


Isolation and Identification of Allelopathic Substances from Forsythia suspensa Leaves, and Their Metabolism and Activity


Abstract
:1. Introduction
2. Results
2.1. Allelopathic Activity of the F. suspensa Leaves
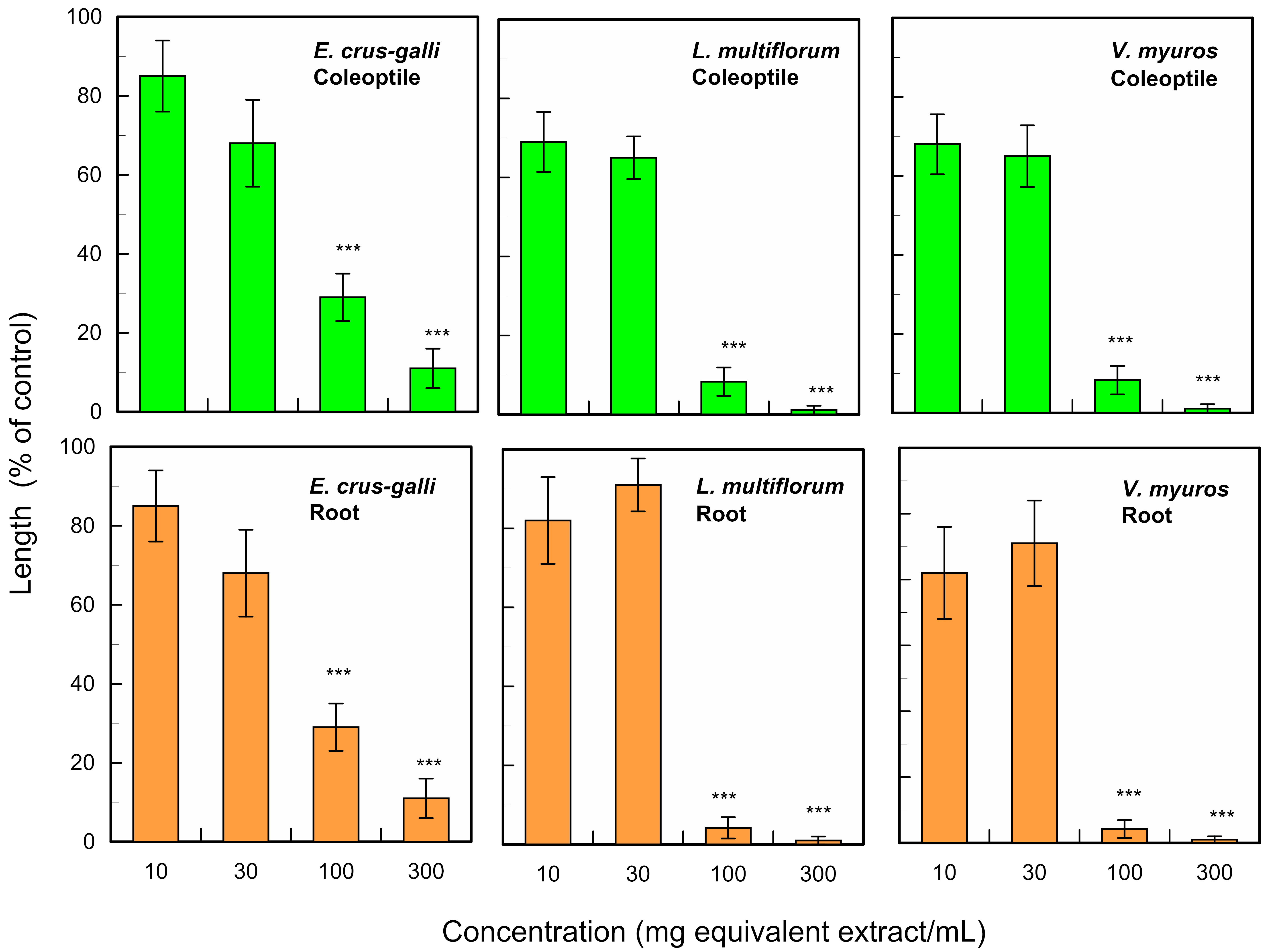
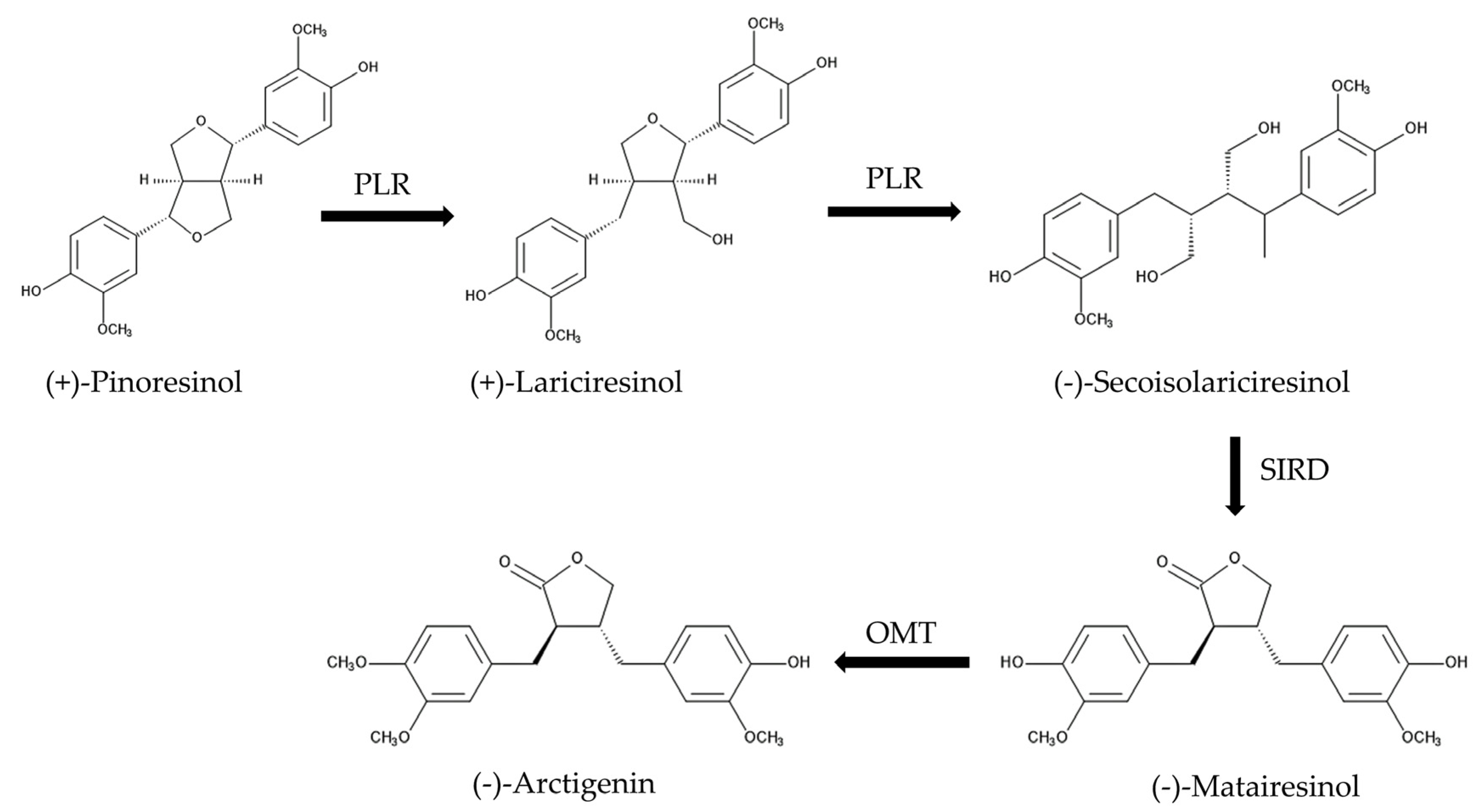
2.2. Purification and Identification of the Allelochemicals in the Leaves
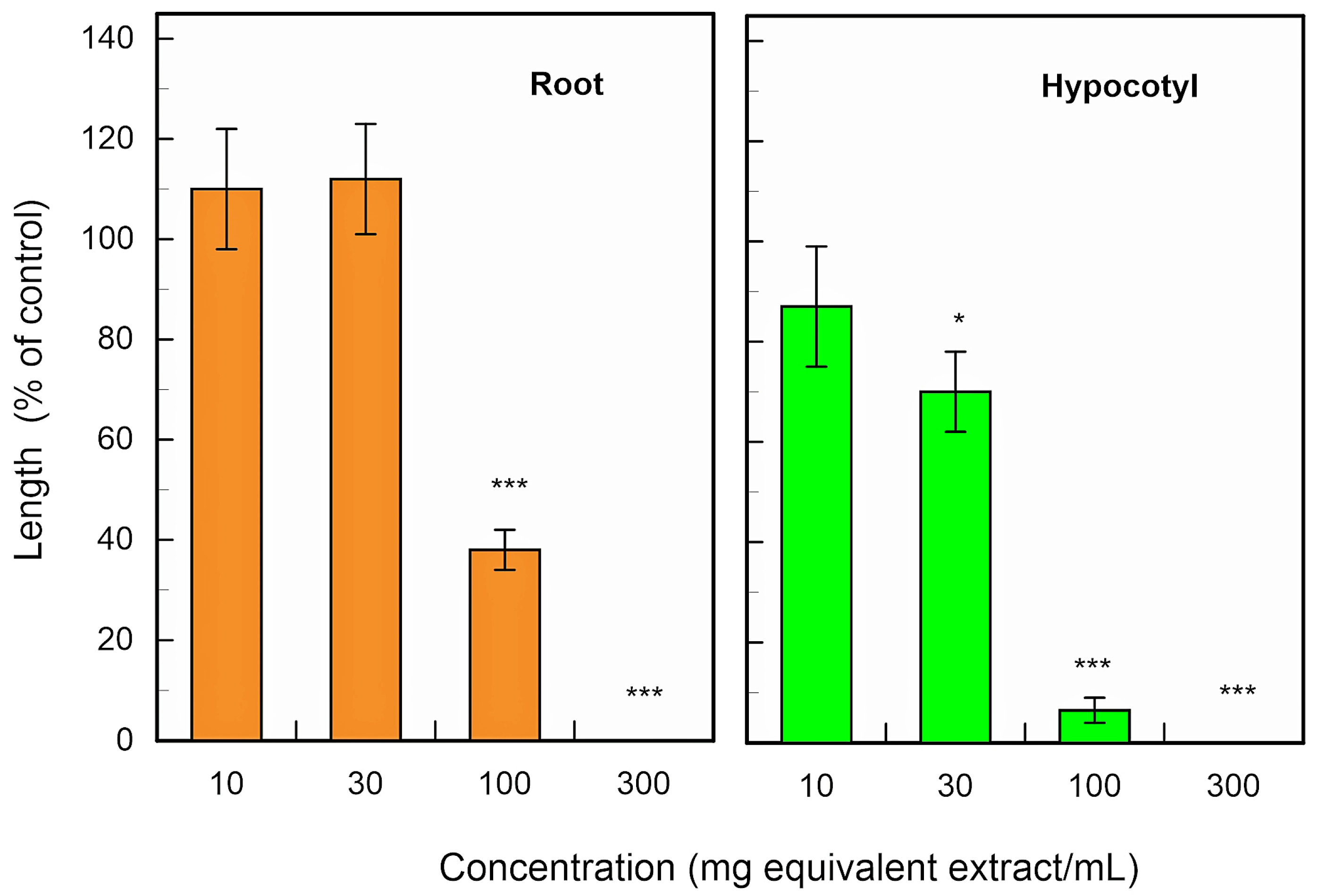
2.3. Allelopathic Activity of the Isolated Compounds and Pinolesinol
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Extraction and Determination of the Allelopathic Activity of F. suspensa
4.3. Separation of the F. suspensa Extract
4.4. Allelopathic Activity of the Isolated Compounds and Pinoresinol
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Z.; Xia, Q.; Liu, X.; Liu, W.; Huang, W.; Mei, X.; Luo, J.; Shan, M.; Lin, R.; Zou, D.; et al. Phytochemistry, pharmacology, quality control and future research of Forsythia suspensa (Thunb.) Vahl: A review. J. Ethnopharmacol. 2018, 210, 318–339. [Google Scholar] [CrossRef] [PubMed]
- Makino, T. Observation on the flora of Japan. Bot. Mag. 1912, 28, 105–107. [Google Scholar] [CrossRef] [PubMed]
- Leshcheniuk, O.; Koniakin, S. Morphological and anatomical parameters of Forsythia suspensa (Thunb.) Vahl leaves in the urban environment. Plant Introd. 2022, 95/96, 75–84. [Google Scholar] [CrossRef]
- Iwatsubo, Y.; Sato, K.; Miura, N.; Ohno, M.; Fujino, H.; Tatsuo, Y.; Sugiyama, R.; Kurosaki, F. Chromosome numbers of the genus Forsythia (Oleaceae). J. Phytogeogr. Taxon. 2008, 56, 95–97. [Google Scholar]
- Shen, J.S.; Xie, Y.J.; Pan, H.T.; Zhang, Q.X. Karyotype analysis of twelve Forsythia species and cultivars. J. Plant Genet. Resour. 2015, 16, 178–184. [Google Scholar]
- Yang, J.; Miao, C.Y.; Mao, R.L.; Li, Y. Landscape population genomics of Forsythia (Forsythia suspensa) reveal that ecological habitats determine the adaptive evolution of species. Front. Plant Sci. 2017, 8, 481. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Lu, P.; Liu, Y.; Liu, C.; Gao, Y.; Nwafor, E.O.; Zhang, Y.; Liu, Z. Research progress on chemical composition, pharmacological effects of Forsythia suspensa (Thunb.) Vahl and predictive analysis on Q-marker. TMR Mod. Herb. Med. 2020, 3, 86–112. [Google Scholar] [CrossRef]
- Bailly, C. Forsythosides as essential components of Forsythia-based traditional Chinese medicines used to treat inflammatory diseases and COVID-19. World J. Traditi. Chin. Med. 2022, 8, 1–20. [Google Scholar] [CrossRef]
- Gong, L.; Wang, C.; Zhou, H.; Ma, C.; Zhang, Y.; Peng, C.; Li, Y. A review of pharmacological and pharmacokinetic properties of Forsythiaside A. Pharmacolo. Res. 2021, 169, 105690. [Google Scholar] [CrossRef]
- Plants of the World. Royal Botanical Gardens—Kew, Mimosa pigra L. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:608900-1 (accessed on 17 December 2023).
- Gardener’s Path. Forsythia suspensa. Available online: https://gardenerspath.com/plants/ornamentals/grow-weeping-forsythia/ (accessed on 17 December 2023).
- Hortipedia. Forsythia suspensa. Available online: https://en.hortipedia.com/Forsythia_suspensa (accessed on 17 December 2023).
- Plant for a Furture. Forsythia suspensa. Available online: https://pfaf.org/User/Plant.aspx?LatinName=Forsythia+suspensa (accessed on 17 December 2023).
- Fujii, Y.; Shibuya, T.; Yasuda, T. Survey of Japanese weed and crops for the detection of water-extractable allelopathic chemicals using Richards’ function fitted to lettuce germination test. Weed Res. Jpn. 1990, 35, 362–370. [Google Scholar]
- Fujii, Y.; Parvez, S.S.; Parvez, M.M.; Ohmae, Y.; Iida, O. Screening of 239 medicinal plant species for allelopathic activity using the sandwich method. Weed Biol. Manag. 2003, 3, 233–241. [Google Scholar] [CrossRef]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: Orlando, FL, USA, 1984; pp. 1–422. [Google Scholar]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef]
- Bonanomi, G.; Sicurezza, M.G.; Caporaso, S.; Esposito, A.; Mazzoleni, S. Phytotoxicity dynamics of decaying plant materials. New Phytol. 2006, 169, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Kato-Noguchi, H. Defensive molecules momilactones A and B: Function, biosynthesis, induction and occurrence. Toxins 2023, 15, 241. [Google Scholar] [CrossRef] [PubMed]
- Belz, R.G. Allelopathy in crop/weed interactions—An update. Pest Manag. Sci. 2007, 63, 308–326. [Google Scholar] [CrossRef] [PubMed]
- Macías, F.A.; Molinillo, J.M.G.; Varela, R.M.; Galindo, J.G.G. Allelopathy—A natural alternative for weed control. Pest Manag. Sci. 2007, 63, 327–348. [Google Scholar] [CrossRef] [PubMed]
- Narwal, S.S. Allelopathy in weed management. In Allelopathy Update. Basic and Applied Aspects; Narwal, S.S., Ed.; Science Publishers Inc.: Enfield, NH, USA, 1999; Volume 2, pp. 203–254. [Google Scholar]
- Duke, S.O.; Dayan, F.E.; Romagni, J.G.; Rimando, A.M. Natural products as sources of herbicide, current status and future trends. Weed Res. 2000, 40, 99–111. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Phytotoxic substances involved in teak allelopathy and agroforestry. Appl. Sci. 2021, 11, 3314. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kato, M. Evolution of the secondary metabolites in invasive plant species Chromolaena odorata for the defense and allelopathic functions. Plants 2023, 12, 521. [Google Scholar] [CrossRef]
- Niemeyer, H.B.; Honig, D.; Lange-Böhmer, A.; Jacobs, E.; Kulling, S.E.; Metzler, M. Oxidative metabolites of the mammalian lignans enterodiol and enterolactone in rat bile and urine. J. Agric. Food Chem. 2000, 48, 2910–2919. [Google Scholar] [CrossRef]
- Fischer, J.; Reynolds, A.J.; Sharp, L.A.; Sherburn, M.S. Radical carboxyarylation approach to lignans. Total synthesis of (−)-arctigenin, (−)-matairesinol, and related natural products. Org. Lett. 2004, 6, 1345–1348. [Google Scholar] [CrossRef] [PubMed]
- Mallik, A.U. Allelopathy and competition in coniferous forests. Environ. For. Sci. 1998, 54, 309–315. [Google Scholar]
- Kato-Noguchi, H.; Kimura, F.; Ohno, O.; Suenaga, K. Involvement of allelopathy in inhibition of understory growth in red pine forests. J. Plant Physiol. 2017, 218, 66–73. [Google Scholar] [CrossRef]
- Matuda, Y.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Allelopathy and allelopathic substances of fossil tree species Metasequoia glyptostroboides. Agronomy 2022, 12, 83. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kurniadie, D. Allelopathy and allelochemicals of Leucaena leucocephala as an invasive plant species. Plants 2022, 11, 1672. [Google Scholar] [CrossRef] [PubMed]
- Kato-Noguchi, H.; Kato, M. Allelopathy and allelochemicals of Solidago canadensis L. and S. altissima L. for their naturalization. Plants 2022, 11, 3235. [Google Scholar] [CrossRef] [PubMed]
- Kato-Noguchi, H. Involvement of allelopathy in the invasive potential of Tithonia diversifolia. Plants 2020, 9, 766. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Allelopathy and allelochemicals of Imperata cylindrica as an invasive plant species. Plants 2022, 11, 2551. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Hamada, Y.; Kojima, M.; Kumagai, S.; Iwasaki, A.; Suenaga, K. Allelopathic substances of Osmanthus spp. for developing sustainable agriculture. Plants 2023, 12, 376. [Google Scholar] [CrossRef]
- Lewis, N.G.; Davin, L.B. Lignans: Biosynthesis and function. In Comprehensive Natural Products Chemistry, I; Barton, S.D., Nakanishi, K., Meth-Cohn, O., Eds.; Elsevier: Amsterdam, The Netherlands, 1999; pp. 639–712. [Google Scholar]
- Ono, E.; Kim, H.J.; Murata, J.; Morimoto, K.; Okazawa, A.; Kobayashi, A.; Umezawa, T.; Satake, H. Molecular and functional characterization of novel furofuran-class lignan glucosyltransferases from Forsythia. Plant Botechnol. 2010, 27, 317–324. [Google Scholar] [CrossRef]
- Schmitt, J.; Petersen, M. Pinoresinol and matairesinol accumulation in a Forsythia × intermedia cell suspension culture. Plant Cell Tissue Organ Cult. 2002, 68, 91–98. [Google Scholar] [CrossRef]
- Ward, R.S. Recent advances in the chemistry of lignans. In Studies in Natural Products Chemistry; Attaur, R., Ed.; Elsevier: Amsterdam, The Netherlands, 2000; pp. 739–798. [Google Scholar]
- Davin, L.B.; Lewis, N.G. A historical perspective on lignan biosynthesis: Monolignol, allylphenol and hydroxycinnamic acid coupling and downstream metabolism. Phytochem. Rev. 2003, 2, 257–288. [Google Scholar] [CrossRef]
- Ferrer, J.L.; Austin, M.B.; Stewart, C., Jr.; Noel, J.P. Structure and function of enzymes involved in the biosynthesis of phenyl-propanoids. Plant Physiol. Biochem. 2008, 46, 356–370. [Google Scholar] [CrossRef] [PubMed]
- Patyra, A.; Kołtun-Jasion, M.; Jakubiak, O.; Kiss, A.K. Extraction techniques and analytical methods for isolation and characterization of lignans. Plants 2022, 11, 2323. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, T.; Okano, M.; Noshita, T.; Funayama, S.; Ohtsuki, K. Characterization of (−)-matairesinol as a potent inhibitor of casein kinase I in vitro. Biol. Pharm. Bull. 2003, 26, 371–374. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Kim, K.H.; Jung, H.J.; Kwon, H.J. Matairesinol inhibits angiogenesis via suppression of mitochondrial reactive oxygen species. Biochem. Biophys. Res. Commun. 2012, 421, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Gandaraa, F.; Torresb, A.; Fernández-Poncec, M.T.; Casasc, L.; Mantellc, C.; Varela, R.; de la Ossa-Fernándezc, E.J.M.; Macías, F.A. Selective fractionation and isolation of allelopathic compounds from Helianthus annuus L. leaves by means of high-pressure techniques. J. Supercriti. Fluids 2019, 143, 32–41. [Google Scholar] [CrossRef]
- Zorrilla, J.G.; Cárdenas, D.M.; Rial, C.; Molinillo, J.M.; Varela, R.M.; Masi, M.; Macías, F.A. Bioprospection of phytotoxic plant-derived eudesmanolides and guaianolides for the control of Amaranthus viridis, Echinochloa crus-galli, and Lolium perenne weeds. J. Agric. Food Chem. 2024, 72, 1797–1810. [Google Scholar] [CrossRef]
- Heděnec, P.; Novotný, D.; Ust’ak, S.; Honzík, R.; Kovářová, M.; Šimáčková, H.; Frouz, J. Allelopathic effect of new introduced biofuel crops on the soil biota: A comparative study. Eur. J. Soil Biol. 2014, 63, 14–20. [Google Scholar] [CrossRef]
- Inderjit, S.; Dakshini, K.M.M. Investigations on some aspects of chemical ecology of cogongrass, Imperata cylindrica (L.) Beauv. J. Chem. Ecol. 1991, 17, 343–352. [Google Scholar] [CrossRef]
- Ishak, M.S.; Sahid, I. Allelopathic effects of the aqueous extract of the leaf and seed of Leucaena leucocephala on three selected weed species. AIP Conf. Proc. 2014, 1614, 659–664. [Google Scholar]
- Oke, S.O.; Awowoyin, A.V.; Osein, S.R.; Adediwura, E.L. Effects of aqueous shoot extract of Tithonia diversifolia on the growth of seedlings of Monodora tenuifolia (Benth.), Dialium guineense (Willd.) and Hildegardia barteri (Mast.) Kosterm. Not. Sci. Biol. 2011, 3, 64–70. [Google Scholar] [CrossRef]
- Zheng, H.; Wei, N.; Wang, L.; He, P. Effects of Lantana camara leaf extract on the activity of superoxide dismutase and accumulation of H2O2 in water hyacinth leaf. J. Plant Physiol. Mol. Biol. 2006, 32, 189–194. [Google Scholar]
- Poonpaiboonpipat, T.; Poolkum, S. Utilization of Bidens pilosa var. radiata (Sch. Bip.) Sherff integrated with water irrigation for paddy weed control and rice yield production. Weed Biol. Manag. 2019, 19, 31–38. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kurniadie, D. Allelopathy of Lantana camara as an invasive plant. Plants 2021, 10, 1028. [Google Scholar] [CrossRef] [PubMed]
- Kato-Noguchi, H. Invasive mechanisms of one of the world’s worst alien plant species Mimosa pigra and its management. Plants 2023, 12, 1960. [Google Scholar] [CrossRef] [PubMed]
- Kato-Noguchi, H. The impact and invasive mechanisms of Pueraria montana var. lobata, one of the world’s worst alien species. Plants 2023, 12, 3066. [Google Scholar] [CrossRef]
- Tominaga, T.; Watanabe, O. Weed growth suppression by cogongrass (Imperata cylindrica) leaves. Weed Sci. Tech. 1997, 42, 289–293. [Google Scholar] [CrossRef]
- Koger, C.H.; Bryson, C.T. Effect of cogongrass (Imperata cylindrica) extracts on germination and seedling growth of selected grass and broadleaf species. Weed Technol. 2004, 18, 236–242. [Google Scholar] [CrossRef]
- Chou, C.H.; Kuo, Y.L. Allelopathic research of subtropical vegetation in Taiwan. J. Chem. Ecol. 1986, 12, 1431–1448. [Google Scholar] [CrossRef]
- Wang, R.; Kang, X.; Quan, G.; Zhang, J. Influence of Lantana camara on soil II. Effects of Lantana camara leaf litter on plants and soil properties. Allelopath. J. 2015, 35, 207–216. [Google Scholar]
- Kato-Noguchi, H. Allelopathy of knotweeds as invasive plants. Plants 2022, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Tsyganova, A.V.; Brewin, N.J.; Tsyganov, V.E. Structure and development of the legume-rhizobial symbiotic interface in infection threads. Cells 2021, 10, 1050. [Google Scholar] [CrossRef]
- Mathesius, U. Are legumes different? Origins and consequences of evolving nitrogen fixing symbioses. J. Plant Physiol. 2022, 276, 153765. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008; pp. 1–815. [Google Scholar]
- Diagne, N.; Ngom, M.; Djighaly, P.I.; Fall, D.; Hocher, V.; Svistoonoff, S. Roles of arbuscular mycorrhizal fungi on plant growth and performance: Importance in biotic and abiotic stressed regulation. Diversity 2020, 12, 370. [Google Scholar] [CrossRef]
- Tang, H.; Hassan, M.U.; Feng, L.; Nawaz, M.; Shah, A.N.; Qari, S.H.; Liu, Y.; Miao, J. The critical role of arbuscular mycorrhizal fungi to improve drought tolerance and nitrogen use efficiency in crops. Front. Plant Sci. 2022, 13, 919166. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cress | L. multiflorum | |||
|---|---|---|---|---|
| Compound | Root | Hypocotyl | Root | Coleoptile |
| (-)-Matairesinol | 1.1 b | 2.1 b | 0.93 b | 2.2 b |
| (-)-Arctigenin | 0.79 a | 1.3 a | 0.85 a | 1.3 a |
| (+)-Pinoresinol | 2.1 c | 2.3 b | 1.1 c | 2.5 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kato-Noguchi, H.; Takahashi, Y.; Tojo, S.; Teruya, T. Isolation and Identification of Allelopathic Substances from Forsythia suspensa Leaves, and Their Metabolism and Activity. Plants 2024, 13, 575. https://doi.org/10.3390/plants13050575
Kato-Noguchi H, Takahashi Y, Tojo S, Teruya T. Isolation and Identification of Allelopathic Substances from Forsythia suspensa Leaves, and Their Metabolism and Activity. Plants. 2024; 13(5):575. https://doi.org/10.3390/plants13050575
Chicago/Turabian StyleKato-Noguchi, Hisashi, Yuga Takahashi, Shunya Tojo, and Toshiaki Teruya. 2024. "Isolation and Identification of Allelopathic Substances from Forsythia suspensa Leaves, and Their Metabolism and Activity" Plants 13, no. 5: 575. https://doi.org/10.3390/plants13050575
APA StyleKato-Noguchi, H., Takahashi, Y., Tojo, S., & Teruya, T. (2024). Isolation and Identification of Allelopathic Substances from Forsythia suspensa Leaves, and Their Metabolism and Activity. Plants, 13(5), 575. https://doi.org/10.3390/plants13050575