 2007, 27, 2239–2250. [Google Scholar]
- Gu, H.Y.; Li, C.H. Biodiversity and flora of the mixed evergreen and deciduous broadleaved forest in Emei. Bull. Bot. Res. 2006, 26, 618–624. [Google Scholar]
- Li, Z.Y.; Shi, L. Plants of Mount Emei; Beijing Science and Technology Press: Beijing, China, 2007. [Google Scholar]
- Zhao, Q.Y.; Zhang, X.; Cao, M.H.; Tao, Y.D.; Zhou, M.; Huang, H.Q.; Qu, S.P.; Huang, M.J. Investigation and evaluation on plant resources of Impatiens in southwest Sichuan. Seed 2023, 42, 64–71+82. [Google Scholar]
- Field, C.B.; Barros, V.R.; Dokken, D.J. (Eds.) IPCC(2014) Summary for Policymakers. Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral As-pects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; World Meteorological Organization: Geneva, Switzerland, 2014; pp. 1–190. [Google Scholar]
- Chen, D.F.; Zhang, Y.; Fang, Z. Study on the content of anthocyanin and related biochemical substances during the petal development in Impatiens hawkeri. J. Agric. Univ. Hebei 2008, 31, 28–32. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S.; Christensen, R.H.B.; Singmann, H.; Dai, B.; Scheipl, F.; Grothendieck, G.; Green, P.; et al. lme4: Linear Mixed-Effects Models Using ‘Eigen’ and s4. R Package Version 1.1-21. 2019. Available online: https://cran.r-project.org/package=lme4/ (accessed on 28 June 2021).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression; Sage Publications: Thousand Oaks, CA, USA, 2011. [Google Scholar]
- Lenth, R.; Boker, B.; Buerkner, P.; Giné-Vázquez, I.; Herve, M.; Jung, M.; Love, J.; Miguez, J.; Riebl, H.; Singmann, H. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Ver. 1. 5. 1. 2020. Available online: https://CRAN.Rproject.org/package=emmeans (accessed on 28 June 2021).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | 2017 | 2018 | W | Y | W × Y | ||
|---|---|---|---|---|---|---|---|
| Control | Warming | Control | Warming | ||||
| Male phase (d) | 2.417 ± 0.072 ab | 2.185 ± 0.068 b | 2.898 ± 0.093 a | 2.583 ± 0.085 ab | NS | * | NS |
| Female phase (d) | 1.065 ± 0.051 a | 1.028 ± 0.051 a | 1.333 ± 0.068 a | 1.213 ± 0.052 a | NS | NS | NS |
| Flower longevity (d) | 3.481 ± 0.083 b | 3.213 ± 0.076 b | 4.231 ± 0.085 a | 3.796 ± 0.070 ab | NS | ** | NS |
| Flowering duration per plant (d) | 65.278 ± 1.474 a | 58.944 ± 1.330 b | 65.917 ± 1.899 a | 53.361 ± 1.840 c | ** | NS | ** |
| Flower number per plant (No.) | 80.778 ± 5.155 a | 73.306 ± 4.251 b | 73.472 ± 4.357 b | 65.500 ± 5.361 c | *** | *** | NS |
| Trait | 2017 | 2018 | W | Y | W × Y | ||
|---|---|---|---|---|---|---|---|
| Control | Warming | Control | Warming | ||||
| Vexillum length (mm) | 12.776 ± 0.245 a | 11.840 ± 0.213 b | 12.546 ± 0.101 a | 12.181 ± 0.152 ab | *** | NS | NS |
| Wing petal length (mm) | 23.058 ± 0.240 ab | 22.289 ± 0.258 b | 23.650 ± 0.263 a | 22.807 ± 0.217 ab | * | NS | NS |
| Corolla diameter (mm) | 21.916 ± 0.407 b | 21.626 ± 0.360 b | 23.434 ± 0.361 a | 22.465 ± 0.384 ab | NS | ** | NS |
| Corolla tube length (mm) | 20.202 ± 0.310 a | 19.275 ± 0.220 b | 20.444 ± 0.215 a | 18.637 ± 0.252 b | *** | NS | NS |
| Stripe number on the labellum (No.) | 11.167 ± 0.232 a | 10.713 ± 0.193 a | 10.667 ± 0.183 a | 10.222 ± 0.186 a | NS | NS | NS |
| Nectar spur length (mm) | 30.079 ± 0.399 a | 27.447 ± 0.536 b | 28.458 ± 0.410 b | 27.820 ± 0.314 b | *** | ** | ** |
| Nectar spur curvature (°) | 303.333 ± 12.626 b | 300.000 ± 10.992 b | 334.352 ± 10.114 a | 326.389 ± 9.408 a | NS | * | NS |
| Pedicel length (mm) | 46.285 ± 1.725 a | 38.664 ± 1.292 b | 40.055 ± 1.107 b | 38.960 ± 1.379 b | *** | ** | * |
| Relative anthocyanin content of vexillum (A. g−1 FW) | 5.624 ± 0.136 a | 5.867 ± 0.159 a | 5.563 ± 0.120 a | 5.851 ± 0.114 a | NS | NS | NS |
| Relative anthocyanin content of corolla tube (A. g−1 FW) | 3.659 ± 0.078 a | 3.722 ± 0.095 a | 3.560 ± 0.056 a | 3.723 ± 0.112 a | NS | NS | NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, J.; Yang, Y.; Wang, Q. Two Growing-Season Warming Partly Promoted Growth but Decreased Reproduction and Ornamental Value of Impatiens oxyanthera. Plants 2024, 13, 511. https://doi.org/10.3390/plants13040511
Tao J, Yang Y, Wang Q. Two Growing-Season Warming Partly Promoted Growth but Decreased Reproduction and Ornamental Value of Impatiens oxyanthera. Plants. 2024; 13(4):511. https://doi.org/10.3390/plants13040511
Chicago/Turabian StyleTao, Jiayu, Youqin Yang, and Qiong Wang. 2024. "Two Growing-Season Warming Partly Promoted Growth but Decreased Reproduction and Ornamental Value of Impatiens oxyanthera" Plants 13, no. 4: 511. https://doi.org/10.3390/plants13040511
APA StyleTao, J., Yang, Y., & Wang, Q. (2024). Two Growing-Season Warming Partly Promoted Growth but Decreased Reproduction and Ornamental Value of Impatiens oxyanthera. Plants, 13(4), 511. https://doi.org/10.3390/plants13040511