
Stable Isotope Analyses Reveal Impact of Fe and Zn on Cd Uptake and Translocation by Theobroma cacao


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Growth and Sample Preparation
2.2. Cadmium Concentration and Stable Isotope Analysis
2.3. Statistical Analysis
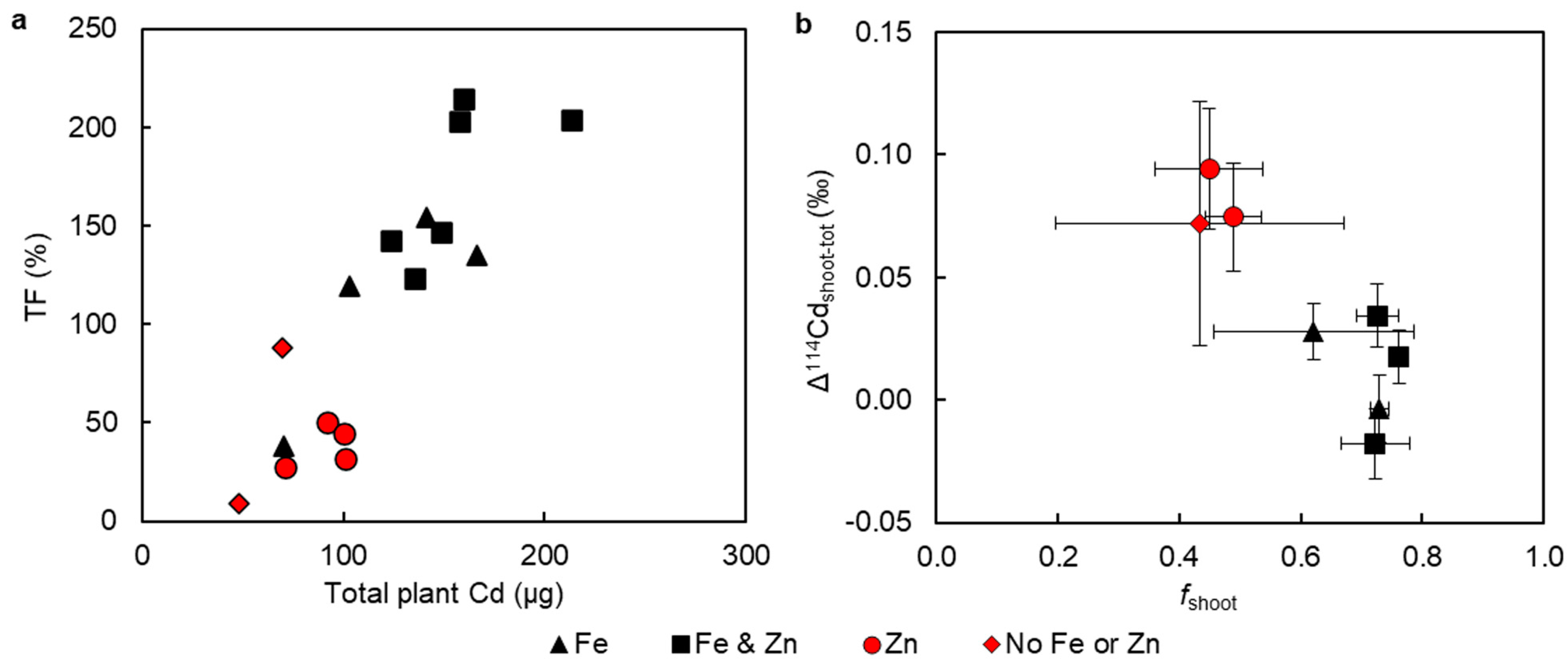
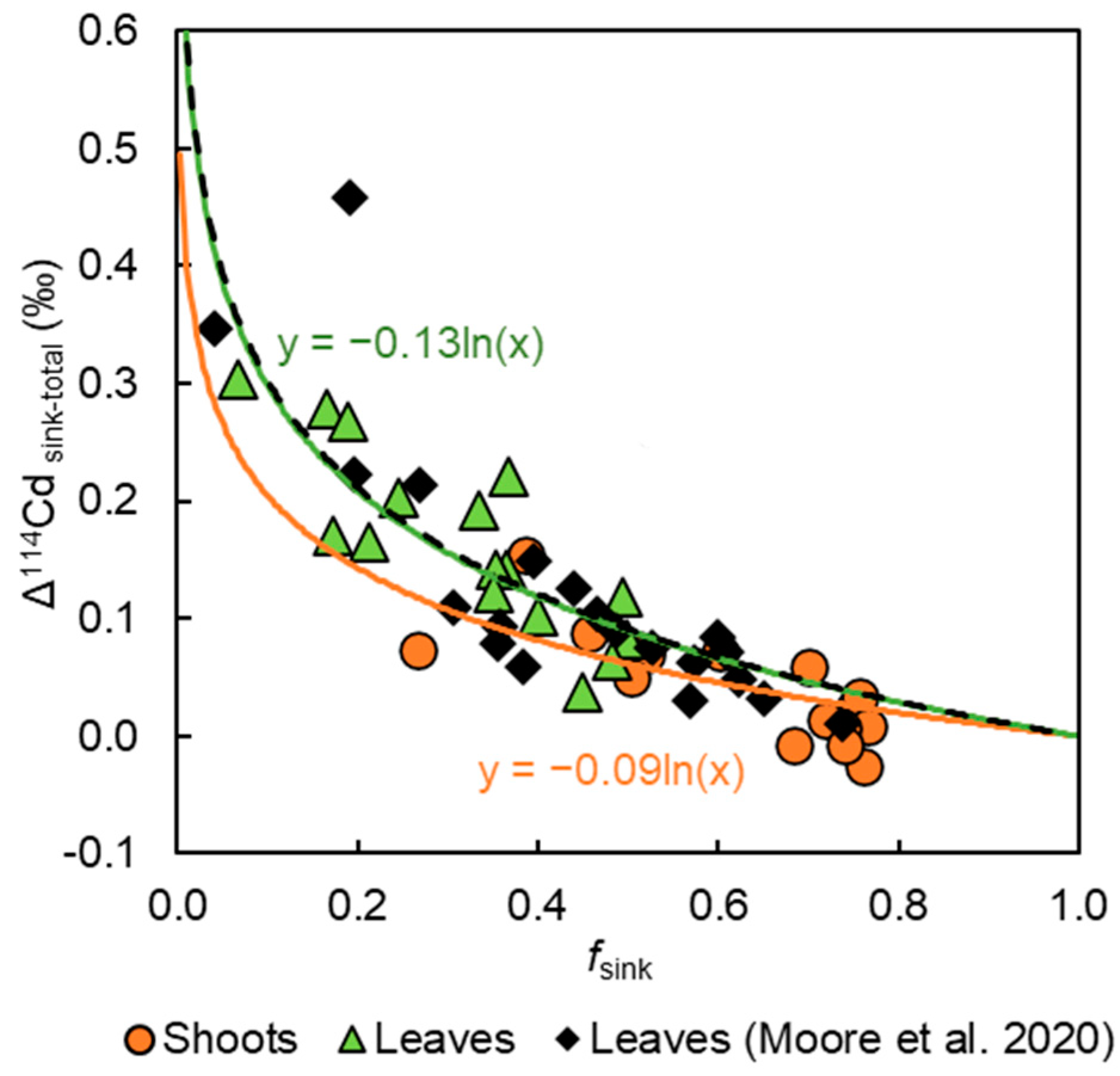
3. Results
3.1. Quality Control
3.2. Cd Speciation in the Hydroponic Solutions
3.3. Cadmium Concentrations and Isotope Compositions
4. Discussion
4.1. Effect of Fe and Zn on Cd Uptake
4.2. Effect of Fe and Zn Treatments on Cd Translocation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhatla, S.C.; Lal, M.A. Plant Physiology, Development and Metabolism; Springer: Singapore, 2018; ISBN 9789811320224. [Google Scholar]
- de Bang, T.C.; Husted, S.; Laursen, K.H.; Persson, D.P.; Schjoerring, J.K. The Molecular-physiological Functions of Mineral Macronutrients and Their Consequences for Deficiency Symptoms in Plants. New Phytol. 2020, 229, 2446–2469. [Google Scholar] [CrossRef]
- Küpper, H.; Andresen, E. Mechanisms of Metal Toxicity in Plants. Metallomics 2016, 8, 269–285. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Liu, H.; Nie, Z.; Rengel, Z.; Gao, W.; Li, C.; Zhao, P. Toxicity of Cadmium and Its Competition with Mineral Nutrients for Uptake by Plants: A Review. Pedosphere 2020, 30, 168–180. [Google Scholar] [CrossRef]
- de Oliveira, V.H.; Ullah, I.; Dunwell, J.M.; Tibbett, M. Mycorrhizal Symbiosis Induces Divergent Patterns of Transport and Partitioning of Cd and Zn in Populus trichocarpa. Environ. Exp. Bot. 2020, 171, 103925. [Google Scholar] [CrossRef]
- Palmer, C.M.; Guerinot, M.L. Facing the Challenges of Cu, Fe and Zn Homeostasis in Plants. Nat. Chem. Biol. 2009, 5, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, S.A.; Krämer, U. The Zinc Homeostasis Network of Land Plants. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 1553–1567. [Google Scholar] [CrossRef] [PubMed]
- Perfus-Barbeoch, L.; Leonhardt, N.; Vavasseur, A.; Forestier, C. Heavy Metal Toxicity: Cadmium Permeates through Calcium Channels and Disturbs the Plant Water Status. Plant J. 2002, 32, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Argüello, D.; Chavez, E.; Lauryssen, F.; Vanderschueren, R.; Smolders, E.; Montalvo, D. Soil Properties and Agronomic Factors Affecting Cadmium Concentrations in Cacao Beans: A Nationwide Survey in Ecuador. Sci. Total Environ. 2019, 649, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Fechner, C.; Greiner, M.; Heseker, H.; Lindtner, O. Dietary Exposure Assessment of Aluminium and Cadmium from Cocoa in Relation to Cocoa Origin. PLoS ONE 2019, 14, e0217990. [Google Scholar] [CrossRef]
- Maddela, N.R.; Kakarla, D.; García, L.C.; Chakraborty, S.; Venkateswarlu, K.; Megharaj, M. Cocoa-Laden Cadmium Threatens Human Health and Cacao Economy: A Critical View. Sci. Total Environ. 2020, 720, 137645. [Google Scholar] [CrossRef]
- Rai, P.K.; Lee, S.S.; Zhang, M.; Tsang, Y.F.; Kim, K.-H. Heavy Metals in Food Crops: Health Risks, Fate, Mechanisms, and Management. Environ. Int. 2019, 125, 365–385. [Google Scholar] [CrossRef] [PubMed]
- Vanderschueren, R.; De Mesmaeker, V.; Mounicou, S.; Isaure, M.P.; Doelsch, E.; Montalvo, D.; Delcour, J.A.; Chavez, E.; Smolders, E. The Impact of Fermentation on the Distribution of Cadmium in Cacao Beans. Food Res. Int. 2020, 127, 108743. [Google Scholar] [CrossRef]
- Vanderschueren, R.; Montalvo, D.; De Ketelaere, B.; Delcour, J.A.; Smolders, E. The Elemental Composition of Chocolates Is Related to Cacao Content and Origin: A Multi-Element Fingerprinting Analysis of Single Origin Chocolates. J. Food Compos. Anal. 2019, 83, 103277. [Google Scholar] [CrossRef]
- Zug, K.L.M.; Huamaní Yupanqui, H.A.; Meyberg, F.; Cierjacks, J.S.; Cierjacks, A. Cadmium Accumulation in Peruvian Cacao (Theobroma cacao L.) and Opportunities for Mitigation. Water, Air, Soil Pollut. 2019, 230, 72. [Google Scholar] [CrossRef]
- Engbersen, N.; Gramlich, A.; Lopez, M.; Schwarz, G.; Hattendorf, B.; Gutierrez, O.; Schulin, R. Cadmium Accumulation and Allocation in Different Cacao Cultivars. Sci. Total Environ. 2019, 678, 660–670. [Google Scholar] [CrossRef] [PubMed]
- Vanderschueren, R.; Argüello, D.; Blommaert, H.; Montalvo, D.; Barraza, F.; Maurice, L.; Schreck, E.; Schulin, R.; Lewis, C.; Vazquez, J.L.; et al. Mitigating the Level of Cadmium in Cacao Products: Reviewing the Transfer of Cadmium from Soil to Chocolate Bar. Sci. Total Environ. 2021, 781, 146779. [Google Scholar] [CrossRef]
- Lewis, C.; Lennon, A.M.; Eudoxie, G.; Umaharan, P. Genetic Variation in Bioaccumulation and Partitioning of Cadmium in Theobroma cacao L. Sci. Total Environ. 2018, 640–641, 696–703. [Google Scholar] [CrossRef]
- The European Commission. Commission Regulation (EU) No 488/ 2014 of 12 May 2014 Amending Regulation (EC) No 1881/2006 as Regards Maximum Levels of Cadmium in Foodstuffs. Off. J. Eur. Union 2014, 138, 75–79. [Google Scholar]
- dos Santos, M.L.S.; de Almeida, A.A.F.; da Silva, N.M.; Oliveira, B.R.M.; Silva, J.V.S.; Junior, J.O.S.; Ahnert, D.; Baligar, V.C. Mitigation of Cadmium Toxicity by Zinc in Juvenile Cacao: Physiological, Biochemical, Molecular and Micromorphological Responses. Environ. Exp. Bot. 2020, 179, 104201. [Google Scholar] [CrossRef]
- Du, J.; Zeng, J.; Ming, X.; He, Q.; Tao, Q.; Jiang, M.; Gao, S.; Li, X.; Lei, T.; Pan, Y.; et al. The Presence of Zinc Reduced Cadmium Uptake and Translocation in Cosmos bipinnatus Seedlings under Cadmium/Zinc Combined Stress. Plant Physiol. Biochem. 2020, 151, 223–232. [Google Scholar] [CrossRef]
- Lin, Y.F.; Aarts, M.G.M. The Molecular Mechanism of Zinc and Cadmium Stress Response in Plants. Cell. Mol. Life Sci. 2012, 69, 3187–3206. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Rehman, M.Z.U.; Maqbool, A. A Critical Review on the Effects of Zinc at Toxic Levels of Cadmium in Plants. Environ. Sci. Pollut. Res. 2019, 26, 6279–6289. [Google Scholar] [CrossRef]
- Huang, X.; Duan, S.; Wu, Q.; Yu, M.; Shabala, S. Reducing Cadmium Accumulation in Plants: Structure–Function Relations and Tissue-Specific Operation of Transporters in the Spotlight. Plants 2020, 9, 223. [Google Scholar] [CrossRef]
- Huang, G.; Ding, C.; Zhou, Z.; Zhang, T.; Wang, X. A Tillering Application of Zinc Fertilizer Based on Basal Stabilization Reduces Cd Accumulation in Rice (Oryza sativa L.). Ecotoxicol. Environ. Saf. 2019, 167, 338–344. [Google Scholar] [CrossRef]
- Lv, G.; Wang, H.; Xu, C.; Shuai, H.; Luo, Z.; Zhang, Q.; Zhu, H.; Wang, S.; Zhu, Q.; Zhang, Y.; et al. Effectiveness of Simultaneous Foliar Application of Zn and Mn or P to Reduce Cd Concentration in Rice Grains: A Field Study. Environ. Sci. Pollut. Res. 2019, 26, 9305–9313. [Google Scholar] [CrossRef]
- Wang, H.; Xu, C.; Luo, Z.C.; Zhu, H.H.; Wang, S.; Zhu, Q.H.; Huang, D.Y.; Zhang, Y.Z.; Xiong, J.; He, Y.B. Foliar Application of Zn Can Reduce Cd Concentrations in Rice (Oryza sativa L.) under Field Conditions. Environ. Sci. Pollut. Res. 2018, 25, 29287–29294. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, Y.; Chen, W.; Wang, M.; Wang, T.; Dai, Y. Dynamic Interactions between Soil Cadmium and Zinc Affect Cadmium Phytoavailability to Rice and Wheat: Regional Investigation and Risk Modeling. Environ. Pollut. 2020, 267, 115613. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Long, J.; Li, J.; Zhang, M.; Xiao, G.; Ye, X.; Chang, W.; Zeng, H. Impact of ZnO Nanoparticles on Cd Toxicity and Bioaccumulation in Rice (Oryza sativa L.). Environ. Sci. Pollut. Res. 2019, 26, 23119–23128. [Google Scholar] [CrossRef]
- Wang, P.; Chen, H.; Kopittke, P.M.; Zhao, F.-J. Cadmium Contamination in Agricultural Soils of China and the Impact on Food Safety. Environ. Pollut. 2019, 249, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Saifullah; Javed, H.; Naeem, A.; Rengel, Z.; Dahlawi, S. Timing of Foliar Zn Application Plays a Vital Role in Minimizing Cd Accumulation in Wheat. Environ. Sci. Pollut. Res. 2016, 23, 16432–16439. [Google Scholar] [CrossRef]
- Sharifan, H.; Ma, X.; Moore, J.M.; Habib, M.R.; Evans, C. Zinc Oxide Nanoparticles Alleviated the Bioavailability of Cadmium and Lead and Changed the Uptake of Iron in Hydroponically Grown Lettuce (Lactuca sativa L. Var. Longifolia). ACS Sustain. Chem. Eng. 2019, 7, 16401–16409. [Google Scholar] [CrossRef]
- Zare, A.A.; Khoshgoftarmanesh, A.H.; Malakouti, M.J.; Bahrami, H.A.; Chaney, R.L. Root Uptake and Shoot Accumulation of Cadmium by Lettuce at Various Cd:Zn Ratios in Nutrient Solution. Ecotoxicol. Environ. Saf. 2018, 148, 441–446. [Google Scholar] [CrossRef]
- Wu, S.; Shi, K.; Hu, C.; Guo, J.; Tan, Q.; Sun, X. Non-Invasive Microelectrode Cadmium Flux Measurements Reveal the Decrease of Cadmium Uptake by Zinc Supply in Pakchoi Root (Brassica chinensis L.). Ecotoxicol. Environ. Saf. 2019, 168, 363–368. [Google Scholar] [CrossRef]
- Kutrowska, A.; Małecka, A.; Piechalak, A.; Masiakowski, W.; Hanć, A.; Barałkiewicz, D.; Andrzejewska, B.; Zbierska, J.; Tomaszewska, B. Effects of Binary Metal Combinations on Zinc, Copper, Cadmium and Lead Uptake and Distribution in Brassica Juncea. J. Trace Elem. Med. Biol. 2017, 44, 32–39. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, V.H.; Tibbett, M. Tolerance, Toxicity and Transport of Cd and Zn in Populus trichocarpa. Environ. Exp. Bot. 2018, 155, 281–292. [Google Scholar] [CrossRef]
- Arévalo-Gardini, E.; Arévalo-Hernández, C.O.; Baligar, V.C.; He, Z.L. Heavy Metal Accumulation in Leaves and Beans of Cacao (Theobroma cacao L.) in Major Cacao Growing Regions in Peru. Sci. Total Environ. 2017, 605–606, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Neto, R.D.O.C.; Júnior, J.O.D.S.; Sodré, G.A.; Baligar, V.C. Growth and Nutrition of Cacao Seedlings Influenced by Zinc Aplication in Soil. Rev. Bras. Frutic. 2015, 37, 1053–1064. [Google Scholar] [CrossRef]
- Júnior, J.O.d.S.; da Silveira, F.G.F.; dos Santos, R.O.; Neves, J.C.L. Zinc Fertilizers and Additives for Foliar Fertilization of Cocoa Seedlings. J. Agric. Sci. 2018, 11, 471. [Google Scholar] [CrossRef]
- Huang, D.; Dai, W. Two Iron-Regulated Transporter (IRT) Genes Showed Differential Expression in Poplar Trees under Iron or Zinc Deficiency. J. Plant Physiol. 2015, 186–187, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, H.; Ogawa, I.; Ishimaru, Y.; Mori, S.; Nishizawa, N.K. Iron Deficiency Enhances Cadmium Uptake and Translocation Mediated by the Fe2+ Transporters OsIRT1 and OsIRT2 in Rice. Soil Sci. Plant Nutr. 2006, 52, 464–469. [Google Scholar] [CrossRef]
- Bashir, A.; Rizwan, M.; Ali, S.; Zia ur Rehman, M.; Ishaque, W.; Atif Riaz, M.; Maqbool, A. Effect of Foliar-Applied Iron Complexed with Lysine on Growth and Cadmium (Cd) Uptake in Rice under Cd Stress. Environ. Sci. Pollut. Res. 2018, 25, 20691–20699. [Google Scholar] [CrossRef]
- Chen, Z.; Tang, Y.T.; Yao, A.J.; Cao, J.; Wu, Z.H.; Peng, Z.R.; Wang, S.Z.; Xiao, S.; Baker, A.J.M.; Qiu, R.L. Mitigation of Cd Accumulation in Paddy Rice (Oryza sativa L.) by Fe Fertilization. Environ. Pollut. 2017, 231, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Ali, S.; Rizwan, M.; Rehman, M.Z.U.; Qayyum, M.F.; Wang, H.; Rinklebe, J. Responses of Wheat (Triticum aestivum) Plants Grown in a Cd Contaminated Soil to the Application of Iron Oxide Nanoparticles. Ecotoxicol. Environ. Saf. 2019, 173, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Chen, X.; Chai, S.; Sheng, H.; Sha, L.; Fan, X.; Zeng, J.; Kang, H.; Zhang, H.; Xiao, X.; et al. TpIRT1 is a Transition Metal Transporter in Polish Wheat (Triticum polonicum L.) with a Broad Substrate Specificity. 2021. [CrossRef]
- Cohen, C.K.; Garvin, D.F.; Kochian, L.V. Kinetic Properties of a Micronutrient Transporter from Pisum sativum Indicate a Primary Function in Fe Uptake from the Soil. Planta 2004, 218, 784–792. [Google Scholar] [CrossRef]
- Wiggenhauser, M.; Moore, R.E.T.; Wang, P.; Bienert, G.P.; Laursen, K.H.; Blotevogel, S. Stable Isotope Fractionation of Metals and Metalloids in Plants: A Review. Front. Plant Sci. 2022, 13, 840941. [Google Scholar] [CrossRef] [PubMed]
- Arnold, T.; Markovic, T.; Kirk, G.J.D.; Schönbächler, M.; Rehkämper, M.; Zhao, F.J.; Weiss, D.J. Iron and Zinc Isotope Fractionation during Uptake and Translocation in Rice (Oryza sativa) Grown in Oxic and Anoxic Soils. Comptes Rendus Geosci. 2015, 347, 397–404. [Google Scholar] [CrossRef]
- Arnold, T.; Kirk, G.J.D.; Wissuwa, M.; Frei, M.; Zhao, F.J.; Mason, T.F.D.; Weiss, D.J. Evidence for the Mechanisms of Zinc Uptake by Rice Using Isotope Fractionation. Plant Cell Environ. 2010, 33, 370–381. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Gao, T.; Liu, Y.; Liu, J.; Li, F.; Chen, Z.; Li, Y.; Lv, Y.; Song, Z.; Reinfelder, J.R.; et al. Isotopic Fingerprints Indicate Distinct Strategies of Fe Uptake in Rice. Chem. Geol. 2019, 524, 323–328. [Google Scholar] [CrossRef]
- Moore, R.E.T.; Ullah, I.; de Oliveira, V.H.; Hammond, S.J.; Strekopytov, S.; Tibbett, M.; Dunwell, J.M.; Rehkämper, M. Cadmium Isotope Fractionation Reveals Genetic Variation in Cd Uptake and Translocation by Theobroma cacao and Role of Natural Resistance-Associated Macrophage Protein 5 and Heavy Metal ATPase-Family Transporters. Hortic. Res. 2020, 7, 71. [Google Scholar] [CrossRef]
- Wiggenhauser, M.; Bigalke, M.; Imseng, M.; Müller, M.; Keller, A.; Murphy, K.; Kreissig, K.; Rehkämper, M.; Wilcke, W.; Frossard, E. Cadmium Isotope Fractionation in Soil–Wheat Systems. Environ. Sci. Technol. 2016, 50, 9223–9231. [Google Scholar] [CrossRef]
- Wiggenhauser, M.; Aucour, A.-M.; Bureau, S.; Campillo, S.; Telouk, P.; Romani, M.; Ma, J.F.; Landrot, G.; Sarret, G. Cadmium Transfer in Contaminated Soil-Rice Systems: Insights from Solid-State Speciation Analysis and Stable Isotope Fractionation. Environ. Pollut. 2021, 269, 115934. [Google Scholar] [CrossRef]
- Zhang, S.N.; Gu, Y.; Zhu, Z.L.; Hu, S.H.; Kopittke, P.M.; Zhao, F.J.; Wang, P. Stable Isotope Fractionation of Cadmium in the Soil-Rice-Human Continuum. Sci. Total Environ. 2021, 761, 143262. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Li, X.; Li, F.; Liu, T.; Huang, F.; Yin, H.; Chen, G. Water Management Alters Cadmium Isotope Fractionation between Shoots and Nodes/Leaves in a Soil-Rice System. Environ. Sci. Technol. 2021, 55, 12902–12913. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Li, X.; Li, F.; Huang, Y.; Liu, T.; Yin, H.; Qiao, J.; Chen, G.; Huang, F. Cadmium Uptake and Transport Processes in Rice Revealed by Stable Isotope Fractionation and Cd-Related Gene Expression. Sci. Total Environ. 2021, 806, 150633. [Google Scholar] [CrossRef]
- Imseng, M.; Wiggenhauser, M.; Keller, A.; Müller, M.; Rehkämper, M.; Murphy, K.; Kreissig, K.; Frossard, E.; Wilcke, W.; Bigalke, M. Towards an Understanding of the Cd Isotope Fractionation during Transfer from the Soil to the Cereal Grain. Environ. Pollut. 2019, 244, 834–844. [Google Scholar] [CrossRef]
- Barraza, F.; Moore, R.E.T.; Rehkämper, M.; Schreck, E.; Lefeuvre, G.; Kreissig, K.; Coles, B.J.; Maurice, L. Cadmium Isotope Fractionation in the Soil-Cacao Systems of Ecuador: A Pilot Field Study. RSC Adv. 2019, 9, 34011–34022. [Google Scholar] [CrossRef]
- Blommaert, H.; Aucour, A.; Wiggenhauser, M.; Moens, C.; Telouk, P.; Landrot, G.; Testemale, D.; Pin, S.; Lewis, C.; Umaharan, P.; et al. From Soil to Cacao Bean: Unravelling the Pathways of Cadmium Translocation in a High Cd Accumulating Cultivar of Theobroma cacao L. Front. Plant Sci. 2022, 13, 1055912. [Google Scholar] [CrossRef]
- Wei, R.; Guo, Q.; Wen, H.; Liu, C.; Yang, J.; Peters, M.; Hu, J.; Zhu, G.; Zhang, H.; Tian, L.; et al. Fractionation of Stable Cadmium Isotopes in the Cadmium Tolerant Ricinus communis and Hyperaccumulator Solanum nigrum. Sci. Rep. 2016, 6, 24309. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Guo, Q.; Yu, G.; Kong, J.; Li, S.; Song, Z.; Hu, J.; Tian, L.; Han, X.; Okoli, C.P. Stable Isotope Fractionation during Uptake and Translocation of Cadmium by Tolerant Ricinus communis and Hyperaccumulator Solanum nigrum as Influenced by EDTA. Environ. Pollut. 2018, 236, 634–644. [Google Scholar] [CrossRef]
- Zhou, J.; Li, Z.; Liu, M.; Yu, H.; Wu, L.; Huang, F.; Luo, Y.; Christie, P. Cadmium Isotopic Fractionation in the Soil − Plant System during Repeated Phytoextraction with a Cadmium Hyperaccumulating Plant Species. Environ. Sci. Technol. 2020, 54, 13598–13609. [Google Scholar] [CrossRef]
- Barati, E.; Moore, R.E.T.; Ullah, I.; Kreissig, K.; Coles, B.J.; Dunwell, J.M.; Rehkämper, M. An Investigation of Zinc Isotope Fractionation in Cacao (Theobroma cacao L.) and Comparison of Zinc and Cadmium Isotope Compositions in Hydroponic Plant Systems under High Cadmium Stress. Sci. Rep. 2023, 13, 4682. [Google Scholar] [CrossRef]
- Wiggenhauser, M.; Bigalke, M.; Imseng, M.; Keller, A.; Archer, C.; Wilcke, W.; Frossard, E. Zinc Isotope Fractionation during Grain Filling of Wheat and a Comparison of Zinc and Cadmium Isotope Ratios in Identical Soil–Plant Systems. New Phytol. 2018, 219, 195–205. [Google Scholar] [CrossRef]
- Zhou, J.; Moore, R.E.T.; Rehkämper, M.; Kreissig, K.; Coles, B.; Sun, Y.; Li, Z.; Luo, Y.; Christie, P.; Wu, L. Zinc Supply Affects Cadmium Uptake and Translocation in the Hyperaccumulator Sedum plumbizincicola as Evidenced by Isotope Fractionation. Environ. Sci. Technol. 2023, 57, 5891–5902. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Moynier, F.; Blichert-Toft, J.; Albarède, F. Density Functional Theory Estimation of Isotope Fractionation of Fe, Ni, Cu, and Zn among Species Relevant to Geochemical and Biological Environments. Geochim. Cosmochim. Acta 2014, 140, 553–576. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, Y.; Wiggenhauser, M.; Yang, J.; Sarret, G.; Cheng, Q.; Liu, J.; Shi, Y. Theoretical Isotope Fractionation of Cadmium during Complexation with Organic Ligands. Chem. Geol. 2021, 571, 120178. [Google Scholar] [CrossRef]
- Murphy, K.; Rehkämper, M.; Kreissig, K.; Coles, B.; van de Flierdt, T. Improvements in Cd Stable Isotope Analysis Achieved through Use of Liquid-Liquid Extraction to Remove Organic Residues from Cd Separates Obtained by Extraction Chromatography. J. Anal. At. Spectrom. 2016, 31, 319–327. [Google Scholar] [CrossRef]
- Aucour, A.M.; Pichat, S.; MacNair, M.R.; Oger, P. Fractionation of Stable Zinc Isotopes in the Zinc Hyperaccumulator Arabidopsis halleri and Nonaccumulator Arabidopsis petraea. Environ. Sci. Technol. 2011, 45, 9212–9217. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Rehkämper, M.; Schönbächler, M.; Statham, P.J.; Coles, B.J. A New Methodology for Precise Cadmium Isotope Analyses of Seawater. Anal. Bioanal. Chem. 2012, 402, 883–893. [Google Scholar] [CrossRef]
- Abouchami, W.; Galer, S.J.G.; Horner, T.J.; Rehkämper, M.; Wombacher, F.; Xue, Z.; Lambelet, M.; Gault-Ringold, M.; Stirling, C.H.; Schönbächler, M.; et al. A Common Reference Material for Cadmium Isotope Studies-NIST SRM 3108. Geostand. Geoanalytical Res. 2013, 37, 5–17. [Google Scholar] [CrossRef]
- Lv, W.; Yin, H.; Liu, M.; Huang, F.; Yu, H. Effect of the Dry-Ashing Method on Cadmium Isotope Measurements in Soil and Plant Samples. Geostand. Geoanalytical Res. 2020, 45, 245–256. [Google Scholar] [CrossRef]
- Borovička, J.; Ackerman, L.; Rejšek, J. Cadmium Isotopic Composition of Biogenic Certified Reference Materials Determined by Thermal Ionization Mass Spectrometry with Double Spike Correction. Talanta 2021, 221, 121389. [Google Scholar] [CrossRef]
- Hossain, K.G.; Islam, N.; Ghavami, F.; Durant, C.; Durant, C.; Johnson, M. Effect of Increased Amounts of Fe, Zn, and Cd on Uptake, Translocation, and Accumulation of Human Health Related Micronutrients in Wheat. Asian J. Agric. food Sci. 2017, 5, 19. [Google Scholar] [PubMed]
- Liu, P.; Xiao, W.; Wang, K.; Yang, Z.; Wang, L. Bioaccessibility of Cd and Its Correlation with Divalent Mineral Nutrients in Locally Grown Rice from Two Provinces in China. Biol. Trace Elem. Res. 2021, 200, 1408–1417. [Google Scholar] [CrossRef] [PubMed]
- Boza, E.J.; Motamayor, J.C.; Amores, F.M.; Cedeño-Amador, S.; Tondo, C.L.; Livingstone, D.S.; Schnell, R.J.; Gutiérrez, O.A. Genetic Characterization of the Cacao Cultivar CCN 51: Its Impact and Significance on Global Cacao Improvement and Production. J. Am. Soc. Hortic. Sci. 2014, 139, 219–229. [Google Scholar] [CrossRef]
- Cullen, J.T. On the Nonlinear Relationship between Dissolved Cadmium and Phosphate in the Modern Global Ocean: Could Chronic Iron Limitation of Phytoplankton Growth Cause the Kink? Limnol. Oceanogr. 2006, 51, 1369–1380. [Google Scholar] [CrossRef]
- Caldelas, C.; Poitrasson, F.; Viers, J.; Araus, J. Stable Zn Isotopes Reveal the Uptake and Toxicity of Zinc Oxide Engineered Nanomaterials in Phragmites australis. Environ. Sci. Nano 2020, 7, 1927–1941. [Google Scholar] [CrossRef]
- Jiang, Y.; Chen, X.; Chai, S.; Sheng, H.; Sha, L.; Fan, X.; Zeng, J.; Kang, H.; Zhang, H.; Xiao, X.; et al. TpIRT1 from Polish Wheat (Triticum polonicum L.) Enhances the Accumulation of Fe, Mn, Co, and Cd in Arabidopsis. Plant Sci. 2021, 312, 111058. [Google Scholar] [CrossRef] [PubMed]
- Ullah, I.; Wang, Y.; Eide, D.J.; Dunwell, J.M. Evolution, and Functional Analysis of Natural Resistance -Associated Macrophage Proteins (NRAMPs) from Theobroma cacao and Their Role in Cadmium Accumulation. Sci. Rep. 2018, 8, 14412. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Beyer, A.; Aebersold, R. On the Dependency of Cellular Protein Levels on mRNA Abundance. Cell 2016, 165, 535–550. [Google Scholar] [CrossRef]
- Dong, C.; He, F.; Berkowitz, O.; Liu, J.; Cao, P.; Tang, M.; Shi, H.; Wang, W.; Li, Q.; Shen, Z.; et al. Alternative Splicing Plays a Critical Role in Maintaining Mineral Nutrient Homeostasis in Rice (Oryza sativa). Plant Cell 2018, 30, 2267. [Google Scholar] [CrossRef]
- He, F.; Liu, Q.; Zheng, L.; Cui, Y.; Shen, Z.; Zheng, L. RNA-seq Analysis of Rice Roots Reveals the Involvement of Post-Transcriptional Regulation in Response to Cadmium Stress. Front. Plant Sci. 2015, 6, 1136. [Google Scholar] [CrossRef] [PubMed]
- Jian, M.; Zhang, D.; Wang, X.; Wei, S.; Zhao, Y.; Ding, Q.; Han, Y.; Ma, L. Differential Expression Pattern of the Proteome in Response to Cadmium Stress Based on Proteomics Analysis of Wheat Roots. BMC Genom. 2020, 21, 343. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Weckwerth, W. Mass Spectrometry Untangles Plant Membrane Protein Signaling Networks. Trends Plant Sci. 2020, 25, 930–944. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Wang, P.; Wang, P.; Yang, M.; Lian, X.; Tang, Z.; Huang, C.-F.; Salt, D.E.; Zhao, F.J. A Loss-of-Function Allele of OsHMA3 Associated with High Cadmium Accumulation in Shoots and Grain of Japonica Rice Cultivars. Plant Cell Environ. 2016, 39, 1941–1954. [Google Scholar] [CrossRef] [PubMed]
- Clemens, S. Toxic Metal Accumulation, Responses to Exposure and Mechanisms of Tolerance in Plants. Biochimie 2006, 88, 1707–1719. [Google Scholar] [CrossRef]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Ogo, Y.; Senoura, T.; Nishizawa, N.K.; Nakanishi, H. The OsHMA2 Transporter Is Involved in Root-to-Shoot Translocation of Zn and Cd in Rice. Plant. Cell Environ. 2012, 35, 1948–1957. [Google Scholar] [CrossRef]
- Andresen, E.; Peiter, E.; Küpper, H. Trace Metal Metabolism in Plants. J. Exp. Bot. 2018, 69, 909–954. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T | Nutrient Solutions | [Cd] | Cd | δ114Cd | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| (mg/kg, ± sd) | (μg, ± sd) | (‰, ± 2sd) | ||||||||
| Hydroponic | Foliar | Roots | Stems | Leaves | Total | Roots | Stems | Leaves | Total | |
| 1 | Fe, Zn | - | 322 ± 67 | 284 ± 12 | 115 ± 33 | 142 ± 9 | −0.74 ± 0.08 | −0.74 ± 0.04 | −0.48 ± 0.08 | −0.64 ± 0.04 |
| 2 | Fe, Zn | Fe | 348 ± 1 | 282 ± 73 | 159 ± 62 | 187 ± 38 | −0.73 ± 0.08 | −0.80 ± 0.02 | −0.58 ± 0.07 | −0.67 ± 0.04 |
| 3 | Fe, Zn | Zn | 352 ± 34 | 253 ± 17 | 109 ± 2 | 141 ± 24 | −0.61 ± 0.08 | −0.76 ± 0.09 | −0.59 ± 0.08 | −0.66 ± 0.05 |
| 4 | Zn | - | 482 ± 14 | 160 ± 23 | 37 ± 6 | 82 ± 14 | −0.70 ± 0.07 | −0.61 ± 0.06 | −0.38 ± 0.08 | −0.62 ± 0.05 |
| 5 | Zn | Fe | 486 ± 6 | 207 ± 31 | 47 ± 19 | 101 ± 0 | −0.74 ± 0.07 | −0.65 ± 0.04 | −0.50 ± 0.09 | −0.67 ± 0.04 |
| 6 * | Fe | - | 377 ± 32 | 240 ± 106 | 69 ± 42 | 118 ± 68 | −0.61 ± 0.05 | −0.77 ± 0.05 | −0.45 ± 0.09 | −0.63 ± 0.04 |
| 7 | Fe | Zn | 316 ± 61 | 308 ± 93 | 90 ± 33 | 122 ± 27 | −0.65 ± 0.07 | −0.80 ± 0.09 | −0.49 ± 0.10 | −0.64 ± 0.05 |
| 8 | - | - | 374 ± 47 | 93 ± 61 | 27 ± 28 | 59 ± 15 | −0.68 ± 0.13 | −0.64 ± 0.09 | −0.43 ± 0.05 | −0.61 ± 0.10 |
| Mean | 382 | 228 | 82 | 119 | −0.68 | −0.72 | −0.49 | −0.64 | ||
| sd | 66 | 72 | 45 | 40 | 0.05 | 0.07 | 0.07 | 0.02 | ||
| Min | 316 | 93 | 27 | 59 | −0.74 | −0.80 | −0.59 | −0.67 | ||
| Max | 486 | 308 | 159 | 187 | −0.61 | −0.61 | −0.38 | −0.61 | ||
| T | Nutrient Solutions | Δ114Cd (‰) | ε114Cd (‰) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Hydroponic | Foliar | L-S | S-R | L-R | Tot-Sol | L-Tot | Sh-Tot | Seq-Mob | |
| 1 | Fe, Zn | - | 0.28 | 0.00 | 0.25 | −0.29 | 0.17 | 0.03 | −0.15 |
| 2 | Fe, Zn | Fe | 0.22 | −0.07 | 0.14 | −0.31 | 0.09 | 0.02 | −0.12 |
| 3 | Fe, Zn | Zn | 0.16 | −0.16 | 0.01 | −0.28 | 0.04 | −0.02 | −0.10 |
| 4 | Zn | - | 0.23 | 0.07 | 0.31 | −0.25 | 0.23 | 0.09 | −0.15 |
| 5 | Zn | Fe | 0.14 | 0.09 | 0.24 | −0.31 | 0.17 | 0.07 | −0.10 |
| 6 | Fe | - | 0.27 | −0.09 | 0.19 | −0.26 | 0.16 | 0.03 | −0.15 |
| 7 | Fe | Zn | 0.27 | −0.15 | 0.14 | −0.29 | 0.13 | 0.00 | −0.11 |
| 8 | - | - | 0.27 | 0.09 | 0.23 | −0.21 | 0.13 | 0.07 | −0.11 |
| Mean | 0.23 | −0.03 | 0.19 | −0.27 | 0.14 | 0.04 | −0.12 | ||
| sd | 0.05 | 0.11 | 0.09 | 0.03 | 0.05 | 0.04 | 0.02 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moore, R.E.T.; Ullah, I.; Dunwell, J.M.; Rehkämper, M. Stable Isotope Analyses Reveal Impact of Fe and Zn on Cd Uptake and Translocation by Theobroma cacao. Plants 2024, 13, 551. https://doi.org/10.3390/plants13040551
Moore RET, Ullah I, Dunwell JM, Rehkämper M. Stable Isotope Analyses Reveal Impact of Fe and Zn on Cd Uptake and Translocation by Theobroma cacao. Plants. 2024; 13(4):551. https://doi.org/10.3390/plants13040551
Chicago/Turabian StyleMoore, Rebekah E. T., Ihsan Ullah, Jim M. Dunwell, and Mark Rehkämper. 2024. "Stable Isotope Analyses Reveal Impact of Fe and Zn on Cd Uptake and Translocation by Theobroma cacao" Plants 13, no. 4: 551. https://doi.org/10.3390/plants13040551
APA StyleMoore, R. E. T., Ullah, I., Dunwell, J. M., & Rehkämper, M. (2024). Stable Isotope Analyses Reveal Impact of Fe and Zn on Cd Uptake and Translocation by Theobroma cacao. Plants, 13(4), 551. https://doi.org/10.3390/plants13040551