
Effect of the General Anaesthetic Ketamine on Electrical and Ca2+ Signal Propagation in Arabidopsis thaliana


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Anaesthetic Ketamine Modified Electrical Signals Mainly in Systemic Leaves
2.2. Anaesthetic Ketamine Did Not Block Ca2+ Wave Propagation
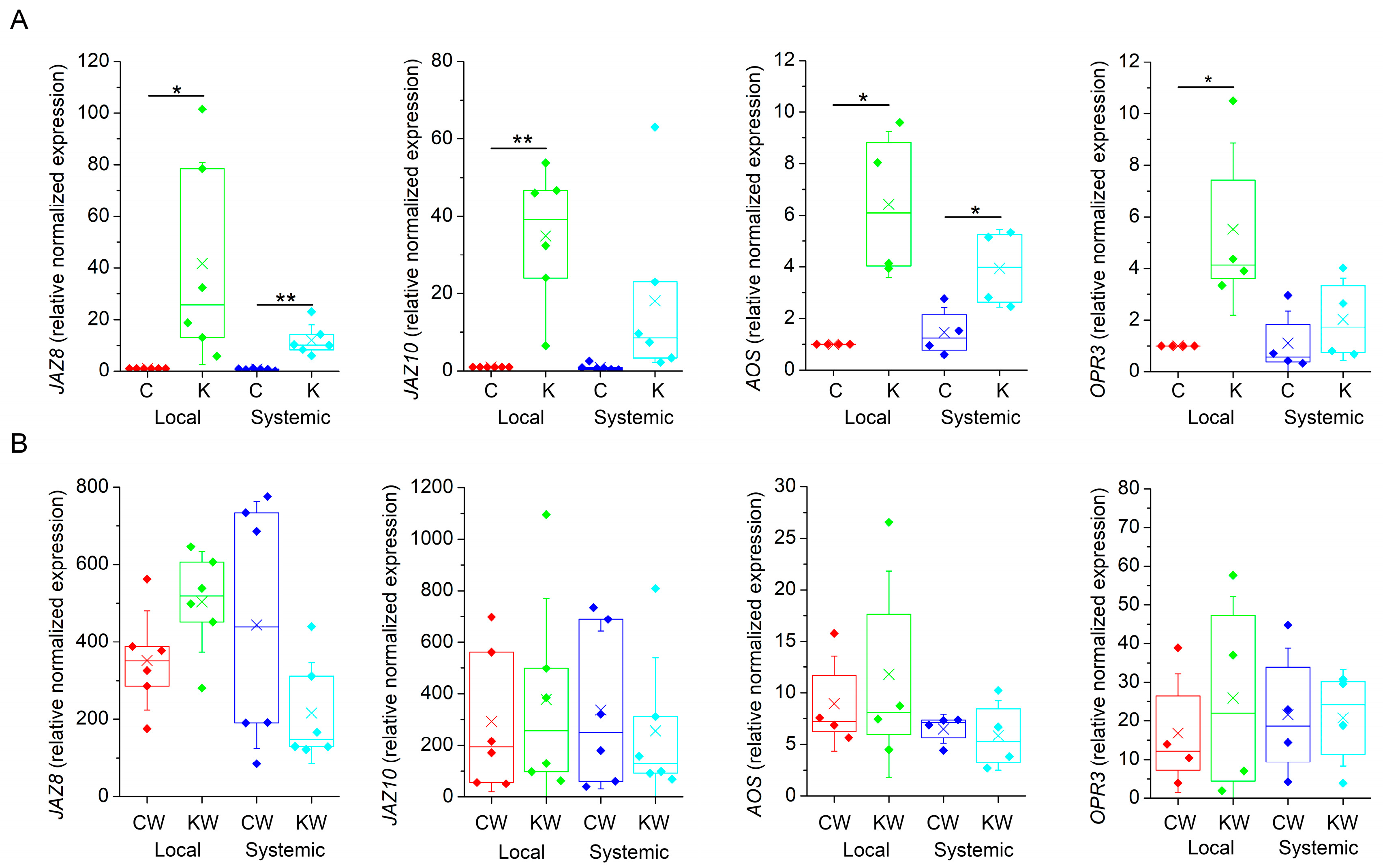
2.3. Ketamine Did Not Block Expression of Jasmonate Responsive Genes
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Setup
4.2. Extracellular Measurements of Electrical Signals
4.3. Aequorin Luminescence Imaging
4.4. qPCR
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Correction Statement
References
- Koo, A.J.K.; Howe, G.A. The wound hormone jasmonate. Phytochemistry 2009, 70, 1571–1580. [Google Scholar] [CrossRef]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef] [PubMed]
- Schilmiller, A.L.; Howe, G.A. Systemic signaling in the wound response. Curr. Opin. Plant Biol. 2005, 8, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, S.A.R.; Chauvin, A.; Pascaud, F.; Kellenberger, S.; Farmer, E.E. GLUTAMATE RECEPTOR-LIKE genes mediate leaf-to-leaf wound signalling. Nature 2013, 500, 422–426. [Google Scholar] [CrossRef]
- Gilroy, S.; Suzuki, N.; Miller, G.; Choi, W.-G.; Toyota, M.; Devireddy, A.R.; Mittler, R. A tidal wave of signals: Calcium and ROS at the forefront of rapid systemic signaling. Trends Plant Sci. 2014, 19, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Gilroy, S.; Białasek, M.; Suzuki, N.; Górecka, M.; Devireddy, A.R.; Karpinski, S.; Mittler, R. ROS, calcium, and electric signals: Key mediators of rapid systemic signaling in plants. Plant Physiol. 2016, 171, 1606–1615. [Google Scholar] [CrossRef] [PubMed]
- Toyota, M.; Spencer, D.; Sawai-Toyota, S.; Jiaqi, W.; Zhang, T. Glutamate triggers long-distance, calcium-based plant defense signaling. Science 2018, 361, 1112–1115. [Google Scholar] [CrossRef]
- Suda, H.; Toyota, M. Integration of long-range signals in plants: A model for wound-induced Ca2+, electrical, ROS, and glutamate waves. Curr. Opin. Plant Biol. 2022, 69, 102270. [Google Scholar] [CrossRef]
- Wildon, D.C.; Thain, J.F.; Minchin, P.E.H.; Gubb, I.R.; Reilly, A.J.; Skipper, Y.D.; Doherty, H.M.; O’Donnell, P.J.; Bowles, D.J. Electrical signalling and systemic proteinase inhibitor induction in the wounded plant. Nature 1992, 360, 62–65. [Google Scholar] [CrossRef]
- Lam, H.M.; Chiu, J.; Hsieh, M.H.; Meisel, L.; Oliveira, I.C.; Shin, M.; Coruzzi, G. Glutamate-receptor genes in plants. Nature 1998, 396, 125–126. [Google Scholar] [CrossRef]
- Price, M.B.; Jelesko, J.; Okumoto, S. Glutamate receptor homologs in plants: Functions and evolutionary origins. Front. Plant Sci. 2012, 3, 235. [Google Scholar] [CrossRef] [PubMed]
- Wudick, M.M.; Michard, E.; Nunes, C.O.; Feijó, J.A. Comparing plant and animal glutamate receptors: Common traits but different fates? J. Exp. Bot. 2018, 69, 4151–4163. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.T.; Kurenda, A.; Stolz, S.; Chételat, A.; Farmer, E.E. Identification of cell populations necessary for leaf-to leaf electrical signaling in a wounded plant. Proc. Natl. Acad. Sci. USA 2018, 115, 10178–10183. [Google Scholar] [CrossRef] [PubMed]
- Alfieri, A.; Doccula, F.G.; Pederzoli, R.; Grenzi, M.; Bonza, M.C.; Luoni, L.; Candeo, A.; Armada, N.R.; Barbiroli, A.; Valentini, G.; et al. The structural bases for agonist diversity in an Arabidopsis thaliana glutamate receptor-like channel. Proc. Natl. Acad. Sci. USA 2020, 117, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Shao, Q.; Gao, Q.; Lhamo, D.; Zhang, H.; Luan, S. Two glutamate- and pH-regulated Ca2+ channels are required for systemic wound signaling in Arabidopsis. Sci. Signal. 2020, 13, eaba1453. [Google Scholar] [CrossRef] [PubMed]
- Grenzi, M.; Bonza, M.C.; Alfieri, A.; Costa, A. Structural insights into long-distance signal transduction pathways mediated by plant glutamate receptor-like channels. New Phytol. 2021, 229, 1261–1267. [Google Scholar] [CrossRef] [PubMed]
- Grenzi, M.; Bonza, M.C.; Costa, A. Signaling by plant glutamate receptor-like channels: What else! Curr. Opin. Plant Biol. 2022, 68, 102253. [Google Scholar] [CrossRef]
- Moe-Lange, J.; Gappel, N.M.; Machado, M.; Wudick, M.M.; Sies, C.S.; Schott-Verdugo, S.N.; Bonus, M.; Mishra, S.; Hartwig, T.; Bezrutczyk, M.; et al. Interdependence of a mechanosensitive anion channel and glutamate receptors in distal wound signaling. Sci. Adv. 2021, 7, eabg4298. [Google Scholar] [CrossRef]
- Bellandi, A.; Papp, D.; Breakspear, A.; Joyce, J.; Johnston, M.G.; de Keijzer, J.; Raven, E.C.; Ohtsu, M.; Vincent, T.R.; Miller, A.J.; et al. Diffusion and bulk flow of amino acids mediate calcium waves in plants. Sci. Adv. 2022, 8, eabo6693. [Google Scholar] [CrossRef]
- Grenzi, M.; Buratti, S.; Parmagnani, A.S.; Aziz, I.A.; Bernacka-Wojcik, I.; Resentini, F.; Šimura, J.; Doccula, F.G.; Alfieri, A.; Luoni, L.; et al. Long-distance turgor pressure changes induce local activation of plant glutamate receptor-like channels. Curr. Biol. 2023, 33, 1019–1035. [Google Scholar] [CrossRef]
- Jakšová, J.; Rác, M.; Bokor, B.; Petřík, I.; Novák, O.; Reichelt, M.; Mithöfer, A.; Pavlovič, A. Anaesthetic diethyl ether impairs long-distance electrical and jasmonate signaling in Arabidopsis thaliana. Plant Physiol. Biochem. 2021, 169, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Yokawa, K.; Kagenishi, T.; Pavlovič, A.; Gall, S.; Weiland, M.; Mancuso, S.; Baluška, F. Anaesthetics stop diverse plant organ movements, affect endocytic vesicle recycling and ROS homeostasis, and block action potentials in Venus flytraps. Ann. Bot. 2018, 122, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Pavlovič, A.; Libiaková, M.; Bokor, B.; Jakšová, J.; Petřík, I.; Novák, O.; Baluška, F. Anaesthesia with diethyl ether impairs jasmonate signalling in the carnivorous plant Venus flytrap (Dionaea muscipula). Ann. Bot. 2020, 125, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Scherzer, S.; Huang, S.; Iosip, A.; Kreuzer, I.; Yokawa, K.; Al-Rasheid, K.A.S.; Heckmann, M.; Hedrich, R. Ether anesthetics prevents touch-induced trigger hair calcium-electrical signals excite the Venus flytrap. Sci. Rep. 2022, 12, 2851. [Google Scholar] [CrossRef] [PubMed]
- MacIver, B.M.; Tanelian, D.L. Volatile anesthetics excite mammalian nociceptor afferents recorded in vitro. Anesthesiology 1990, 72, 1022–1030. [Google Scholar] [CrossRef] [PubMed]
- Meyer, H. Zur theorie der Alkoholnarkose. Arch. Exp. Pathol. Phar. 1899, 42, 109–118. [Google Scholar] [CrossRef]
- Overton, C.E. Studien über die Narkose Zugleich ein Beitrag zur Allgemeinen Pharmakologie; Fischer Verlag: Berlin, Germany, 1901. [Google Scholar]
- Pavel, M.A.; Petersen, N.; Wang, H.; Lerner, R.A.; Hansen, S.B. Studies on the mechanism of general anesthesia. Proc. Natl. Acad. Sci. USA 2020, 117, 13757–13766. [Google Scholar] [CrossRef] [PubMed]
- Franks, N.P.; Lieb, W.R. Do general anesthetics act by competitive binding to specific receptors? Nature 1984, 310, 599–601. [Google Scholar] [CrossRef]
- Franks, N.P. Molecular targets underlying general anesthesia. Br. J. Pharmacol. 2006, 147, S72–S81. [Google Scholar] [CrossRef]
- Crowder, C.M. Does natural selection explain the universal response of metazoans to volatile anesthetics? Anesth. Analg. 2008, 107, 862–863. [Google Scholar] [CrossRef]
- Eckenhoff, R.G. Why can all of biology be anesthetized? Anesth. Analg. 2008, 107, 859–861. [Google Scholar] [CrossRef] [PubMed]
- Franks, N.P. General anaesthesia: From molecular targets to neuronal pathways of sleep and arousal. Nat. Rev. Neurosci. 2008, 9, 370–386. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ye, F.; Zhang, T.; Lv, S.; Zhou, L.; Du, D.; Lin, H.; Guo, F.; Luo, C.; Zhu, S. Structural basis of ketamine action on human NMDA receptors. Nature 2021, 596, 301–305. [Google Scholar] [CrossRef]
- Yu, B.; Wu, Q.; Li, X.; Zeng, R.; Min, Q.; Huang, J. GLUTAMATE RECEPTOR-like gene OsGLR3.4 is required for plant growth and systemic wound signaling in rice (Oryza sativa). New Phytol. 2022, 233, 1238–1256. [Google Scholar] [CrossRef]
- Pavlovič, A.; Jakšová, J.; Kučerová, Z.; Špundová, M.; Rác, M.; Roudnický, P.; Mithöfer, A. Diethyl ether anesthesia induces transient cytosolic [Ca2+] increase, heat shock proteins, and heat stress tolerance of photosystem II in Arabidopsis. Front. Plant Sci. 2022, 13, 995001. [Google Scholar] [CrossRef] [PubMed]
- Kelz, M.B.; Mashour, G.A. The biology of general anesthesia from paramecium to primate. Curr. Biol. 2019, 29, R1199–R1210. [Google Scholar] [CrossRef]
- Kumari, A.; Chételat, A.; Nguyen, C.T.; Farmer, E.E. Arabidopsis H+-ATPase AHA1 controls slow wave potential duration and wound-response jasmonate pathway activation. Proc. Natl. Acad. Sci. USA 2019, 116, 20226–20231. [Google Scholar] [CrossRef]
- Marwaha, J. Some mechanisms underlying actions of ketamine on electromechanical coupling in skeletal muscle. J. Neurosci. Res. 1980, 5, 43–50. [Google Scholar] [CrossRef]
- MacDonalds, J.F.; Bartlett, M.C.; Mody, I.; Pahapill, P.; Reynolds, J.N.; Salter, M.W.; Schneiderman, J.H.; Pennefather, P.S. Actions of ketamine, phencyclidine and MK-801 on NMDA receptor currents in cultured mouse hippocampal neurones. J. Physiol. 1991, 432, 483–508. [Google Scholar] [CrossRef]
- Hatakeyama, N.; Yamazaki, M.; Shibuya, N.; Yamamura, S.; Momose, Y. Effects of ketamine on voltage-dependent calcium currents and membrane potentials in single bullfrog atrial cells. J. Anesth. 2001, 15, 149–153. [Google Scholar] [CrossRef]
- Yin, J.; Fu, B.; Wang, Y.; Yu, T. Effects of ketamine on voltage-gated sodium channels in the barrel cortex and the ventral posteromedial nucleus slices of rats. Neuroreport 2019, 30, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Zorumski, C.F.; Izumi, Y.; Mennerick, S. Ketamine: NMDA receptors and beyond. J. Neurosci. 2016, 36, 11158–11164. [Google Scholar] [CrossRef] [PubMed]
- De Luccia, T.P. Mimosa pudica, Dionaea muscipula and anesthetics. Plant Signal. Behav. 2012, 7, 1163–1167. [Google Scholar] [CrossRef] [PubMed]
- Böhm, J.; Scherzer, S. Signaling and transport processes related to the carnivorous lifestyle of plants living on nutrient-poor soil. Plant Physiol. 2021, 187, 2017–2031. [Google Scholar] [CrossRef] [PubMed]
- Iosip, A.L.; Böhm, J.; Scherzer, S.; Al-Rasheid, K.A.S.; Dreyer, I.; Schultz, J.; Becker, D.; Kreuzer, I.; Hedrich, R. The Venus flytrap trigger hair-specific potassium channel KDM1 can reestablish the K+ gradient required for hapto-electric signaling. PLoS Biol. 2020, 18, e3000964. [Google Scholar] [CrossRef]
- Hermanns, H.; Hollmann, M.W.; Stevens, M.F.; Lirk, P.; Brandenburger, T.; Piegeler, T.; Werdehausen, R. Molecular mechanisms of action of systemic lidocaine in acute and chronic pain: A narrative review. Brit. J. Anaesth. 2019, 123, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Hedrich, R. Ion channels in plants. Physiol Rev. 2012, 92, 1777–1811. [Google Scholar] [CrossRef] [PubMed]
- Milne, A.; Beamish, T. Inhalational and local anesthetics reduce tactile and thermal responses in Mimosa pudica. Can. J. Anaesth. 1999, 46, 287–289. [Google Scholar] [CrossRef]
- Hagihara, T.; Mano, H.; Miura, T.; Hasebe, M.; Toyota, M. Calcium-mediated rapid movements defend against herbivorous insects in Mimosa pudica. Nat. Commun. 2022, 13, 6412. [Google Scholar] [CrossRef]
- Pavlovič, A.; Kopečná, M.; Hloušková, L.; Koller, J.; Hřivňacký, M.; Ilík, P.; Bartoš, J. Diethyl ether anaesthesia inhibits de-etiolation of barley seedlings by locking them in intermediate skoto-photomorphogenetic state. Physiol. Plantarum 2024, 176, e14144. [Google Scholar] [CrossRef]
- Ilík, P.; Hlaváčková, V.; Krchňák, P.; Nauš, J. A low-noise multi-channel device for the monitoring of systemic electrical signal propagation in plants. Biol. Plantarum 2010, 54, 185–190. [Google Scholar] [CrossRef]
- Kiep, V.; Vadassery, J.; Lattke, J.; Maaβ, J.-P.; Boland, W.; Peiter, E.; Mithöfer, A. Systemic cytosolic Ca2+ elevation is activated upon wounding and herbivory in Arabidopsis. New Phytol. 2015, 207, 996–1004. [Google Scholar] [CrossRef]
- Chini, A.; Monte, I.; Zamarreño, A.M.; Hamberg, M.; Lassuer, S.; Reymond, P.; Weiss, S.; Stintzi, A.; Schaller, A.; Porzel, A.; et al. An OPR3-independent pathway uses 4,5-didehydrojasmonate for jasmonate synthesis. Nat. Chem. Biol. 2018, 14, 171–178. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, 2003–2007. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavlovič, A.; Ševčíková, L.; Hřivňacký, M.; Rác, M. Effect of the General Anaesthetic Ketamine on Electrical and Ca2+ Signal Propagation in Arabidopsis thaliana. Plants 2024, 13, 894. https://doi.org/10.3390/plants13060894
Pavlovič A, Ševčíková L, Hřivňacký M, Rác M. Effect of the General Anaesthetic Ketamine on Electrical and Ca2+ Signal Propagation in Arabidopsis thaliana. Plants. 2024; 13(6):894. https://doi.org/10.3390/plants13060894
Chicago/Turabian StylePavlovič, Andrej, Lucie Ševčíková, Martin Hřivňacký, and Marek Rác. 2024. "Effect of the General Anaesthetic Ketamine on Electrical and Ca2+ Signal Propagation in Arabidopsis thaliana" Plants 13, no. 6: 894. https://doi.org/10.3390/plants13060894
APA StylePavlovič, A., Ševčíková, L., Hřivňacký, M., & Rác, M. (2024). Effect of the General Anaesthetic Ketamine on Electrical and Ca2+ Signal Propagation in Arabidopsis thaliana. Plants, 13(6), 894. https://doi.org/10.3390/plants13060894