
Polyethylene Glycol Priming Enhances the Seed Germination and Seedling Growth of Scutellaria baicalensis Georgi under Salt Stress

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. PEG Priming Significantly Alleviates the Germination Inhibition of S. baicalensis Seeds under Salt Stress
2.2. PEG Priming Significantly Enhanced the Growth of S. baicalensis Seedlings under Salt Stress
2.3. Effect of PEG Priming on Osmolyte Content and Relative Water Content of S. baicalensis Shoots under Salt Stress
2.4. Effect of PEG Priming on Lipid Peroxidation and Membrane Permeability of S. baicalensis Shoots under Salt Stress
2.5. Effect of PEG Priming on Antioxidant Systems of S. baicalensis Shoots under Salt Stress
2.6. Effect of PEG Priming on Contents of Sodium and Potassium Ions in S. baicalensis Shoots under Salt Stress
2.7. Effect of PEG Priming on Transcript Levels of Antioxidant-Related Genes in S. baicalensis Shoots under Salt Stress
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Priming
4.2. Germination Test
4.3. Measurements of Growth Parameters and Sampling
4.4. Assay of Physiological Indicators and Ion Contents
4.5. qRT-PCR Analysis
4.6. Statistical Analysis
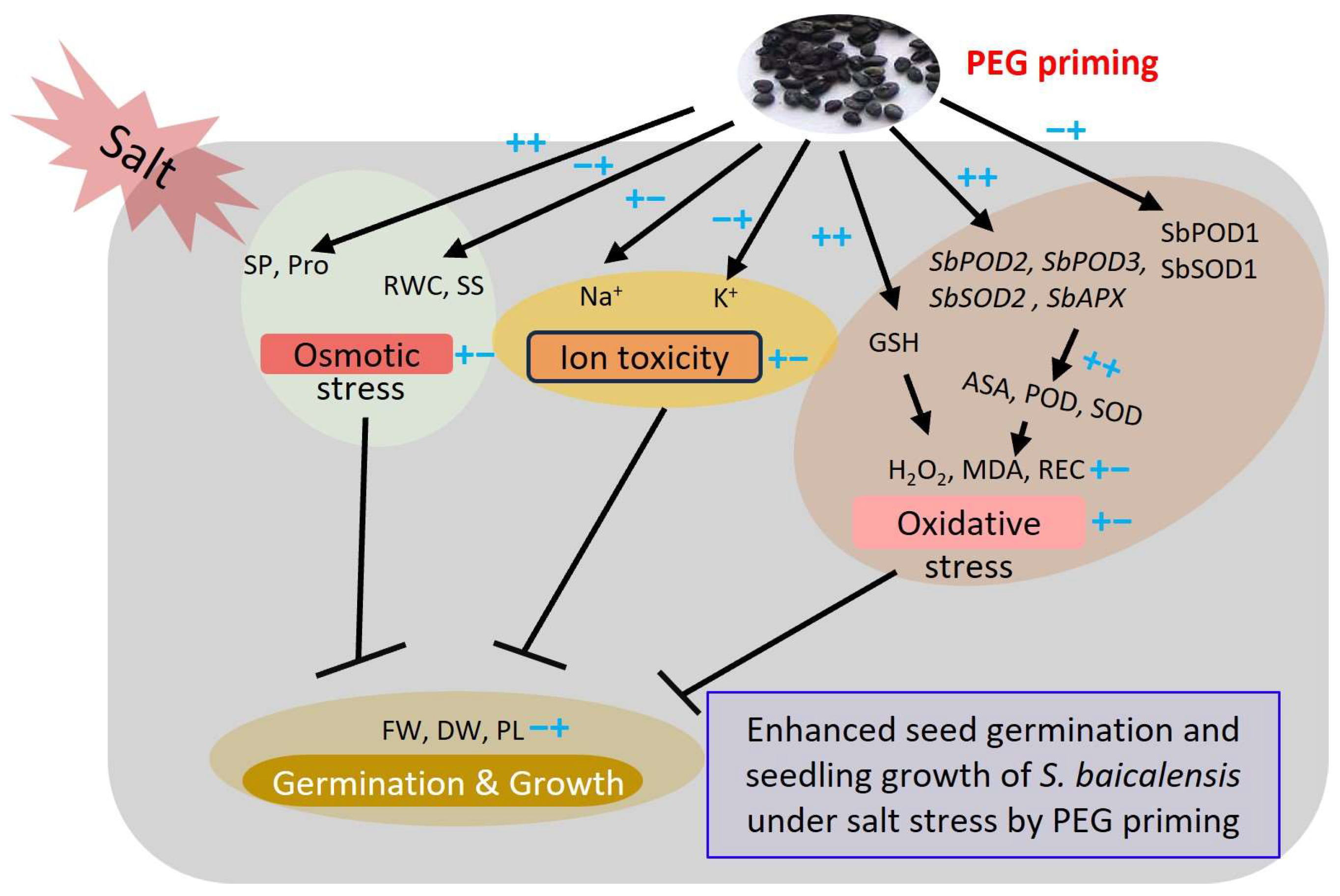
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Haj-Amor, Z.; Araya, T.; Kim, D.G.; Bouri, S.; Lee, J.; Ghiloufi, W.; Yang, Y.; Kang, H.; Jhariya, M.K.; Banerjee, A.; et al. Soil salinity and its associated effects on soil microorganisms, greenhouse gas emissions, crop yield, biodiversity and desertification: A review. Sci. Total Environ. 2022, 843, 156946. [Google Scholar] [CrossRef]
- Ivushkin, K.; Bartholomeus, H.; Bregt, A.K.; Pulatov, A.; Sousa, L.D. Global mapping of soil salinity change. Remote Sens. Environ. 2019, 231, 111260. [Google Scholar] [CrossRef]
- Wang, J.L.; Huang, X.J.; Zhong, T.Y.; Chen, Z.G. Review on sustainable utilization of salt-affected land. Acta Geogr. Sin. 2011, 66, 673–684. [Google Scholar]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guo, Y. Elucidating the molecular mechanisms mediating plant salt-stress responses. New Phytol. 2018, 217, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Abdelaal, K.; Alsubeie, M.S.; Hafez, Y.; Emeran, A.; Moghanm, F.; Okasha, S.; Omara, R.; Basahi, M.A.; Darwish, D.B.E.; Ibrahim, M.F.M.; et al. Physiological and biochemical changes in vegetable and field crops under drought, salinity and weeds stresses: Control strategies and management. Agriculture 2022, 12, 2084. [Google Scholar] [CrossRef]
- Biswas, S.; Seal, P.; Majumder, B.; Biswas, A.K. Efficacy of seed priming strategies for enhancing salinity tolerance in plants: An overview of the progress and achievements. Plant Stress 2023, 9, 100186. [Google Scholar] [CrossRef]
- Ibrahim, E.A. Seed priming to alleviate salinity stress in germinating seeds. J. Plant Physiol. 2016, 192, 38–46. [Google Scholar] [CrossRef]
- Li, J.; Zhao, M.A.; Liu, L.G.; Guo, X.M.; Pei, Y.H.; Wang, C.X.; Song, X.Y. Exogenous sorbitol application confers drought tolerance to maize seedlings through up-regulating antioxidant system and endogenous sorbitol biosynthesis. Plants 2023, 12, 2456. [Google Scholar]
- Bradford, K.J. Manipulation of seed water relations via osmotic priming to improve germination under stress conditions. HortScience 1986, 21, 1105–1112. [Google Scholar] [CrossRef]
- Yan, M. Seed priming stimulate germination and early seedling growth of Chinese cabbage under drought stress. S. Afr. J. Bot. 2015, 99, 88–92. [Google Scholar] [CrossRef]
- Alam, A.; Ullah, H.; Thuenprom, N.; Tisarum, R.; Cha-um, S.; Datta, A. Seed priming with salicylic acid enhances growth, physiological traits, fruit yield, and quality parameters of cantaloupe under water-deficit stress. S. Afr. J. Bot. 2022, 150, 1–12. [Google Scholar] [CrossRef]
- Benadjaoud, A.; Dadach, M.; El-Keblawy, A.; Mehdadi, Z. Impacts of osmopriming on mitigation of the negative effects of salinity and water stress in seed germination of the aromatic plant Lavandula stoechas L. J. Appl. Res. Med. Aromat. Plants 2022, 31, 100407. [Google Scholar] [CrossRef]
- Sako, K.; Nguyen, H.M.; Seki, M. Advances in chemical priming to enhance abiotic stress tolerance in plants. Plant Cell Physiol. 2021, 61, 1995–2003. [Google Scholar] [CrossRef]
- Zhou, H.; Shi, H.; Yang, Y.; Feng, X.; Chen, X.; Xiao, F.; Lin, H.; Guo, Y. Insights into plant salt stress signaling and tolerance. J. Genet. Genom. 2023, 51, 16–34. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hou, J.; Jiang, C.; Li, G.; Lu, H.; Meng, F.; Shi, L. Deep sequencing of the Scutellaria baicalensis Georgi transcriptome reveals flavonoid biosynthetic profiling and organ-specific gene expression. PLoS ONE 2015, 10, e0136397. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, W.; Wang, D.; Hu, S.; Zhang, Q.; Wang, Z.; Cui, L. Genome-wide identification and characterization of the WRKY gene family in Scutellaria baicalensis Georgi under diverse abiotic stress. Int. J. Mol. Sci. 2022, 23, 4225. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Hu, S.; Cao, Y.; Chen, R.; Wang, Z.; Cao, X. Selection and evaluation of reference genes for qRT-PCR of Scutellaria baicalensis Georgi under different experimental conditions. Mol. Biol. Rep. 2021, 48, 1115–1126. [Google Scholar] [CrossRef]
- Cai, Z.; Liu, X.; Chen, H.; Yang, R.; Chen, J.; Zou, L.; Wang, C.; Chen, J.; Tan, M.; Mei, Y.; et al. Variations in morphology, physiology, and multiple bioactive constituents of Lonicerae Japonicae Flos under salt stress. Sci. Rep. 2021, 11, 3939. [Google Scholar] [CrossRef]
- Cayuela, E.; Perezalfocea, F.; Caro, M.; Bolarin, M.C. Priming of seeds with NaCl induces physiological changes in tomato plants grown under salt stress. Physiol. Plant. 1996, 96, 231–236. [Google Scholar] [CrossRef]
- Rubinstein, B. Regulation of H+ excretion: Effects of osmotic shock. Plant Physiol. 1982, 69, 939–944. [Google Scholar] [CrossRef] [PubMed]
- Amini, R. Drought stress tolerance of barley (Hordeum vulgare L.) affected by priming with PEG. Int. J. Farming Allied Sci. 2013, 2, 803–808. [Google Scholar]
- Al-Shammari, W.B.; Altamimi, H.R.; Abdelaal, K. Improvement in physiobiochemical and yield characteristics of Pea plants with nano-silica and melatonin under salinity stress conditions. Horticulturae 2023, 9, 711. [Google Scholar] [CrossRef]
- Shabala, S. Learning from halophytes: Physiological basis and strategies to improve abiotic stress tolerance in crops. Ann. Bot. 2013, 112, 1209–1221. [Google Scholar] [CrossRef]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [PubMed]
- AlKahtani, M.D.F.; Hafez, Y.M.; Attia, K.; Rashwan, E.; Husnain, L.A.; AlGwaiz, H.I.M.; Abdelaal, K.A.A. Evaluation of silicon and proline application on the oxidative machinery in drought-stressed sugar beet. Antioxidants 2021, 10, 398. [Google Scholar] [CrossRef]
- Yang, S.Y.; Hao, D.L.; Jin, M.; Li, Y.; Liu, Z.T.; Huang, Y.N.; Chen, T.X.; Su, Y.H. Internal ammonium excess induces ROS-mediated reactions and causes carbon scarcity in rice. BMC Plant Biol. 2020, 20, 143. [Google Scholar] [CrossRef]
- Wang, Q.; Peng, X.; Lang, D.; Ma, X.; Zhang, X. Physio-biochemical and transcriptomic analysis reveals that the mechanism of Bacillus cereus G2 alleviated oxidative stress of salt-stressed Glycyrrhiza uralensis Fisch. seedlings. Ecotoxicol. Environ. Saf. 2022, 247, 114264. [Google Scholar] [CrossRef]
- Vazayefi, M.; Shekari, F.; Zangani, E.; Dolatabadian, A.; Janda, T.; Mastinu, A. Seed treatment with chlormequat chloride improves the physiological and biochemical characteristics of Brassica napus L. under salt stress. Plant Stress 2023, 9, 100175. [Google Scholar] [CrossRef]
- Yue, Y.; Zhang, M.C.; Zhang, J.C.; Duan, L.S.; Li, Z.H. SOS1 gene overexpression increased salt tolerance in transgenic tobacco by maintaining a higher K+/Na+ ratio. J. Plant Physiol. 2012, 169, 255–261. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signaling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, C.; Liang, Y.; Wang, C.; Yang, C.; Liu, G. A novel bZIP gene from Tamarix hispida mediates physiological responses to salt stress in tobacco plants. J. Plant Physiol. 2010, 167, 222–330. [Google Scholar] [CrossRef]
- Ashraf, M. Biotechnological approach of improving plant salt tolerance using antioxidants as markers. Biotechnol. Adv. 2009, 27, 84–93. [Google Scholar] [CrossRef]
- Yusuf, C.S.; Chand, R.; Mishra, V.K.; Joshi, A.K. The association between leaf malondialdehyde and lignin content and re-sistance to spot blotch in wheat. J. Phytopathol. 2016, 164, 896–903. [Google Scholar] [CrossRef]
- Aminian, M.; Nabatchian, F.; Vaisi-Raygani, A.; Torabi, M. Mechanism of Coomassie Brilliant Blue G-250 binding to cetyltrimethylammonium bromide: An interference with the Bradford assay. Anal. Biochem. 2013, 434, 287–291. [Google Scholar] [CrossRef]
- Hackmann, T.J.; Keyser, B.L.; Firkins, J.L. Evaluation of methods to detect changes in reserve carbohydrate for mixed rumen microbes. J. Microbiol. Methods 2013, 93, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Benitez, L.C.; Vighi, I.L.; Auler, P.A.; Amaral, M.N.; Moraes, G.P.; Rodrigues, G.S.; Maia, L.C.; Júnior, A.M.; Braga, E.J. Correlation of proline content and gene expression involved in the metabolism of this amino acid under abiotic stress. Acta Physiol. Plant. 2016, 38, 267. [Google Scholar] [CrossRef]
- Martinez-Gutierrez, R.; Mora-Herrera, M.E.; Lopez-Delgado, H.A. Exogenous H2O2 in phytoplasma-infected potato plants promotes antioxidant activity and tuber production under drought conditions. Am. J. Potato Res. 2012, 89, 53–62. [Google Scholar] [CrossRef]
- Usluoglu, A.; Arabaci, G. The effect of acid dyes on antioxidant enzymes from cress (Lepidum sativum). Curr. Opin. Biotechnol. 2011, 22, 140. [Google Scholar] [CrossRef]
- Williams, V.; Twine, S. Flame photometric method for sodium, potassium and calcium. In Modern Methods of Plant Analysis; Peach, K., Tracey, M.V., Eds.; Springer-Verlag: Berlin/Heidelberg, Germany, 1960; pp. 3–5. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; Li, C.; Zeng, L.; Liu, L.; Xi, J.; Li, J. Polyethylene Glycol Priming Enhances the Seed Germination and Seedling Growth of Scutellaria baicalensis Georgi under Salt Stress. Plants 2024, 13, 565. https://doi.org/10.3390/plants13050565
Wang R, Li C, Zeng L, Liu L, Xi J, Li J. Polyethylene Glycol Priming Enhances the Seed Germination and Seedling Growth of Scutellaria baicalensis Georgi under Salt Stress. Plants. 2024; 13(5):565. https://doi.org/10.3390/plants13050565
Chicago/Turabian StyleWang, Renjie, Chenxuan Li, Li Zeng, Ligong Liu, Jiayi Xi, and Jun Li. 2024. "Polyethylene Glycol Priming Enhances the Seed Germination and Seedling Growth of Scutellaria baicalensis Georgi under Salt Stress" Plants 13, no. 5: 565. https://doi.org/10.3390/plants13050565
APA StyleWang, R., Li, C., Zeng, L., Liu, L., Xi, J., & Li, J. (2024). Polyethylene Glycol Priming Enhances the Seed Germination and Seedling Growth of Scutellaria baicalensis Georgi under Salt Stress. Plants, 13(5), 565. https://doi.org/10.3390/plants13050565